
板栗抗性淀粉消化前后的益生作用及结构变化
2014-03-16 06:23杨春丰亢灵涛唐正辉
湖南工程学院学报(自然科学版) 2014年2期
杨春丰,亢灵涛,唐正辉,高 娟,谢 涛
(湖南工程学院 化学化工学院,湘潭 411104)
板栗抗性淀粉消化前后的益生作用及结构变化
杨春丰,亢灵涛,唐正辉,高 娟,谢 涛
(湖南工程学院 化学化工学院,湘潭 411104)
制备与纯化得到了板栗抗性淀粉及消化抗性淀粉,研究了它们的益生作用与结构变化.结果表明:板栗抗性淀粉及消化抗性淀粉对双歧杆菌和乳酸杆菌都有显著的增殖作用,对大肠杆菌和产气荚膜梭菌有强抑制作用,对粪肠球菌、梭状杆菌、兼性细菌无明显影响;它们的发酵液总酸度增大,说明它们能被肠道益生菌发酵利用;板栗抗性淀粉经消化处理后比表面积增加,经发酵后比表面积更大.板栗抗性淀粉的平均聚合度较之原淀粉显著变小,发酵后板栗抗性淀粉或消化抗性淀粉的平均聚合度降低;板栗抗性淀粉的晶型为V型,经消化后转变为B型,板栗抗性淀粉及消化抗性淀粉经发酵后,晶型都转变为A型,微晶度、亚微晶度及总结晶度较之发酵前都明显降低.
板栗;抗性淀粉;益生作用;结构变化
0 前 言
抗性淀粉(resistant starch)又称抗酶解淀粉、难消化淀粉,这种淀粉较其他淀粉难降解,在体内消化缓慢.抗性淀粉本身仍然是淀粉,其化学结构不同于纤维,但其性质类似溶解性纤维.抗性淀粉的定义为:在小肠中不能被酶解,但在人的肠胃道结肠中可以与挥发性脂肪酸起发酵反应的一类淀粉结构.淀粉不能被吸收并且能够进入结肠具有重要的生理功效.早在20世纪80年代初,国外就开始对抗性淀粉展开了研究,并认为这是一种与膳食纤维功能相似,但口感更优的一种新型益生元制剂,其生理功能及加工特征优良,具有广泛的应用前景[1,2].近些年来,国内外许多学者将研究的领域转向了非粮淀粉资源[3,4],且已有许多关于抗性淀粉的研究报道,但主要针对制备工艺的优化[4,5];而关于板栗抗性淀粉及其消化抗性淀粉的研究几无报道.因此,本文采用二次循环压热法制备并纯化得到了板栗抗性淀粉,再经人工胃液和人工胃肠液处理制得2个消化抗性淀粉样品,然后研究了这3个样品的益生作用及其结构变化,以期探讨消化前后的抗性淀粉在益生作用过程中可能的结构变化及更多的功能.
1 材料与方法
1.1 材料
选择性培养基:麦康凯琼脂培养基(大肠杆菌)、甘露醇氯化钠琼脂培养基(双歧杆菌)、LAMVAB琼脂培养基(乳酸杆菌)、麦芽糖琼脂培养基(粪肠球菌)、亚硫酸盐-环丝氨酸琼脂培养基(梭状杆菌)、甘露醇琼脂培养基(产气荚膜梭菌)、需氧-厌氧菌琼脂培养基购自美国Life Tech公司;耐热α-淀粉酶、糖化酶、胃蛋白酶、胰酶-5.0购自美国Sigma公司;其他试剂均为分析纯.
1.2 抗性淀粉的制备与纯化
[6]的方法制备和纯化得到板栗纯抗性淀粉(RS).
1.3 胃液、胃肠液消化抗性淀粉的制备
按参考文献[7]的方法制备人工胃液和人工胃肠液消化抗性淀粉,分别简称为DRSAGJ、DRSAGIJ.
1.4 体外厌氧发酵
1.4.1 培养基
改良MRS培养基:以2%的板栗抗性淀粉或板栗消化抗性淀粉取代MRS培养基中的葡萄糖作为碳源.选择性培养基:见1.1.
1.4.2 厌氧发酵
参照文献[7]方法进行厌氧发酵实验,在37℃厌氧培养至10h、20h、30h分别取样,用于肠道微生物菌群组成和总酸度的测定分析.
1.4.3 肠道菌群分析
取样品液1mL,用生理盐水稀释一定倍数后涂布于选择性平板上,于37℃厌氧培养48~72h后,使用全自动菌落分析仪计数.
1.4.4 发酵液总酸度测定
参照GB/T 12456-1990法测定每份发酵液的总酸度,并以乳酸含量表示.
1.4.5 发酵残余物制备
取10mL发酵液,离心(15min,3000g),沉淀用10mL蒸馏水清洗三次后烘干,粉碎过100目筛,这些发酵残余物分别称为发酵抗性淀粉(FRS)、发酵胃液消化抗性淀粉(FDRSAGJ)和发酵胃肠液消化抗性淀粉(FDRSAGIJ).
1.5 结构测定方法
平均聚合度(DP值)测定采用碘吸收法[8];扫描电子显微(SEM)分析按参考文献[9]的方法测定;X-射线衍射(XRD)分析采用参考文献[10]的方法.
所有数据为3个平行实验的平均值,且采用SPSS 20.0进行方差分析.
2 结果与分析
2.1 板栗抗性淀粉消化前后对肠道菌群生长的影响
以不添加板栗抗性淀粉或胃肠液消化抗性淀粉为对照,在10h、20h、30h取样,测定板栗抗性淀粉及胃肠液消化抗性淀粉对肠道菌群生长的影响结果见表1.从表1可知,发酵至第30h时各种肠道微生物菌浓均比发酵至第20h时的低,这是可能是30h时细菌生长已达衰老期,导致活细胞数目减少.当发酵20h后,板栗抗性淀粉及其胃肠液消化抗性淀粉对双歧杆菌和乳酸杆菌都有显著的增殖作用,其中抗性淀粉的增殖效果达到102倍、消化抗性淀粉的增殖效果更是高达近104倍左右;对大肠杆菌、粪肠球菌和产气荚膜梭菌的抑制作用也很强;对梭状杆菌的影响不明显.板栗胃肠液消化抗性淀粉比其抗性淀粉对双歧杆菌、乳酸杆菌增殖效果更加明显,对产气荚膜菌的抑制作用也更强些,说明抗性淀粉经消化道酶及其环境条件的作用是其发挥益生作用所必需的,也就是说消化抗性淀粉具有更好的益生作用,这与文献[11]、[12]报道的结果非常一致.

表1 板栗抗性淀粉消化前后对肠道菌群生长的影响(lg CFU/mL)
2.2 发酵液的酸度分析
发酵液的总酸度在一定程度上代表了益生菌尤其是双歧杆菌和乳酸杆菌的增殖情况,双歧杆菌和乳酸杆菌的增殖会促进丙酸、丁酸、乳酸等合成[13].表2为添加不同板栗抗性淀粉和胃肠液消化抗性淀粉发酵10h、20h、30h所测得的总酸度(以乳酸值表示).在整个发酵期间,板栗抗性淀粉及其胃肠液消化抗性淀粉的发酵液总酸度均比对照值要高出至少20%,说明抗性淀粉及其胃肠液消化抗性淀粉被肠道益生菌发酵过程中产生的短链脂肪酸数量增多,而且后者产生的脂肪酸数量要多些,这在文献[14]中也有报道.

表2 板栗抗性淀粉及消化抗性淀粉发酵液的总酸度(以乳酸表示,单位g/L))
2.3 发酵前后板栗抗性淀粉及消化抗性淀粉的超微结构变化
图1为发酵前后板栗抗性淀粉及其消化抗性淀粉的扫描电子显微照片.由图1A可看出,经压热制备的板栗抗性淀粉变为由大小、厚薄不一的片层堆积而成的叠层结构,且表面粗糙不平.造成这种变化的原因是:在糊化过程中,直链淀粉先从原淀粉颗粒中溶出,进入水中,冷却时,直链淀粉分子相互靠近依靠氢键及范德华力形成双螺旋并沉降,形成非常稳定的晶体;在后续纯化过程中,粗抗性淀粉中的无定形部分被酶解,剩下高度结晶的直链淀粉.板栗抗性淀粉经人工胃液消化后,叠层结构逐渐解体,剥落成大小、厚薄不一且无规堆积的碎片(图1B).板栗抗性淀粉经人工胃肠液联合作用后,大的碎片进一步细化、厚的碎片变薄,最终变成由大小不一的薄片疏松堆积的结构(图1C).发酵后板栗抗性淀粉由原来单一的叠层结构变为由少量片层与大量薄片混合堆积的结构,但薄片比例较高(图1D).人工胃液消化的抗性淀粉经发酵后,厚的碎片与大的薄片消失,变成以较小薄片为主的疏松结构(图1E).经人工胃肠液消化的抗性淀粉发酵后,薄片继续细化,最后变成由大小均匀的细小薄片堆积成的疏松结构(图1F).

图1 发酵前后板栗抗性淀粉及消化抗性淀粉的超微结构(放大3000倍)
2.4 发酵前后抗性淀粉、消化抗性淀粉的平均聚合度变化
图2为板栗抗性淀粉、消化抗性淀粉发酵前后的平均聚合度(DP).板栗抗性淀粉的DP值比其原淀粉的DP值减少了将近一半.这是由于在制备过程中,高温、高压使部分直链淀粉分子断裂,支链淀粉的部分支链断裂转化为直链淀粉,故而抗性淀粉的DP值要比其原淀粉的小;同时直链淀粉分子越大,越容易断裂.消化后板栗抗性淀粉的DP值比消化前几无降低,说明消化作用对抗性淀粉分子的DP值影响很小,而影响的主要是其结构.发酵后板栗抗性淀粉或消化抗性淀粉较之发酵前的DP值显著下降,但与抗性淀粉较之原淀粉DP值的降低幅度都要小得多.从图2还可看出,胃肠液消化过的板栗抗性淀粉经发酵后的DP值比板栗抗性淀粉经发酵后的DP值降低33.3%,由此进一步说明胃肠液的消化作用对抗性淀粉在肠道内的益生作用相当重要.
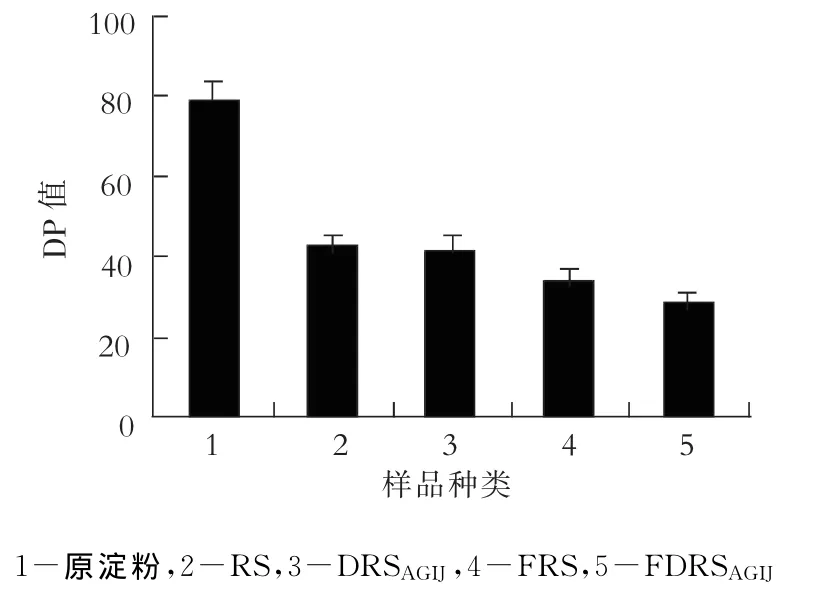
图2 板栗抗性淀粉及消化抗性淀粉发酵前后的DP值
2.5 发酵前后抗性淀粉、消化抗性淀粉的结晶度变化
淀粉是主要由A型和B型晶体及少量V型晶体组成的混合物.图3是板栗抗性淀粉及其消化抗性淀粉在体外厌氧发酵前后的X-射线衍射图谱,由其计算得到的结晶参数如表3所示.板栗抗性淀粉的晶型为V型,说明在制备抗性淀粉的过程中,其分子链通过断裂、重组,形成了更为稳定的晶体结构.板栗抗性淀粉经胃液或胃肠液消化后的晶型均转变为B型,与抗性淀粉相比,胃液消化抗性淀粉的微晶度、亚微晶度及总结晶度基本没有变化,但胃肠液消化抗性淀粉亚微晶度与总结晶度也有明显降低,说明经过人工胃液的强酸侵蚀与分散作用,增强了人工肠液的消化作用.板栗抗性淀粉及其消化抗性淀粉经发酵后,晶型都转变为A型,微晶度、亚微晶度及总结晶度较之发酵前都明显降低,说明发酵作用对板栗抗性淀粉及消化抗性淀粉的结晶结构有较大影响.

图3 板栗抗性淀粉及消化抗性淀粉发酵前后的X-射线衍射图谱
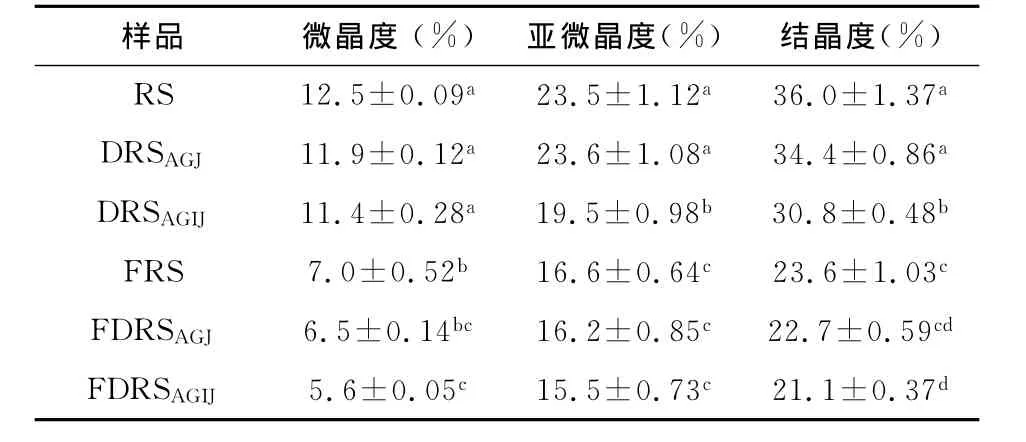
表3 板栗抗性淀粉及消化抗性淀粉发酵前后的结晶参数
3 结 论
(1)板栗抗性淀粉及消化抗性淀粉对双歧杆菌和乳酸杆菌都有显著的增殖作用,对大肠杆菌和产气荚膜梭菌有强抑制作用,对粪肠球菌、梭状杆菌无明显影响.它们的发酵液总酸度增大,说明它们被肠道益生菌发酵利用产生了更多的短链脂肪酸.
(2)板栗抗性淀粉经人工胃液、胃肠液消化处理后结构越来越疏松,且比表面积增加;板栗抗性淀粉及其消化抗性淀粉经发酵后,它们进一步碎化为以细小薄片为主体的有序结构,这种结构可能具有更大的比表面积和堆积密度,可能与抗性淀粉在肠道中发挥的其他生理功能有关.
(3)板栗抗性淀粉的DP值较之原淀粉显著变小.消化后板栗抗性淀粉的DP值比消化前几无降低,说明胃肠液主要影响抗性淀粉的结构.胃肠液消化过的板栗抗性淀粉经发酵后的DP值比板栗抗性淀粉经发酵后的DP值要小.
(4)板栗抗性淀粉的晶型为V型.经胃液或胃肠液消化后,板栗抗性淀粉的晶型均转变为B型,胃肠液消化抗性淀粉亚微晶度与总结晶度有所降低.板栗抗性淀粉及消化抗性淀粉经发酵后,晶型都转变为A型,微晶度、亚微晶度及总结晶度较之发酵前都明显降低.
参 考 文 献
[1]Rodríguez-Cabezas M E,Camuesco D,Arribas B,et al.The Combination of Fructoologosaccharides and Resistant Starch Shows Prebiotic Additive Effects in Rats[J].Clinical Nutrition,2010,29:832-839.
[2]Fuentes-Aaragozae E, Riquelme-Navarrete M J,Sánchez-Zapata E,et al.Resistant Starch as Functional Ingredient:a Review[J].Food Research International,2010,43:931-942.
[3]Jiranuntakul W,Puttanlek C,Rungsardthong V,et al.Microstructural and Physicochemical Properties of Heat-moisture Treated Waxy and Normal Starches[J].Journal of Food Engineering,2011,104:246-258.
[4]Sankhon A,Yao W R,Wang H Y,et al.The Yield of Improvement of Resistant Starches from Africa Locust(Parkia biglobosa):the Influence of Heat-moisture,Autoclaving-cooling and Cross-linking Treatments[J].American Journal of Food Technology,2012,7(7):386-397.
[5]李素玲.绿豆抗性淀粉的制备及性质研究[D].广州:华南理工大学硕士学位论文,2011,6.
[6]谢 涛,张 儒.绿豆抗性淀粉消化前后益生作用与结构变化的比较[J].现代食品科技,2013,29(12):2834-2839.
[7]曾红华,谢 涛,杨 莉,等.几种薯类与豆类抗性淀粉的抗消化性及其益生效应[J].中国粮油学报,2012,27(11):31-33.
[8]Takeda C,Takeda Y,Hizukuri S.Physicochemical Properties of Lily Starch[J].Cereal Chemistry,1983,60(1):212-216.
[9]Sushil D,Ashok K S,Michael J G.Effect of Cryomilling on Starches:Functionality and Digestibility[J].Food Hydrocolloids,2010,24(1):152-163.
[10]谢 涛,张淑远,王美桂,等.重结晶红薯淀粉体外消化前后益生作用与结构变化[J].农业机械学报,2013,44(8):203-208.
[11]Lopez-Rubio A,Flanagan B M,Gilbert E P.Molecular Rearrangement of Starch During in Vitro Digestion:Toward a Better Understanding of Enzyme Resistant starch Formation in Processed Starches[J].Biomacromolecules,2008,9(7):1951-1958.
[12]Mutungi C,Onyango C,Doert T,et al.Long-and short-range Structural Changes of Recrystallised Cassava Starch Subjected to in Vitro Digestion[J].Food Hydrocolloids,2011,25(2):477-485.
[13]Marzorati M,Verhelst A,Luta G,et al.In Vitro Modulation of the Human Gastrointestinal Microbial Community by Plant-derived Polysaccharide-rich Dietary Supplements[J].International Journal of Food Microbiology,2010,139(3):168-176.
[14]谢 涛,张 儒.锥栗直链淀粉-脂肪酸复合物的体外消化及益生元活性[J].中国粮油学报,2012,27(8):20-23.
Probiotic Functions and Structural Changes of Castanea Mollissima Resistant Starch and Its Products After Digestion and Fermentation
YANG Chun-feng, KANG Ling-tao, TANG Zheng-hui, GAO Juan, XIE Tao
(College of Chemical Engineering,Hunan Institute of Engineering,Xiangtan 411104,China)
Resistant starches from Castanea mollissima before and after in vitro digestion are made and purified.Their probotic functions and structural changes are intensively studied.The results demonstrate that these resistant starches before and after digestion have good proliferative effect on bifidobacterium and lactobacillus,have inhibitory impact on Escherichia coli and Clostridium perfringens,and have no obious influence on Enterococcus faecalis,Corynebacterium fusifome and facultative bacterial.The increasing total acidities of fermentation liquids demonstrate that these resistant starches before and after digestion can be used by intestinal probiotics.Compared with resistant starch,digested resistant starches have increasing specific surface areas,and have more greater specific surface areas after in vitro anaerobic fermentation.In comparison with native starch,average degree of polymerization of resistant starch decreases significantly.After fermentation,average degree of polymerizations of resistant starches before and after digestion decreases.Crystal type of resistant starch is type V,and then turns into type B after digestion.After fermentation,crystal types of resistant starches before and after digestion all become type A,and the degrees of microcrystalline,submicrocrystal and total crystallinity obviously decreases compared with before fermentation.
castanea mollissima;resistant starch;probotic function;structural change
TS235.2
A
1671-119X(2014)02-0061-05
2014-03-10
湖南省自然科学基金资助项目(2011JJ6009);国家级大学生创新创业训练计划项目(教高司函[2013]8号)
杨春丰(1992-),男,本科生,研究方向:淀粉化学与营养.
谢 涛(1970-),男,博士,教授,研究方向:再生资源与食品、生物化工.
猜你喜欢
公民与法治(2022年12期)2023-01-07
小学生作文(低年级适用)(2022年11期)2022-12-02
西南石油大学学报(自然科学版)(2019年5期)2019-12-20
创新作文(小学版)(2019年10期)2019-09-25
中成药(2018年10期)2018-10-26
小学生作文(中高年级适用)(2018年5期)2018-06-11
天然产物研究与开发(2018年4期)2018-05-07
中成药(2018年1期)2018-02-02
现代检验医学杂志(2016年1期)2016-11-12
兽医导刊(2016年12期)2016-05-17
