
九孔鲍(Haliotis diversicolor supertexta)耐低盐与生长性状的遗传参数评估*
2014-03-09 03:40刘建勇赖志服
海洋与湖沼 2014年3期
蒋 湘 刘建勇① 赖志服
(1.广东海洋大学水产学院 湛江 524025;2.汕尾市红海湾新海生养殖有限公司 汕尾 516600)
九孔鲍(Haliotis diversicolor supertexta)又称杂色鲍,自然分布于我国东南沿海海域,最北至日本、韩国海域,最南至菲律宾沿岸(Lindberg,1992),其营养价值与经济价值高,为海产八珍之一。中国的九孔鲍人工育苗养殖技术研究始于20世纪 70年代(柯才焕等,2011),90年代中期由于陆地工厂化养鲍技术的引进,九孔鲍迅速成为中国南方海域主要水产养殖品种之一(Nieet al,2004)。但是伴随着九孔鲍养殖业的发展,育苗和养成期暴发大规模病害,加之鲍鱼养殖企业不注重亲鲍选择与育苗工艺改进,持续近亲繁殖导致鲍鱼品种种质下降,对环境的适应力降低等,都给九孔鲍养殖业造成巨大的经济损失(宋振荣等,2000;徐力文等,2006;王江勇等,2007),严重制约了中国鲍鱼养殖业的发展。目前已有很多关于九孔鲍遗改良的研究报道,如种间杂交(柯才焕等,2000;蔡明夷等,2006),群体间远缘杂交(Youet al,2009;游伟伟等,2005),群体选育(Youet al,2010a)与分子标记辅助育种(Zhanet al,2009)。
估计遗传参数是为了更好制定育种策略,开展遗传育种研究。遗传参数主要包括遗传力、遗传相关与重复力,尤其是遗传力和遗传相关,对个体育种值估计、育种规划决策都起着十分重要的作用。有关水产动物遗传力的估计研究较多(郭华阳等,2011;Lucaset al,2006;Youet al,2010b;邓岳文,2005;孙长森等,2010),主要在一些数量性状方面,而与抗逆性状相关的遗传参数估计研究不多,王晓清等(2009)通过建立大黄鱼(Larimichthys crocea)15个半同胞家系,进行鱼苗对低盐、低溶氧和低pH值的抗性试验,并估计家系耐环境因子遗传力参数;张庆文等(2002)和杨翠华(2007)估计中国对虾(Fenneropenaeus chinensis)抗病群体的遗传参数,栾生等(2008)基于表型值和育种值对中国对虾生长、抗逆性状进行相关分析,国外学者研究凡纳滨对虾(Litopenaeus vannamei)对桃拉综合征病毒(TVS)抗逆性(Argueet al,2002),以及凡纳滨对虾(L.vannamei)对白斑综合征病毒(WSSV)抗逆性(Gitterleet al,2005)。有关鲍的抗逆性遗传参数估计研究未见报道,本研究根据不平衡巢式设计方法,建立 36个全同胞家系,利用线性动物模型与公母畜阈模型对九孔鲍生长与耐低盐性状的遗传参数进行评估,为九孔鲍育种项目的后续选育工作提供基础依据与参考。
1 材料与方法
1.1 实验材料
实验所用的九孔鲍家系材料培育于2011年10月,在广东省汕尾市红海湾经济开发区遮浪镇新海生养殖有限公司基地进行。所选亲鲍的表型性状:壳长为(5.46±0.47)cm,壳宽为(3.37±0.32)cm,体质量为(20.64±4.45)g。
1.2 实验设计
选择健康、性腺发育良好的 100只个体进行实验。选取性腺发育良好的12个雄性和36个雌性鲍,室内催产并采集精卵。通常雄鲍排精比雌鲍产卵要提前半个小时,将适量精卵混合进行授精,采用Comstock等(1952)建立的巢式交配设计,每个雄性鲍分别与3个雌性鲍交配,构建12个半同胞家系和36个全同胞家系,家系之间严格避免相互混杂,其它水质控制、养殖密度、饵料投喂和日常管理等操作,均按照标准化程序进行。当养殖到240日龄时,每个家系分别随机选取 40个,测量壳宽、壳长和体质量等生长性状。
耐低盐预实验,设定盐度梯度20,18,16,14和12,一个海水对照组,得到九孔鲍低盐度实验适宜盐度为16,选择盐度16来进行家系的耐低盐遗传参数估计。每个家系随机取活力较好的九孔鲍 40只,分为2组,每组20只,置于四角砖下,装入黑色塑料框(30cm×20cm×15cm)内,所有塑料框都做好标记避免混淆,全都置于大水泥池(7m×3m×1.35m)内养殖。保证养殖条件一致,提高实验准确度,24h充气,饲养48h。定时观察九孔鲍的活力并统计存活率。
1.3 数据统计分析
记录个体的性状,并按顺序排列。按照方差组分和遗传参数估计所使用软件 ASReml(Gilmouret al,2009)的要求,对数据进行整理和排列,系谱关系另建文件保存,表型参数由SPSS17.0软件包计算获得。
耐低盐性状的表示方法:实验结束后,以二进制数据来表示个体的存活状态,1为存活,0为死亡。做好记录,并统计家系存活率(Ødegårdaet al,2007)。
1.4 统计分析模型
建立无重复观测值的个体动物模型,对于每一个体生长性状的观察值y均可剖分为:
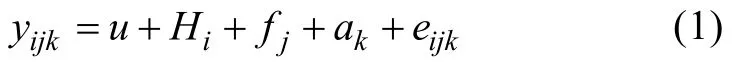
其中,i,j,k分别表示池号、全同胞组号、个体编号,yijk为性状观测值,u为总体平均值,Hi为池固定效应,fj为全同胞家系效应,ak为个体随机效应,eijk为随机残差效应,壳长(L)、壳宽(d)与体重(w)的遗传力计算公式为:
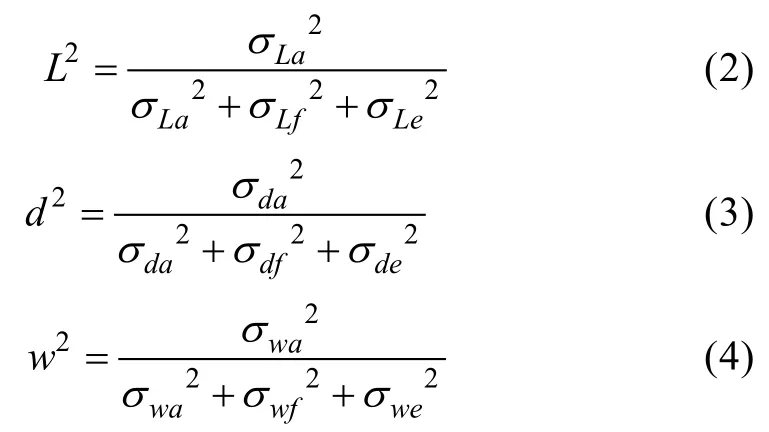
抗逆性实验结束后,统计所有实验个体的存活情况,得到各家系存活率,应用阈模型与广义线性混合模型分析方法(GLMM)估计存活性状的方差组分(logistic分布),育种分析模型为公母畜模型:
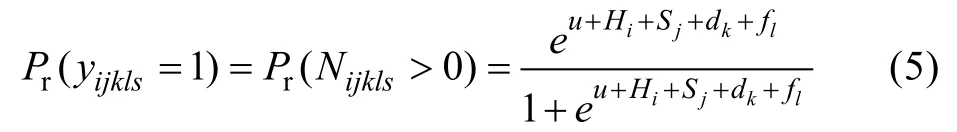
其中,l表示全同胞组号,yijkls为第s个体的存活状态,(1为存活,0为死亡),Nijkls为潜在变量,如果Nijkls>0,那么yijkls=1,如果Nijkls≤0,那么yijkls=0,Hi为池固定效应,Sj为公畜遗传效应,dk为母畜遗传效应。
存活性状遗传力计算公式:
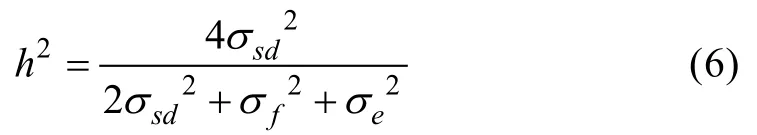
在Restricted Maximum Likelihood Method(REML)迭代过程中,选择不同的方差初始值重复计算,以保证达到整体而不是局部的最大化,收敛标准为 2次迭代所得估计值的方差小于1×10-6。比较最大似然函数在收敛后的似然函数值,取其中似然函数值最大的一次结果作为遗传方差组分估计值。
2 结果与分析
2.1 生长和耐低盐性状的描述性统计量
4个性状的变异系数(标准差与平均数的比值)范围为0.12—0.52,其中的耐低盐性状变异系数为0.52,变异系数较大;壳长,壳宽与体重变异系数较少,分别为0.13、0.12和0.35(表1)。图1以箱线图的形式给出九孔鲍36个家系的壳长,壳宽与体重的最大值、最小值、中位数、第一四分位数、第三四分位数与奇异值,同时列出各家系存活性状的条形图,由图可以看出家系之间生长性状的中位数差异较大,一般线性模型(GLM)分析表明家系间生长性状存在极显著差异(P<0.01);存活性状变化范围为0—0.83,家系间的差异极大。上述分析表明,九孔鲍养殖群体生长性状与耐低盐性状存在丰富的遗传变异,选择潜力高。
2.2 遗传参数估计
据统计 240日龄时九孔鲍在低盐度下生长性状遗传力与方差组分,壳长,壳宽,体重与耐低盐性状的遗传力分别为 0.18±0.04,0.13±0.06,0.18±0.15,0.056±0.022。在低盐度环境中,九孔鲍养殖群体耐低盐性状遗传力要低于生长性状的遗传力(表2)。
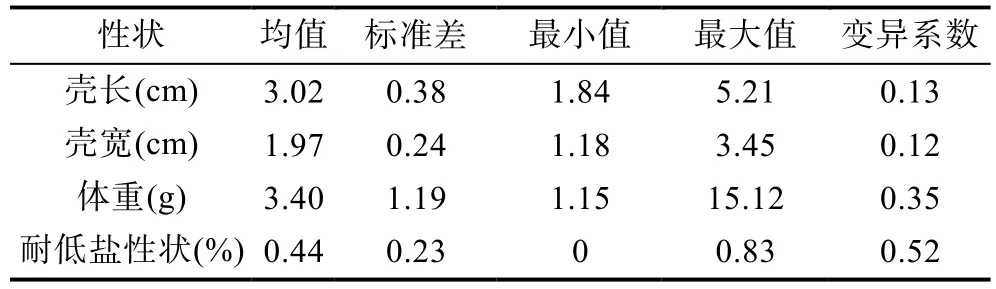
表1 日龄为240d的九孔鲍生长性状的表型参数Tab.1 Phenotypic parameter of growth traits of H.diversicolor supertexta
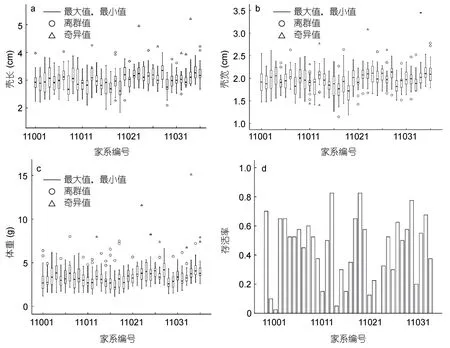
图1 九孔鲍各家系主要生长性状的箱线图(a—c)与耐低盐存活率条形图(d)Fig.1 Box plots of growth traits(a—c) and bar charts of salinity tolerance survival rate(d) in all H.diversicolor aquatili falimiles
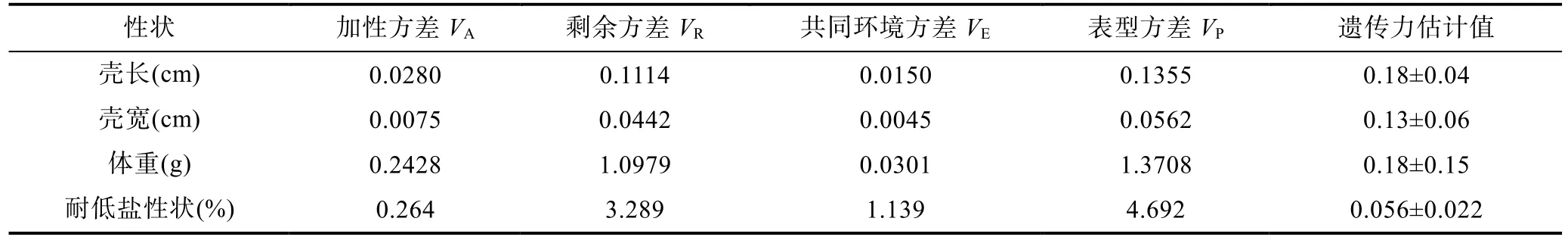
表2 九孔鲍家系生长与耐低盐性状的方差组分与遗传力Tab.2 Heritability and variance components of growth and low salinity tolerance traits of H.diversicolor supertexta
2.3 表型性状间的表型相关与遗传相关
壳长,壳宽与体重3个主要生长性状与耐低盐性状的表型相关与遗传相关系数分别见表3、表4,壳长,壳重与体重之间的相关系数为 0.76—0.91,为极显著正相关(P<0.01),但它们与耐低盐性状之间都呈不显著的负相关;遗传相关与表型相关变化一致,表明抗逆性状与生长性状间无显著的相关关系,从壳长,壳宽与体重3个性状中任意一个进行选择都不能达到间接选育群体抗逆性的目的。

表3 九孔鲍各性状间的表型相关Tab.3 Phenotypic correlations among all traits of H.diversicolor supertexta

表4 九孔鲍各性状间的遗传相关Tab.4 Genetic correlations among all traits of H.diversicolor supertexta
3 结论
由表1可见,九孔鲍壳长,壳宽与体重的变异系数均较大,其中低盐条件下存活率变异系数最大,达到0.52。实验中各家系均在相同环境下养殖,造成成活率差异的原因主要是个体的遗传效应,这表明耐低盐性状具较高选育潜力。许多研究表明,贝类的主要经济性状在不同家系间存在着显著的差异,如 Liu等(2011)在相同养殖环境下对海湾扇贝 12个同胞家系进行养殖,家系间的体重,壳长与存活表现出显著差异,Wang等(2010)通过巢式设计建立36个墨西哥湾扇贝(Argopecten irradians concentricus Say)全同胞家系,研究结果发现家系间体重差异显著,他们认为造成这种现象的原因主要是个体的遗传效应。
国内外关于鲍数量性状遗传参数估计研究报道较多,Lucas 等(2006)、You 等(2010b)、邓岳文,(2005)等分别估计了耳鲍(Haliotis asinina)、杂色鲍(Haliotis diversicolor)与皱纹盘鲍(Haliotis discus hannai)不同生长阶段主要生长性状的遗传力,其估计值为0.14—0.48.本研究中壳长、壳宽与体重的遗传力分别为 0.18±0.04,0.13±0.06,0.18±0.15,明显低于上述近缘类群的估计值。同一物种不同性状间遗传力差别较大;而对于同一性状,不同的研究群体、不同的遗传评估方法或遗传模型均会对遗传力的评估结果造成影响。本研究在遗传评估模型中考虑了共同环境效应,且发现该效应所占总方差组分较高,平均达到11.38%,这可能是造成本研究中遗传力估计值偏低的主要原因。由于实验条件限制,许多水产动物的家系在进行标记前,是单独培育的,共同环境效应是由全同胞家系单独养殖而引起的环境效应,属非加性遗传,在遗传分析中应当从加性效应中剔除。
有关水产动物抗逆性状遗传力的报道较少,极端条件(如高温、高盐等)下的存活率或存活时间常被作动物抗性大小的指标(Argueet al,2002;Gitterleet al,2005;栾生等,2008)。存活性状在遗传学上被划分为阈性状,是一类表型符合二项分布(存活和死亡),但在遗传上由多基因决定的复杂性状,其遗传参数的估计有多种,已在水产动物中应用的有线性动物模型、公母畜阈模型与方差分析等方法(王晓清等,2009;栾生等,2012),殷宗俊等(2005)认为线性思想下阈性状遗传分析得不到理想效果,利用广义线性混合模型(GLMM)方法和公母畜阈模型进行分析更为合适(Nielsenet al,2010;Ødegårdaet al,2007;Ødegårdet al,2010)。本实验利用公母畜阈模型估计了九孔鲍养殖群体耐低盐性状的遗传力,估计值为0.056±0.022,属低遗传力。许多水产动物抗逆性状的遗传力估计值均较低,如:尼罗罗非鱼(Oreochromis niloticus)(Rezket al,2009;Charo-Karisaaet al,2006),存活性状遗传力的估计值分别为 0.12±0.0346,0.03—0.14;王晓清等(2009)估计了大黄鱼(Larimichthys crocea)耐环境因子的遗传参数,耐低盐、低溶氧和低pH值的遗传力分别为0.23,0.10,0.23;在中国对虾(栾生等,2008),凡纳滨对虾(L.s vannamei)(Argueet al,2002;Gitterleet al,2005)的抗逆性选育研究中,存活率性状的遗传力范围为 0.03—0.16,都表现为中低遗传力水平,这些结果与本研究相类似。高遗传力的性状适宜用群体选育的方法来进行育种,而对于低遗传力性状则用家系选育较为合适,可见,九孔鲍耐低盐性状选育宜采用家系选育方法,另外,低遗传力意味着进行遗传改良时,需要更多的世代和家系,每个家系测定更多的个体。
在育种工作中,性状间的遗传相关与表型相关参数常用来辅助选种过程,当通过直接选育难以达到要求,或不方便直接对某一性状进行选育时,可以选另外一个与目标性状有着高遗传相关性状来完成选种。本研究结果表明九孔鲍生长性状与抗逆性状的相关关系不显著。许多研究结果表明,水产动物的生长与抗逆性状的相关性较小或没有相关关系,如:Krishna等(2011)在斑节对虾(Penaeus monodon)的遗传参数估计中,发现收获体重与存活率的相关系数为 0.05(P>0.05),刘宝锁等(2011)大菱鲆(Scophthalmus maximus)生长和耐高温性状的相关系数仅为 0.04(P>0.05),栾生等(2008)对中国对虾的抗逆性和生长性状进行相关分析,发现二者无显著相关关系,张天时(2010)对中国对虾育种分析模型与遗传参数评估,发现抗 WSSV存活性状与生长性状遗传相关系数仅为 0.038,且不显著。以上研究结果与本研究相类似,可见,在进行九孔鲍育种规划时,有必要采用经济加权系数或百分比赋值方式,对抗逆性状进行一定的加权,制定多性状综合选择指数,据此评估和选择优秀的留种亲本,以加快育种进程。
王江勇,郭志勋,冯 娟等,2007.养殖杂色鲍暴发病超微病理学研究.海洋科学,31(3):28—32
王晓清,王志勇,何湘蓉,2009.大黄鱼(Larimichthys crocea)耐环境因子试验及其遗传力的估计.海洋与湖沼,40(6):781—785
邓岳文,2005.皱纹盘鲍数量遗传与育种研究.青岛:中国科学院海洋研究所博士学位论文,1—33
刘宝锁,张天时,孔 杰等,2011.大菱鲆生长和耐高温性状的遗传参数估计.水产学报,35(11):1601—1606
孙长森,林志华,董迎辉等,2010.泥蚶(Tegillarca granosa)主要经济性状遗传参数的估算.海洋与湖沼,41(6):907—913
杨翠华,2007.中国对虾与抗性相关性状的遗传学参数分析.青岛:中国海洋大学 博士学位论文
宋振荣,纪荣兴,颜素芬,2000.引起九孔鲍大量死亡的一种球状病毒.水产学报,24(5):463—466
张天时,2010.中国对虾(Fenneropenaeus chinensis)育种分析模型与遗传参数评估.青岛:中国海洋大学 博士学位论文
张庆文,刘 萍,王伟继等,2002.中国对虾抗病群体选育的初步研究.海洋水产研究,23(2):53—57
柯才焕,田 越,周时强等,2000.杂色鲍与皱纹盘鲍盘鲍杂交的初步研究.海洋科学,24(11):39—41
柯才焕,游伟伟,2011.杂色鲍的遗传育种研究进展.厦门大学学报:自然科学版,50(2):425—430
徐力文,王江勇,陈毕生,2006.我国南方鲍鱼养殖业的困境与发展探讨.湛江海洋大学学报:自然科学版,26(4):100—104
殷宗俊,张 勤,2005.利用GLMM方法估计家畜阈性状的遗传力.中国农业大学学报,10(6):33—36
栾 生,孔 杰,张天时等,2008.基于表型值和育种值的中国对虾生长、抗逆性状相关分析.海洋水产研究,29(3):14—20
栾 生,边文冀,邓 伟等,2012.斑点叉尾基础群体生长和存活性状遗传参数估计.水产学报,36(9):1313—1321
郭华阳,张殿昌,李恒德等,2011.合浦珠母贝幼体生长性状的遗传力及其相关性分析.湖北农业科学,50(21):4441—4444
游伟伟,柯才焕,蔡明夷等,2005.杂色鲍日本群体与台湾群体杂交的初步研究.厦门大学学报:自然科学版,44(5):700—705
蔡明夷,柯才焕,王桂忠等,2006.杂色鲍与盘鲍种间杂交受精率的影响因素.中国水产科学,13(2):28—32
Argue B J,Arce S M,Lotz J Met al,2002.Selective breeding of Pacific white shrimp(Litopenaeus vannamei) for growth and resistance to Taura Syndrome Virus.Aquculture,204(3—4):447—460
Charo-Karisaa H,Komen H ,Rezk M Aet al,2006.Heritability estimates and response to selection for growth of Nile tilapia(Oreochromis niloticus) in low-input earthen ponds.Aquculture,261(2):479—486
Comstock R E,Robinson H F,1952.Estimation of average dominance of genes.Ames,IA:Iowa State College Press,494—516
Gilmour A R,Gogel B J,Cullis B Ret al,2009.ASREML User's Manual Guide Release 3.0,Hemel Hempstead HP1 1ES,UK:VSN international Ltd
Gitterle T,Salte R,Gjerde Bet al,2005.Genetic(co)variation in resistance to White Spot Syndrome Virus(WSSV) and harvest weight inPenaeus Litopenaeus vannamei.Aquculture,246(1—4):139—149
Krishna G,Gopikrishna G,Gopal Cet al,2011.Genetic parameters for growth and survival inpenaeus monodoncultured in India.Aquaculture,318(1—2):74—78
Lindberg D R,1992.Evolution,didtribution and systematics ofHaliotiade.In:Shepherd S A,Tegner M J,Guzman del Proo S A ed.Abalone of the world:biology,fisheries and culture.Blackwell Scientific Publishing,London:3—18
Liu J Y,Liu Z G,Sun X Z,2011.The Effects of Inbreeding on Production Traits of the Southern Bay ScallopArgopecten irradians concentricus.Journal of Shellfish Research,30(1):109—113
Lucas T,Macbeth M,2006.Heritability estimates for growth in the tropical abaloneHaliotis asininausing microsatellites to assign parentage.Aquaculture,259(1—4):146—152
Nie Z Q,Wang S P,2004.The status of abalone culture in China.Journal of Shellfish Research,23:941—946
Nielsen H M,Ødegård J,Olesen Iet al,2010.Genetic analysis of common carp(Cyprinus carpio) strains I:Genetic parameters and heterosis for growth traits and survival.Aquculture,304:14—21
Ødegård J,Meuwisse T H,Heringstad Bet al,2010.A simple algorithm to estimate genetic variance in an animal threshold model using Bayesian inference.Genetics Selection Evolution,42:29—35
Ødegårda J,Olesenb I,Gjerde Bet al,2007.Evaluation of statistical models for genetic analysis of challenge-test data on ISA resistance in Atlantic salmon(Salmo salar):Prediction of progeny survival.Aquculture,266(1—4):70—76
Rezk M A,Ponzoni R W,Khaw H Let al,2009.Selective breeding for increased body weight in a synthetic breed of Egyptian Nile tilapia,Oreochromis niloticus:Response to selection and genetic parameters.Aquculture,293(3—4):187—194
Wang H,Du X D,Liu Z Get al,2010.Estimating the heritability for growth-related traits in the pearl oyster,Pinctada fucata martensii(Dunker).Aquaculture Research,42(1):57—64
You W W,Ke C H,Luo X,2009.Growth and survival of three small abaloneHaliotis diversicolorpolulations and their reciprocal crosses.Aquaculture Research,40(13):1474—1480
You W W,Ke C H,Luo X,2010a.Divergent selection for shell length in two stocks of small abalone,Haliotis diversicolor.Aquaculture Research,41(6):921—929
You W W,Ke C H,Luo Xet al,2010b.Heritability of growth traits for small abalone estimated from sib matings.Journal of Shellfish Research,29(3):705—708
Zhan X,Hu H Y,Ke C H,2009.Isolation and characterization of eleven microsatellite loci in small abalone,Haliotis diversicolor.Conservation Genetic,10(4):1185—1187
猜你喜欢
养猪(2022年4期)2022-08-17
农民致富之友(2019年31期)2019-11-23
爱你(2018年24期)2018-08-16
爱你·阳光少年(2018年8期)2018-05-14
现代交际(2016年20期)2017-02-15
中国照明(2016年4期)2016-05-17
湖北畜牧兽医(2015年11期)2016-01-11
湖北畜牧兽医(2015年11期)2016-01-11
医学研究杂志(2015年9期)2015-07-01
中国果菜(2015年2期)2015-03-11
