
刺五加细胞色素P450基因的克隆与表达分析
2014-03-17 11:46邢朝斌,吴鹏,李非非等
生物技术通报 2014年1期
刺五加细胞色素P450基因的克隆与表达分析
邢朝斌 吴鹏 李非非 刘岩 周秘 修乐山
(河北联合大学生命科学学院,唐山 063000)
根据已报道的人参、三七等植物的细胞色素P450(Cytochrome P450,P450)基因的cDNA序列设计引物,利用RT-PCR法克隆刺五加P450基因的cDNA全长序列,并分析其在不同生长发育时期和器官中的表达情况。结果显示,克隆了全长为1 410 bp的刺五加P450基因的cDNA序列,该基因编码469个氨基酸残基组成的蛋白质。GenBank登录号为KF498590,与人参、三七的P450氨基酸序列一致性分别为91.5%和90.4%。刺五加的P450基因在不同生长发育时期和器官中均有表达,但表达量具有显著差异(P<0.05)。最大表达量出现在盛花期,为最低表达量(萌芽期)的1.26倍。各器官中,叶片的表达量最高,是最低量幼茎的1.49倍。
刺五加 细胞色素P450 克隆 表达分析
刺五加Eleutherococcus senticosus(Rupr.et Maxim)Maxim是我国传统的珍贵药用植物,具有抗肿瘤、抗辐射、提高机体免疫力等多种药理作用和生理活性,皂苷类化合物为其主要活性成分之一[1]。与其他药用植物相似,刺五加中的皂苷含量普遍较低。探明三萜类化合物的生物合成途径是利用代谢工程直接合成或通过基因工程工业化生产三萜类化合物的关键基础[2]。三萜类化合物的生物合成一般分为,前体的合成、形成三萜皂苷骨架和后修饰过程3个阶段[3]。目前对三萜类合成的分子机理的研究主要集中在前体合成和形成三萜皂苷骨架阶段,而对决定三萜皂苷种类起主要作用的三萜碳环的复杂修饰过程研究较少[4]。细胞色素P450(cytochrome P450,P450)是三萜皂苷生物合成后修饰过程中的一个关键酶,该酶含有血红素配基,是植物中最大的一个超基因家族,其不同家族成员参与了多种植
物的次生代谢途径[5]。目前仅见少数几个参与三萜皂苷生物合成的P450基因的报道[4]。因此克隆刺五加P450基因的全长序列,并分析其在不同生长发育时期、不同器官中的表达规律,对深入研究P450基因在刺五加皂苷生物合成中的作用机制和表达调控具有重要的意义。
1 材料与方法
1.1 材料
1.1.1 植物材料 供试刺五加采自黑龙江省鸡西市,经河北联合大学生命科学学院邢朝斌副教授鉴定。分别以4-9月的叶和8月的根、茎、叶片和叶柄(编号为1-10)为提取总RNA的试材。
1.1.2 试剂 RevertAidTMFirst strand cDNA synthesis Kit购自Thermo公司。质粒小提试剂盒和琼脂糖凝胶DNA回收试剂盒购自Biomiga公司。TOP10感受态细胞、PGM-T克隆试剂盒和植物总RNA提取试剂盒购自天根生化科技(北京)有限公司。TaqDNA聚合酶和LATaqDNA聚合酶购自TaKaRa公司。X-Gal、IPTG、dNTPs、异硫氰酸胍购自北京拜尔迪生物技术有限公司。引物由生工生物工程(上海)股份有限公司合成,PAGE纯化。
1.2 方法
1.2.1 刺五加总RNA的提取与P450基因的获得 根据植物总RNA提取试剂盒的说明,分别提取1-10号刺五加样本的总RNA,并参照RevertAidTMFirst strand cDNA synthesis Kit的说明,以2 μL的总RNA为模板,将上述样本分别逆转录为cDNA。利用根据GenBank登录的与刺五加同科的人参(JX036031)、三七(GU997670)的P450基因序列设计的上游引物P4501S1:5'- ATGGATCTCTTTATCTCATC -3'和下游引物P4501X1:5'- ATGGATCTCTTTATCTCATC -3',以逆转录获得的刺五加叶片cDNA为模板,PCR扩增刺五加P450基因的cDNA序列。反应体系50 μL,其中上下游引物各1 μL,10×LATaqBuffer 5 μL,2.5 mmol/L dNTP 4 μL,模板cDNA 2 μL,LATaqDNA聚合酶0.6 μL,补ddH2O至50 μL。反应条件为:预变性95℃、3 min;变性94℃、30 s;退火50℃、30 s;延伸72℃、1 min 40 s。35个循环后72℃补充延伸10 min。扩增产物经1.5%琼脂糖凝胶电泳、回收、克隆入PGM-T质粒载体,转化大肠杆菌TOP10后,送Invitrogen公司测序。
1.2.2 刺五加P450基因的生物信息学分析 参照邢朝斌等[6]的方法,利用MotifScan、ProtParam、ProtScale、SOPMA和TMHMM 2.0等在线软件进行生物信息学分析。使用MEGA 5.21软件中的Neighbor-Joining法构建系统发育树。
1.2.3 刺五加P450基因的表达分析 以逆转录获得的1-10号的刺五加cDNA为模板,参照文献[6]的方法,利用预计扩增P450基因长度为139 bp的上游引物P450RTS:5'- TGGGTATCCAAAGAAGCAGG-3',下游引物P450RTX:5'-CAGCCAAGCCGAGTAGAGC -3',和预计扩增刺五加GAPDH基因长度为134 bp的引物RGS和RGX[6],进行表达量分析。
2 结果
2.1 刺五加P450基因的克隆与鉴定
利用引物P4501S1和P4501X1,RT-PCR扩增刺五加的cDNA,测序获得长1 410 bp的片段(图1),与预期长度相符。将扩增片段重组入PGM-T Vector后,PCR扩增产物的大小与刺五加总RNA的RTPCR产物大小相等。

图1 刺五加P450基因的克隆
2.2 刺五加P450基因的生物信息学分析
测序结果表明,所获得的刺五加P450基因cDNA的长度为1 410 bp,上游为起始密码子ATG,下游为终止密码子TAA,推测其编码469个氨基酸残基组成的蛋白质,GenBank登录号为KF498590。预测的蛋白质分子质量为53.083 kD,理论等电点(pI)为8.62。
通过NCBI的BLAST比对发现,刺五加P450蛋白与已知的蛋白质结构功能数据库中其他物种的
P450具有相似的结构功能域。刺五加P450的409-418位的氨基酸[FGGGPRMCPG]为P450家族的典型保守结构域,27-466位的氨基酸为P450的标志性序列。通过TMHMM软件分析得知,刺五加P450蛋白的5-21和276-292位氨基酸处存在跨膜螺旋,定位于线粒体。运用SOPMA软件预测刺五加P450蛋白的二级结构的结果表明,该蛋白含有222个α螺旋,占47.33%;61个延伸链,占13.01%;18个β折叠,占3.84%;168个无规则蜷曲,占35.82%。
2.3 P450的系统进化分析
利用MEGA软件,将GenBank中登载的12个物种的P450蛋白与刺五加的P450蛋白进行聚类分析,构建P450蛋白的系统进化树(图2)。刺五加与同为五加科的人参、三七和西洋参的亲缘关系最近,首先聚为一支,可信度达100%,之后与其他双子叶植物聚为一个大的分支,单子叶植物和裸子植物单独聚为一个分支,之后与人类聚在一起。

图2 P450氨基酸序列系统发育树
2.4 刺五加P450基因的表达分析
刺五加P450基因在不同生长发育时期和器官中的表达量变化如图3所示。自萌芽后的整个生长期中,P450基因均有表达,但表达量差异显著(P<0.05)。在整个生长期中,P450的表达呈现先低后高的变化趋势。萌芽期(4月26日)的表达量最低,之后显著升高,进入盛花期(6月26日)至叶片衰老期(9月26日)后,P450的表达量始终维持在较高水平。其中盛花期(6月26日)的表达量最高,是最低表达量萌芽期的1.26倍,但盛花期、果实快速生长期(7月26日)、果实基本成熟期(8月26日)和叶片衰老期的表达量间差异未达显著水平。
刺五加的P450基因在叶片、叶柄、幼茎和根中均有表达,但表达量具有显著差异(P<0.05)(图3)。叶片中的表达量最高,叶柄次之,分别为最低表达量(幼茎)的1.49和1.24倍,幼茎和根中的表达量间差异不显著。
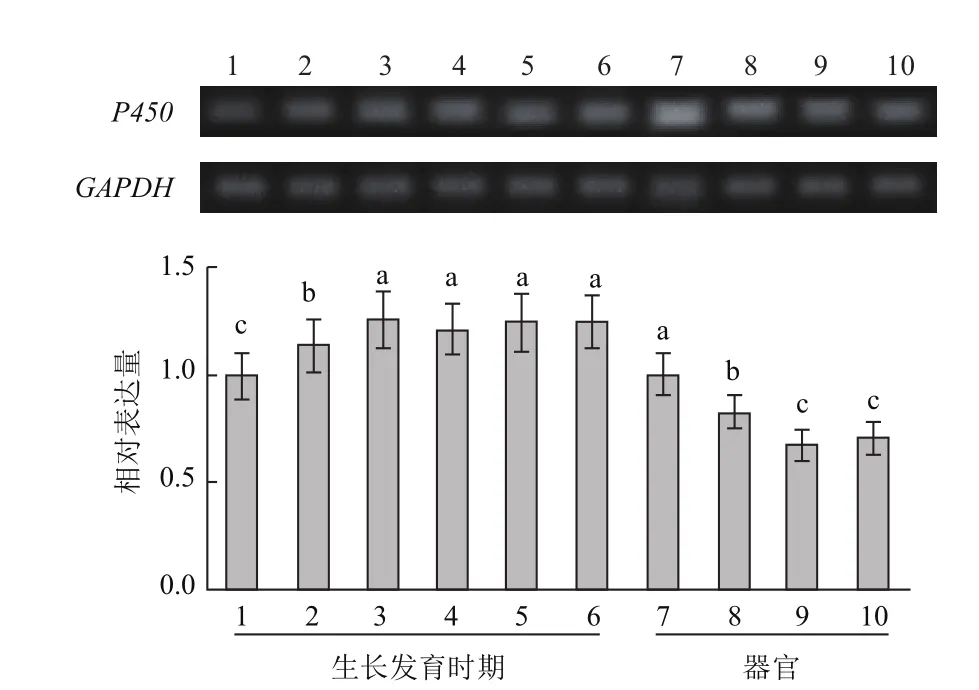
图3 刺五加不同生长发育时期及器官中P450基因的表达变化
3 讨论
细胞色素P450基因是一个古老的超基因家族,其编码蛋白为植物中最大的酶蛋白家族,在植物体内担当着重要的功能[7]。在所有已知结构的P450中都存在一个保守的血红素结合域,含有保守的FxxGxRxCxG序列,该序列是鉴定P450的主要特征,其中的半胱氨酸(C)是亚铁血红素的第5个轴配体,在所有的P450 中完全保守[7,8]。本试验所克隆出的刺五加P450基因的编码蛋白409-418位的氨基酸为[FGGGPRMCPG],与上述特征完全相符。氨基酸序列比对的结果显示,刺五加P450的氨基酸序列与人参(Panax ginseng)、三七(P. notoginseng)、西洋参(P. quinquefolius)和丹参(Salvia miltiorrhiza)的一致性分别达到91.5%、90.4%、89.3%和70.6%,依据“2个P450的氨基酸序列一致性如果大于
55%,则它们属于同一个亚家族”的原则[7],初步判定刺五加的P450属于P450家族中调控三萜皂苷类化合物形成的CPY85亚家族[9-11]。同时,依据P450的氨基酸序列进行的系统进化分析结果也与传统的分类结果相一致。
刺五加P450基因在不同生长发育时期和器官中表达量的分析结果表明,刺五加的P450基因在不同生长发育时期和叶片、叶柄、幼茎和根中均有表达,其表达量随着生长发育的进行逐渐上升,其中,盛花期的叶中的表达量最高。这与甘草[12]和蒺藜苜蓿[13]P450基因的表达特点相似。所不同的是甘草和蒺藜苜蓿的P450恒定高表达于根器官中,而刺五加中则为叶片中表达量最高,这与甘草[12]和蒺藜苜蓿[13]中皂苷类化合物主要存在于根,刺五加则主要存在于叶片中[1]的特点相符,这预示着P450在刺五加三萜皂苷类化合物的次生代谢过程中发挥着重要的作用。
4 结论
刺五加P450基因编码的蛋白与五加科其他植物P450的氨基酸序列一致性最高,并具有P450基因家族的典型特征序列。
刺五加的P450基因在不同生长发育时期和器官中均有表达,但表达量具有显著差异。
[1] 涂正伟, 周渭渭, 单淇, 等. 刺五加的研究进展[J]. 药物评价研究, 2011, 34(3):213-216.
[2] 赵云生, 万德光, 陈新, 等. 五环三萜皂苷生物合成与调控的研究进展[J]. 中草药, 2009, 40(2):327-330.
[3] 隋春, 徐洁森, 赵立子, 等. 北柴胡UGT基因的克隆及其过量表达和RNAi转基因载体的构建[J]. 中国中药杂志, 2012(5):558-563.
[4] 徐洁森, 魏建和, 赵立子, 等. 北柴胡细胞色素P450酶基因的克隆及其植物过量表达载体的构建[J]. 生物技术通讯, 2012, 23(4):537-541.
[5] Brazier M, Cole DJ, Edwards R. O-Glucosyltransferase activities toward phenolic natural products and xenobiotics in wheat and herbicide-resistant and herbicide-susceptible black-grass(Alopecurus myosuroides)[J]. Phytochemistry, 2002, 59(2):149-156.
[6] 邢朝斌, 龙月红, 何闪, 等. 刺五加法尼基焦磷酸合酶基因的克隆、 生物信息学及表达分析[J]. 中国中药杂志, 2012, 37(12):1725-1730.
[7] 贺丽虹, 赵淑娟, 胡之璧. 植物细胞色素P450基因与功能研究进展[J]. 药物生物技术, 2008, 15(2):142-147.
[8] Rupasinghe S, Schuler M. Homology modeling of plant cytochrome P450s[J]. Phytochem Rev, 2006, 5(2-3):473-505.
[9] Han JY, Hwang HS, Choi SW, et al. Cytochrome P450 CYP716A53v2 catalyzes the formation of protopanaxatriol from protopanaxadiol during ginsenoside biosynthesis in Panax ginseng[J]. Plant Cell Physiol, 2012, 53(9):1535-1545.
[10] Luo H, Sun C, Sun Y, et al. Analysis of the transcriptome of Panax notoginseng root uncovers putative triterpene saponin-biosynthetic genes and genetic markers[J]. BMC Genomics, 2011, 12(S5):1-15.
[11] Sui C, Zhang J, Wei J, et al. Transcriptome analysis of Bupleurum chinense focusing on genes involved in the biosynthesis of saikosaponins[J]. BMC Genomics, 2011, 12:539.
[12] Seki H, Sawai S, Ohyama K, et al. Triterpene functional genomics in licorice for identification of CYP72A154 involved in the biosynthesis of glycyrrhizin[J]. Plant Cell, 2011, 23(11):4112-4123.
[13] Carelli M, Biazzi E, Panara F, et al. Medicago truncatula CYP716A12 is a multifunctional oxidase involved in the biosynthesis of hemolytic saponins[J]. Plant Cell, 2011, 23(8):3070-3081.
(责任编辑 李楠)
Cloning and Expression Analysis of Cytochrome P450 Gene in Eleutherococcus senticosus
Xing Zhaobin Wu Peng Li Feifei Liu Yan Zhou Mi Xiu Leshan
(College of Life Science,Hebei United University,Tangshan 063000)
In order to clone cytochrome P450(P450)in Eleutherococcus senticosus, the primers were designed according to cDNA of the reported Panax ginseng and P. notoginseng. The full length cDNA of the E. senticosus P450 was cloned by RT-PCR, and analyzed the expression of P450 in different growth periods and organs of E. senticosus. The results showed that the full length of P450 was 1 410 bp, encoded a protein with 469 amino-acid residues, GenBank accession number KF498590. To compare the amino acid sequence of E.senticosus P450 with P. ginseng and P. notoginseng, the amino acid homology was 91.5% and 90.4%. P450 expressed in different growth periods and organs of E. senticosus, and the expression amount differed significantly(P<0.05). The highest content of the expression showed up when full opening flower stage, which was 1.26 times as much as that in the lowest at germination stage. The highest content of the expression was in the leaves which was 1.49 times as much as that of the lowest in young stem.
Eleutherococcus senticosus Cytochrome P450 Cloning Expression analysis
2013-09-18
国家自然科学基金项目(30701086),河北省自然科学基金项目(C2009001252),河北省自然科学基金-石药集团医药联合研究基金项目(H2012401006),河北联合大学培育基金项目(GP201306)
邢朝斌,男,副教授,研究方向:分子生药学、药用植物细胞工程;E-mail:xzbheuu@126.com
猜你喜欢
今日农业(2022年15期)2022-09-20
世界科学技术-中医药现代化(2021年12期)2021-04-19
中成药(2018年9期)2018-10-09
中成药(2017年9期)2017-12-19
中成药(2017年6期)2017-06-13
天然产物研究与开发(2016年1期)2016-06-05
中成药(2016年4期)2016-05-17
中成药(2016年4期)2016-05-17
中国药业(2014年16期)2014-05-14
中国民间疗法(2014年3期)2014-01-25
