抗TSLP 全人源单链抗体筛选与初步鉴定①
2014-03-18 11:41朱建光徐文峰年四季泸州医学院基础医学院泸州646000
中国免疫学杂志 2014年12期
朱建光 袁 青 黄 黎 徐文峰 年四季 (泸州医学院基础医学院,泸州646000)
人胸腺基质淋巴细胞生成素(Thymic stromal lymphopoietin,TSLP)是一种新型的、与白细胞介素-7(IL-7)类似的细胞因子,当过敏性炎症反应时在皮肤角质形成细胞和气道上皮细胞高度表达,能强烈刺激髓系树突状细胞(Dendritic cell,DC)介导的Th2 型炎症反应发生[1,2]。1994 年,由Firend 等[3]首次从胸腺基质细胞的培养上清中分离获得,但是在随后几年时间里对TSLP 的研究进展缓慢,2000年后,随着对TSLP 及其受体(TSLPR)的克隆和测序,人们对其功能才有了较全面的了解,特别是在阐明过敏性炎症疾病的发病机制方面有了更新的认识。TSLP 分子主要作用于CD11c+DC,对DC 的活化、分化起着重要的调控作用。该受体复合物由IL-7R 和另一公用ε 链类似分子(εcR)组成的异源二聚体构成[4]。研究显示,CD11c+DC 可同时表达IL-7R 和εcR,而这种现象在其他细胞则较少见。受TSLP 作用而被激活的CD11c+DC 可促进CD4+T 细胞增殖,分泌大量IL-5、IL-13 及TNF-α[5,6]。同时,CD8+T 细胞在TSLP 激活DC 的作用下,可产生大量IFN-γ、IL-5 和IL-13。IL-5、IL-13 和IFN-γ 水平的升高,可导致炎性反应的加剧以及组织损伤,表明TSLP 可能是过敏性炎症发生过程中一个重要的启动因子[7],为治疗过敏性炎症疾病的主要靶分子。
基于TSLP 在过敏性炎症发生过程中起到的重要作用,制备抗TSLP 全人源单链抗体,从而封阻TSLP 的生物学活性,对于治疗过敏性炎症疾病提供了新途径。
1 材料与方法
1.1 材料 Invitrogen 公司合成TSLP cDNA;pET101/ D-TOPO 载体、Ni-NTA 亲和纯化系统、蛋白预染Marker(BenchMarkTMpre-stained protein ladder)、链霉亲和素M-280 免疫磁珠(streptavidincoated Dynabeads M-280)购自Invitrogen 公司;Ex Taq 酶、DNA 标准分子量(1 000 bp DNA ladder)、低分子量蛋白Marker 购自TaKaRa 公司;E.coli L21、E.coli DH5aF'、E.coli TG1、辅助噬菌体M13K07 及pCANTAB-5E 噬菌体载体均购自Gene 公司;EZLink (tm)Sulfo-NHS-LC-Biotin 生物素购自Thermo Scientific 公司。
1.2 方法
1.2.1 TSLP 基因扩增 公司合成TSLP cDNA,根据基因序列,设计引物。用于扩增TSLP 的上游引物TSLPF1-Forword:5' CACCATGTTCCCTTTT-GCCTTACT 3',下游引物TSLPR1-Reverse:5' CTGTTGTTTCAGTAAGGGTCG 3' 。取 10 ng/μl TSLP cDNA 2 μl 作为模板,进行PCR 扩增。扩增程序为94℃2 min,然后94℃30 s,55℃30 s,72℃1 s,30次循环,最后延伸72℃10 min。
1.2.2 TSLP 的表达纯化 取TSLP PCR 产物,按照pET Directional kit (Invitrogen,USA)说 明 书 与pET101/D-TOPO 载体连接,连接产物转化E.coli One Shot-TOPO,PCR 鉴定阳性克隆子,提取阳性克隆子质粒,测序验证正确后,将含正确序列的TSLP/pET101阳性重组质粒转化E.coli BL21。挑单克隆子过夜培养后取5 ml 接种到100 ml LB 含50 μg/ml 羧苄青霉素的液体培养基中,37℃,200 r/min 培养至OD600=0.6 左右,加入终浓度为1 mmol/L IPTG 于37℃,180 r/min 培养5 h 后,8 000 r/min 离心15 min,收集沉淀用于SDS-PAGE 鉴定和后续纯化。
按Invitrogen 公司的Ni-NTA 亲和纯化系统说明书对表达的TSLP 蛋白进行纯化,并进行Western blot鉴定。由于pET101/D-TOPO 载体带有6 × His-tag,鉴定表达、纯化的蛋白采用抗His-tag(GE Healthcare,来源于小鼠)和羊抗小鼠(IgG)辣根过氧化物酶标记抗体(Promega,USA),最后加入化学发光剂(Luminata Crescendo Western HRP substrate,USA)进行显色。
1.2.3 抗TSLP 全人源单链抗体的筛选 本实验前期已构建了天然全人源scFv 抗体文库,文库容量达到2.5×108,多样性好[8],纯化的TSLP 蛋白根据EZ-Link (tm)Sulfo-NHS-LC-Biotin 试剂盒说明书(Thermo Scientific)进行生物素化。约5×1011TU 新鲜配制的scFv 抗体文库噬菌体用封闭液于室温封闭1 h,加入15 μg 生物素化蛋白于封闭后的scFv抗体文库噬菌体中,于室温缓慢振荡孵育1 h,静置1 h;按说明书加入室温封闭1 h 后的链霉亲和素M-280 免疫磁珠于混合液中,室温缓慢振荡30 min 捕获TSLP 蛋白/抗TSLP 特异性噬菌体复合体。用1×PBS 和1×PBST(1×PBS 含0.05% Tween 20)缓冲液各洗涤磁珠5~10 次,然后用0.1 mol/L Glyline-HCl (pH2.2)使特异性噬菌体解离,感染大肠杆菌TG1 后涂2×YTAG (2×YT 固体培养基平板含100 μg/ml 氨苄青霉素和2% 葡萄糖),从平板上洗下所有菌落用于噬菌体文库扩增。分别以7 μg、3 μg 生物素化的TSLP 蛋白用于第2 轮和第3 轮抗体的筛选。
1.2.4 噬菌体扩增 取200 μl 从平板上洗下的抗体文库菌液加入到30 ml 2×YTAG 培养基中,于37℃,200 r/min 培养至OD600=0.2。离心收集细菌沉淀,用30 ml 2×YTA (2×YT 液体培养基含100 μg/ml 氨苄青霉素)重悬细菌沉淀,加入约3×109TU 辅助噬菌体M13K07,37℃静止感染15min,然后37℃,200 r/min 振荡培养2 h。加入终浓度20 μg/ml 卡拉霉素后于32℃,180 r/min 振荡培养过夜。次日用PEG/NaCl 溶液(20% PEG,2.5 mol/L NaCl)沉淀噬菌体,用1×PBS 缓冲液重悬抗体文库噬菌体。
1.2.5 噬菌体 ELISA 包被液(0.1 mol/L NaHCO3/ Na2CO3溶液,pH9.6)将TSLP 纯化蛋白稀释为30 μg/ml,于酶标板内4℃包被过夜。次日封闭液(1×PBS 含5% 脱脂奶粉和0.05% Tween 20)37℃封闭酶标孔1 h,1×PBST(1×PBS 含0.05%Tween 20)缓冲液洗涤酶标孔。加入等体积封闭液室温封闭30 min 的抗体文库噬菌体溶液,37℃孵育1 h。洗涤酶标孔后,加入抗M13 单克隆抗体37℃孵育1 h。洗涤酶标孔后,加入马抗鼠抗AP 标记抗体37℃孵育1 h。最后用PNPP 显色液显色,于405 nm 处读取吸光值(A)。
2 结果
2.1 TSLP 的扩增 公司合成TSLP cDNA 片段。采用设计的引物对TSLP cDNA 进行PCR 扩增,得到扩增片段大小为423 bp,与目的基因大小一致,见图1。经测序和NCBI BLAST 比对,扩增的基因序列正确,为TSLP。
2.2 TSLP 的纯化及鉴定 pET101/D-TOPO 融合表达载体携带有氨苄青霉素抗性基因,但是由于蛋白表达量比较低,所以选择用羧苄青霉素作为抗性筛选对蛋白进行表达。pET101/ D-TOPO 融合表达载体携带有6×His 标签,有利于目的蛋白的鉴定。表达后,对表达产物进行SDS-PAGE 电泳分析,表达的TSLP 蛋白大小为26 kD 左右。经Ni-NTA 亲和纯化系统在变性条件下纯化得到TSLP 蛋白(图2),并进行Dot blot、Western blot 鉴定,证明表达纯化的蛋白即为目的蛋白TSLP(图3)。
2.3 抗TSLP 全人源单链抗体的筛选 用TSLP 生物素化蛋白作为抗原,采用免疫磁珠法对天然全人源scFv 抗体文库进行了3 轮噬菌体展示富集,从3轮富集后的scFv 抗体文库中随机挑取克隆子,进行单克隆子噬菌体扩增表达,对表达的scFv 以抗M13单克隆抗体和马抗鼠抗AP 标记抗体进行ELISA 检测,结果显示:经3 轮富集后,特异性单链抗体得到富集。取富集后48 个克隆子进行ELISA 检测,其中取2 个作为阴性对照(不加入抗原),结果显示,阴性对照OD405值为0.05左右,大部分克隆子与抗原有结合反应,而OD405>0.8 的阳性单链抗体占35%左右(图4)。

图1 PCR 扩增TSLP cDNAFig.1 Amplification of cDNA of TSLP by PCR

图2 SDS-PAGE 鉴定纯化的TSLP 蛋白Fig.2 Purification and identification of TSLP by SDS-PAGE
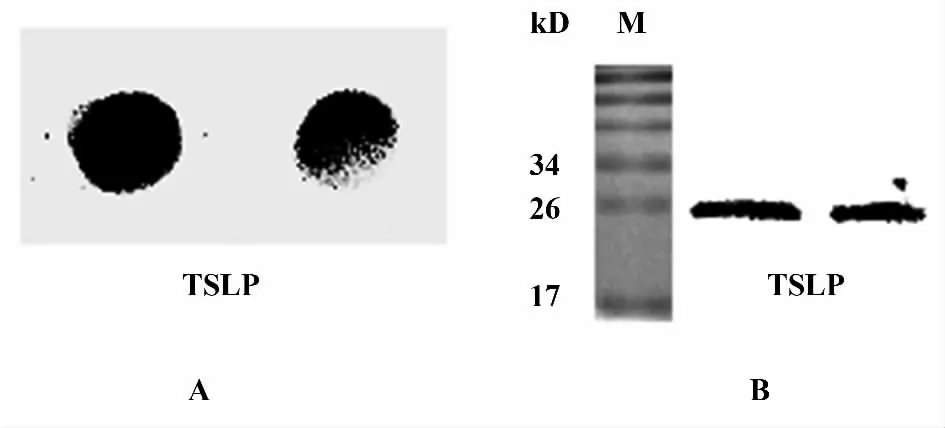
图3 Dot blot 和Western blot 鉴定表达的TSLP 蛋白Fig.3 Identification of expressive TSLP by Dot blot and Western blot

图4 ELISA 筛选抗TSLP 单链抗体Fig.4 Selection of scFv against TSLP by ELISA

图5 Dot blot 和Western blot 鉴定表达的抗TSLP 单链抗体Fig.5 Identification of expressive scFv against TSLP by Dot blot and Western blot
2.4 单链抗体的鉴定 通过ELISA 检测,挑取与TSLP 抗原结合能力强的2 株单链抗体进行表达,对表达产物进行Dot blot、Western blot 鉴定,结果显示表达产物在32 kD 处有单一条带,为表达的单链抗体(图5)。测序结果表明:序列为开放阅读框,为linker (Ser4Gly)3连接的VH 和VL,测序的氨基酸序列见图6。说明从天然全人源scFv 抗体文库中成功的筛选到抗TSLP 全人源单链抗体,但从天然抗体文库中筛选得到单链抗体其亲和力相对较低,后期我们将对单链抗体亲和力进行进化,针对抗TSLP单链抗体构建突变抗体文库,以筛选到高亲和力的全人源抗TSLP 单链抗体。

图6 测序鉴定的TSLP 单链抗体氨基酸序列Fig.6 Amino acid sequence of selected scFv against
3 讨论
TSLP 优先表达于肺、肠、皮肤等部位的上皮细胞,当过敏性炎症发生时,上皮细胞受抗原刺激上调表达TSLP,并在局部激活DC 细胞分化,释放趋化因子,活化成熟的DC 细胞迁移至淋巴结,促使CD4+T 细胞分化并产生Th2 型细胞因子,这些细胞因子进一步放大免疫反应[9,10]。这些研究强调了TSLP 在Th2 介导的炎症反应中的重要性,因此TSLP 靶标的治疗为过敏性炎症疾病提供新手段[11]。体外实验发现,TSLP 转基因鼠易受特异性抗原诱导而发生哮喘,而TSLP 受体敲除的小鼠症状明显减轻,说明TSLP 对于气道过敏反应的发生是必需的[12]。因此针对TSLP 制备中和性抗体成为研究热点,李艳红等[13]体外实验中使用中和性TSLP 抗体对致敏小鼠进行干预,发现Th2 型细胞因子(IL-4、IL-5 和IL-13)明显减少,这说明阻断TSLP可以抑制DC 活化和Th2 型炎症的产生。
本研究中制备的单链抗体为全人源天然抗体文库筛选得到,全人源单链抗体相对于鼠源型、人源化抗体特异性更强,排斥性更小。目前国内外均未见抗TSLP 全人源抗体的问世。本实验后期将对筛选得到的TSLP 单链抗体进行亲和力进化,构建抗TSLP 突变文库,筛选得到亲和力和特异性更高的抗TSLP 单链抗体,为过敏性炎症反应寻找效果更好的治疗途径。
[1]Soumelis V,Reche PA,Kanzler HW,et al.Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP[J].Nat Immunol,2002,3:673-680.
[2]Liu YJ.Thymic stromal lymphopoietin:master switch for allergic inflammation[J].Exp Med,2006,203:269-273.
[3]Firend SL,HosierS,Nelson A,et a1.A thymic stromal cell line supports in vitro development of surface IgM+B cells and produces a novel growth afecting B nad T lineage cells[J].Exp Hematol,1994,22(1):321-328.
[4]Pandey A,Ozaki K,Baumann H,et al.Cloning of a receptor subunit required for signaling by thymic stromal lymphopoietin[J].Nat Immunol,2001,1(1):59-64.
[5]Soumelis V,Reche PA,Kanzler H,et al.Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP[J].Nat Immunol,2002,3(7):673-680.
[6]Watanabe N,Hanabuchi S,Soumelis V,et al.Human thymic stromal lymphopoietin promotes dendritic cell-mediated CD4+Tcell homeostatic expansion[J].Nat Immunol,2004,5(4):426-434.
[7]Gilliet M,Soumelis V,Watanabe N,et al.Human dendritic cell sactivated by TSLP and CD40L induce proallergic cytotoxic T cells[J].Exp Med,2003,97(8):1059-1063.
[8]Qing Y,Huang L,Wang X,et al.Construction of human nonimmune library and selection of scFV against IL-33[J].Appl Biochem Bio-technol,2012,167:498-509.
[9]Quentmeier H,Drexler HG,Fleckenstein D,et al.Cloning of human thymic stremal lymPhoPoietin(TSLP)and signaling mechanisms leading to Proliferation [J].Leukemia,2001,15:1286-1292.
[10]Reehe PA,Soumelis V,Gorman DM,et al.Human thymic stromal lymPhoPoietin preferentially stimulates myeloid cells [J].Immunol,2001,167(1):336-343.
[11]Huston DP,Liu YJ.Thymic stromal lymphopoietin:a potential therapeutic target for allergy and asthma[J].Curr Allergy Asthma Rep,2006,6:372-376.
[12]Zhou BH,Comeau MR,Smelt TD,et al.Thym ic stromal lymphopoietin as a key initiator of allergic airway inflammation in m ice[J].Nat Immunol,2005,6 (10):1047-1053.
[13]李艳红,李鸿佳,亓慧娟,等.哮喘小鼠气道上皮TSLP 表达及激活DCs 加重气道炎症的研究[J].中华微生物学和免疫学杂志,2010,30(4):303-308.
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
植物保护(2021年4期)2021-11-12
中老年保健(2021年2期)2021-08-22
中老年保健(2021年3期)2021-08-22
皮肤病与性病(2021年3期)2021-07-30
力学学报(2021年2期)2021-03-10
中国民间疗法(2020年22期)2021-01-07
供应链管理(2020年10期)2020-12-23
科学24小时(2020年4期)2020-05-14
中国海洋大学学报(自然科学版)(2019年2期)2019-12-07