
基于环境因子的东南太平洋茎柔鱼资源补充量预报模型研究*
2014-03-19 12:17汪金涛陈新军
海洋与湖沼 2014年6期
汪金涛 陈新军① 高 峰 雷 林
(1. 上海海洋大学海洋科学学院 上海 201306; 2. 大洋渔业资源可持续开发省部共建教育部重点实验室, 上海 201306;3. 国家远洋渔业工程技术研究中心 上海 201306; 4. 远洋渔业协同创新中心 上海 201306)
茎柔鱼(Dosidicus gigas)是一种大洋性头足类,广泛分布于东太平洋海域(王尧耕等, 2005)。目前世界上多个国家和地区已对茎柔鱼资源进行了开发和利用, 年产量超过80万吨(Nigmatullinet al, 2001;Waludaet al, 2004)。资源量预报属于中长期渔情预报的一种, 对资源补充量进行精确的预报是确保渔业资源科学管理、合理开发的关键(陈新军, 2004)。目前, 已有一些关于茎柔鱼资源补充量的研究。如Anderson等(2001)和Waluda等(2006)研究认为, 厄尔尼诺和拉尼娜事件会影响茎柔鱼幼体和成熟体的生长、栖息等生活史过程。陈新军等(2006)指出, 茎柔鱼资源状况与海洋环境关系密切, 特别是厄尔尼诺和拉尼娜事件。徐冰(2012)等利用典型相关性分析,分析了表温(SST)、表温距平均值(SSTA)、茎柔鱼产卵场、索饵场最适表层水温范围占总面积的比例(分别用Ps、PF表示)与资源丰度、补充量相关性, 并建立了基于SST因子的资源丰度线性预报模型。也有研究(Ichiiet al, 2002)指出, 水温并非影响茎柔鱼资源丰度和补充量的主要限制因子。以上前人的研究基本上是围绕水温因子展开的。因此, 本文在前人研究的基础上增加更多的海洋环境因子, 尝试找出更为合适的影响茎柔鱼资源补充量的海洋环境指标, 并利用误差反向传播神经网络(Error Backpropagation Network, EBP)建立更为准确的资源补充量预报模型,为东南太平洋海域茎柔鱼的科学管理和生产提供参考依据。
1 材料与方法
1.1 材料来源
东南太平洋茎柔鱼的生产数据来自上海海洋大学鱿钓技术组, 时间为2003年1月—2012年12月。内容包括日期、经度、纬度、日产量、渔船数。
SST、SSH、chla浓度数据均来自http: //oceanwatch.pifsc.noaa.gov/las/servlets/dataset, 时间分辨率为月; SST空间分辨率为0.1°×0.1°, SSH、chla浓度空间分辨率为0.25°×0.25°; 经纬度范围是20°S—20°N、110°W—70°W。
本文环境数据空间分辨率统一为0.5°×0.5°, 不同空间尺度的环境数据都是由原始空间尺度转换而成, 如每一个空间尺度为0.5°×0.5°的SST数据是计算25个原始数据的平均值而得到, 空间分辨率转换工作由作者自主开发软件FisheryDataProcess完成。
1.2 研究方法
1.2.1 研究海域范围 前人研究表明, 茎柔鱼广泛分布于东太平洋的加利福利亚(37°N)到智利(47°S)的海域中, 在赤道附近海域可达到140°W(Nigmatullinet al, 2001; Waludaet al, 2004)。根据我国鱿钓船的实际作业情况, 选定本研究的海域范围为20°S—20°N、110°W—70°W。
1.2.2 CPUE计算 CPUE(Catch per unit effort)定义如下:

其中CatchY、VesY分别表示Y年的捕捞产量和作业渔船数。计算每年的单船平均日产量CPUE(t/d)作为茎柔鱼资源丰度指标。由于茎柔鱼是短生命周期, 产卵后即死, 没有剩余群体的生物学特性, 因此CPUE也可作为反映其资源补充量的指标。
1.2.3 影响因子选取 茎柔鱼资源补充量与其产卵场和索饵场的栖息环境密切相关(Nigmatullinet al,2001; Waludaet al, 2004; 陈新军等, 2006; 徐冰等,2012)。因此, 根据公式(2)分别计算分析1—12月份研究海域每点(0.5°×0.5°) SST、SSH、chla浓度组成的时间序列值与本年和次年CPUE组成的时间序列值的相关性, 选取相关性高海域的SST、SSH、chla浓度作为茎柔鱼资源补充量的影响因子。其中, SST、SSH、chla浓度与本年CPUE相关性高的海域表示索饵栖息环境对资源补充量的影响; SST、SSH、chla浓度与次年CPUE相关性高的海域表示产卵栖息环境对资源补充量的影响。

其中x、y分别表示环境、CPUE组成的系列值,为x、y的平均值,r表示相关系数。
产卵场、索饵场最适表层水温范围占总面积的比例是衡量栖息地环境优劣的参量(Nigmatullinet al,2001; Waludaet al, 2004)。有文献表明(Nigmatullinet al, 2001; Taipeet al, 2001; Ichiiet al, 2002), 9月份茎柔鱼产卵时适宜SST为24—28°C; 7月份茎柔鱼的索饵时适宜SST为17—22°C。因此分别计算9月份产卵时、7月份索饵时最适表层水温范围占总面积的比例(分别用PS、PF表示), 用PS、PF表达产卵场索饵场栖息环境的适宜程度。
1.2.4 建立预报模型 EBP属于多层前向神经网络, 采用误差反向传播的监督算法, 能够学习和存储大量的模式映射关系, 已被广泛应用于各个领域(Benediktssonet al, 1990; Hushet al, 1993)。
EBP神经网络有输入层、隐含层和输出层, 每层由不同个数的神经元或节点组成(图1), 其学习的过程分为信号的正向传播与误差的反向传播两个阶段。在正向传播阶段, 输入信息从输入层经隐含层传向输出层, 在输出层产生输出信号。在信号的向前传递过程中网络的权值固定不变, 每一层神经元的状态只影响下一层神经元的状态。如果在输出层不能得到期望的输出, 则转入误差反向传播。在反向传播阶段,未能满足精度要求的误差信号由输出端开始, 以某种方式逐层向前传播, 并将误差分摊给各层的所有单元, 依据误差信号动态的调整各单元的连接权值。通过周而复始的正向传播和反向调节, 神经元间的权值得到不断的修正。当误差满足设定的精度后, 将停止学习。
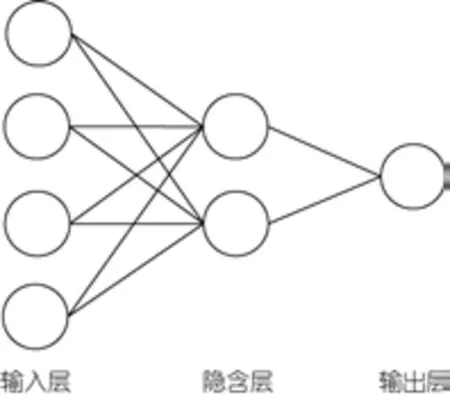
图1 EBP神经网络结构Fig.1 The structure of EBP(error backpropagation network)
EBP神经网络的建立在Matlab软件中完成, 首先对样本进行归一化处理, 使样本处在0—1间。使用神经网络工具箱的拟合工具, 将2003—2011年的样本作为训练样本, 2012年的样本作为验证样本。网络设计的参数为: 输入层神经元个数根据1.2.3选定的显著相关因子、PF以及PS的组合而决定, 输出层神经元1个为CPUE, 隐含层神经元个数根据经验公式(公式(3))得到。学习速率为0.1, 动量参数为0.5。网络训练的终止参数为: 最大训练批次100次, 最大误差给定0.001。模型训练10次, 取最优结果, 同时防止过拟合状态的出现。

其中l、n、m分别表示输入层、隐藏层、输出层神经元个数, d为0到10之间的常数。
EBP模型以均方误差(MSE)作为判断最优模型的标准。拟合残差是将预报值与实际值进行比较所得,其函数定义式为yk为CPUE的实际值,yˆk为CPUE的预报值,N表示CPUE的个数,k表示CPUE序数。
2 结果
2.1 年CPUE变化
图2显示, 2003—2012年茎柔鱼CPUE年间波动较大, CPUE较低年份出现在2005年、2008年、2009年和2012年, 最低年份CPUE出现在2007年为4.03t/d, CPUE最高年份出现在2004年为7.07t/d。
2.2 特征环境因子分析
在1—12月份20°S—20°N、110°W—70°W海域范围内, 各月每一个0.5°×0.5°的SST与本年和次年CPUE作相关性分析, 发现SST与本年CPUE相关性最大值出现在7月份的13°N, 102°W(Point1)(表1, 图3,图4a), SST与次年CPUE相关性最大值出现在6月份的8°N, 103.5°W(Point2)(表1, 图3, 图4c)处。
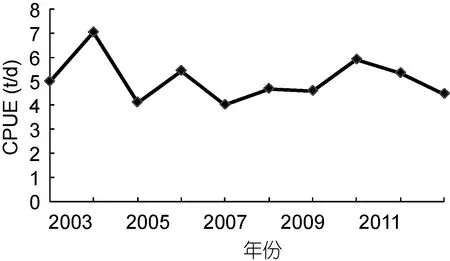
图2 2003—2012年东南太平洋茎柔鱼CPUE变化Fig.2 Variation in CPUE of Dosidicus gigas during 2003 to 2012

图3 与茎柔鱼资源补充量显著相关的关键海区Fig.3 The key points significantly associated with CPUE and its environmental factors
在1—12月份20°S—20°N、110°W—70°W海域范围内, 各月每一个0.5°×0.5°的SSH与本年和次年CPUE作相关性分析, 发现SSH与本年CPUE相关性最大值出现在9月份的11°N, 102°W(Point3)(表1, 图3, 图4b)处, SSH与次年CPUE相关性最大值出现在2月的12°N, 97.5°W(Point4)(表1, 图3, 图4d)处。
在1—12月份20°S—20°N, 110°W—70°W海域范围内, 各月每一个0.5°×0.5°的chla浓度与本年和次年CPUE作相关性分析, 发现chla浓度与本年CPUE相关性最大值出现在3月份的8°S, 107°W(Point5)(表1,图3, 图4e)处, chla浓度与次年CPUE相关性最大值出现在10月的10°S, 93.5°W(Point4)(表1, 图3, 图4f)处。
2.3 预报模型实现及结果比较
利用选定的关键海区环境因子以及PS、PF的不同组合作为EBP预报模型的输入因子, 构造多种EBP预报模型, 分别为:

图4 特征环境因子与茎柔鱼资源补充量的线性关系式Fig.4 The linear regression of CPUE and environmental factors at the key points
方案1: 选取Point1的SST、Point3的SSH、Point5的chla浓度、PF共4个因子作为输入层, 构造4: 5: 1的EBP网格结构, 表示利用索饵环境关键影响因子建立的预报模型。
方案2: 选取Point2的SST、Point4的SSH、Point6的chla浓度、PS共4个因子作为输入因子, 构造4: 5:1的网络结构, 表示利用产卵环境关键影响因子建立的预报模型。
方案3: 选择Point1与Point2的SST、Point3与Point4的SSH、Point5与Point6的chla浓度、PS、PF共8个因子作为输入因子, 构造8: 9: 1的网络结构,表示利用综合环境关键因子建立的预报模型。
利用Matlab进行建模, 计算三种方案下的均方误差(图5), 方案2和方案3的均方误差相近且优于方案1, 其准确率在90%左右。

图5 不同神经网络模型的模拟结果和准确率Fig.5 The simulation result of different neural network models
3 讨论
茎柔鱼属于短生命周期种类。通常认为, 东太平洋海域受两个低速东部边界流(秘鲁海流和加利福尼亚海流)影响, 并在信风作用下产生上升流。上升流区域环境对茎柔鱼幼体和成熟体的生长、栖息等生活史过程具有十分重要的影响。两海流的相对强弱影响上升流区域的栖息环境, 进而影响茎柔鱼的产卵、索饵等过程, 最终影响年度资源丰度的变化。如2003年Point2的SST为29.2°C, 温度较高, 形成产卵场PS比例大, 产卵环境好, 同时2004年Point1的SST为28.98°C温度偏低, 说明秘鲁海流强劲, 形成的上升流中海水的营养盐更为丰富, 索饵环境较好, 形成2004年CPUE较高的原因。相反地, 2006年Point2的SST为28.5°C, 温度偏低, 形成产卵场PS比例小,产卵环境较差, 加上2007年Point1的SST为29.6°C温度偏高, 秘鲁海流势力弱, 索饵环境较差, 导致2007年的的CPUE很低。
已有的研究表明, 利用茎柔鱼栖息地环境因子来预报茎柔鱼资源补充量是可行的(Taipeet al, 2001;陈新军等, 2006; 徐冰等, 2012)。本文利用SST、SSH、chla浓度三种环境因子与CPUE进行相关性分析, 选取的相关系数最大值点与随机选取的2010年相同月份的环境因子叠加发现(图6), 关键海区均在环境因子等值线之上或附近, 这些位置环境因子的变化可以作为资源丰度或资源补充量的一个“指示器”, 其中SST位置与徐冰(2012)的研究结果基本相似。
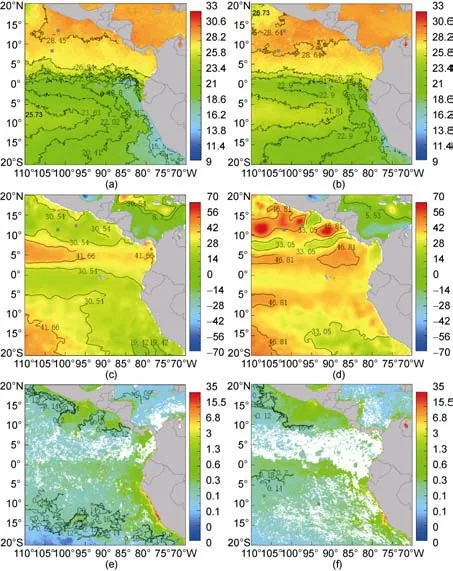
图6 关键区域与其2010年环境因子, (a)(b)为7月份和6月份SST, (c)(d)为9月份和2月份SSH, (e)(f)为3月份和10月份chl a浓度Fig.6 The key points and its environmental data in 2010 a: SST in July; b: SST in June; c: SSH in September; d: SSH in February; e: chl-a concentration in March; f: chl-a concentration in October
比较三种方案下的EBP神经网络模型发现(图5)方案2和方案3结果相近且优于方案1, 表明产卵栖息环境对资源补充量的影响要大于索饵栖息环境对资源补充量的影响, 比较符合短生命周期种类的特征。
茎柔鱼资源补充量预报是一件极其复杂的系统工作。茎柔鱼在其早期生活阶段不仅仅受到海域环境因素的影响, 还受到来自外部和内部的捕食者的影响, 利用其产卵、索饵等生活阶段海域环境因素进行资源量评估只是其中一种重要方法。今后需要结合物理海洋学、生态系统动力学等, 综合其个体的生长、死亡等因素以及海流、初级生产力等因子, 建立更全面、科学的茎柔鱼资源补充量预报模型, 为茎柔鱼资源合理利用和科学管理提供依据。
王尧耕, 陈新军, 2005. 世界大洋性经济柔鱼类资源及其渔业.北京: 海洋出版社, 240—264
陈新军, 2004. 渔业资源与渔场学. 北京: 海洋出版社, 169—192
陈新军, 赵小虎, 2006. 秘鲁外海茎柔鱼产量分布及其与表温关系的初步研究. 上海水产大学学报, 15(1): 65—70
徐 冰, 2012. 秘鲁外海茎柔鱼渔场时空分布及资源补充量与环境的关系. 上海: 上海海洋大学硕士学位论文, 45—50
徐 冰, 陈新军, 李建华, 2012. 海洋水温对茎柔鱼资源补充量影响的初探. 上海海洋大学学报, 21(5): 878—883
徐 冰, 陈新军, 陆化杰等, 2013 . 秘鲁外海茎柔鱼资源丰度和补充量与海表温度的相关关系. 海洋渔业, 35(3): 296—302
Anderson C I H, Rodhouse P G, 2001. Life cycles, oceanography and variability: ommastrephid squid in variable oceanography environments. Fisheries Research, 54(1): 133—143
Benediktsson J, Swain P H, Ersoy O K, 1990. Neural network approaches versus statistical methods in classification of multisource remote sensing data. Transactions on Geoscience and Remote Sensing, 28(4): 540—552
Cairistiona I, Anderson H, Rodhouse P, 2001. Life cycles,oceanography and variability: Ommastrephid squid in variable oceanographic environments. Fisheries Research(Amsterdam), 54(1): 133—143
Hush D R, Horne B G, 1993. Progress in supervised neural networks. Signal Processing Magazine, 10(1): 8—39
Ichii T, Mahapatra K. Watanabe Tet al, 2002. Occurrence of jumbo flying squidDosidicus gigasaggregations associated with the countercurrent ridge off the Costa Rica Dome during 1997 El Niño and 1999 La Niña. Marine Ecology,231: 151—166
Nigmatullin C M, Nesis K N, Arkhipkin A, 2001. A review of the biology of the jumbo squid Dosidicus gigas (Cephalopoda:Ommastrephidae). Fisheries Research, 54(1): 9—19
Taipe A, Yamashiro C, Mariategui Let al, 2001. Distribution and concentrations of jumbo flying squid (Dosidicus gigas) off the Peruvian coast between 1991 and 1999. Fisheries Research (Amsterdam), 54(1): 21—32
Waluda C, Yamashiro C, Elvidge Cet al, 2004. Quantifying light-fishing forDosidicus gigasin the eastern Pacific using satellite remote sensing. Remote Sensing of Environment,91(2): 129—133
Waluda C, Yamashiro C, Rodhouse P, 2006. Influence of the ENSO cycle on the light-fishery forDosidicus gigasin the Peru Current: An analysis of remotely sensed data. Fisheries Research, 79(1—2): 56—63
猜你喜欢
矿山安全信息(2022年22期)2022-11-24
中国宝玉石(2021年5期)2021-11-18
哈尔滨轴承(2020年2期)2020-11-06
石油化工建设(2020年1期)2020-08-24
今日中国·法文版(2020年7期)2020-07-04
中国新闻周刊(2019年46期)2019-12-20
中国特种设备安全(2019年1期)2019-03-13
海洋信息技术与应用(2017年2期)2017-06-21
山东青年(2016年2期)2016-02-28
中华海洋法学评论(2013年2期)2013-03-11
