
低温胁迫对6种珍贵树种苗木光合荧光特性的影响
2014-02-28 08:47张毅龙张卫强甘先华
生态环境学报 2014年5期
张毅龙,张卫强,甘先华
1. 华南农业大学林学院,广东 广州 510642;2. 广东省林业科学研究院,广东 广州 510520
低温胁迫对6种珍贵树种苗木光合荧光特性的影响
张毅龙1,张卫强2*,甘先华2
1. 华南农业大学林学院,广东 广州 510642;2. 广东省林业科学研究院,广东 广州 510520
以格木(Erythrophleum fordii)、降香黄檀(Dalbergia odorifera)、闽楠(Phoebe bournei)檀香 ( Santalum album)、铁刀木(Cassia siamea)和樟树(Cinnamomum camphora)6种珍贵树种苗木为研究对象,分析自然降温过程中苗木相对叶绿素含量、气体交换参数及叶绿素荧光参数等指标的变化,探讨低温胁迫对这6种苗木光合荧光特性的影响,并利用隶属函数法对6种珍贵树种耐寒能力进行了评价,以期了解这几种植物的抗寒能力差异及低温对其光合能力的影响,为扩大引种和栽培提供依据。结果表明,与对照相比,低温胁迫下6种珍贵树种苗木净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)均有不同程度的下降,其中,格木、降香黄檀、闽楠、檀香、铁刀木净光合效率的下降很大程度上来源于非气孔因素抑制,而樟树净光合速率下降可能来源于气孔限制因素。闽楠、檀香和樟树水分利用效率(WUE)呈上升趋势,而格木与降香黄檀WUE均有不同程度的降低,表明闽楠、檀香和樟树能较好地协调碳同化和水分耗散。低温胁迫下6种珍贵树种Fm均有所下降,表明低温可对珍贵树种苗木叶片PSII反应中心的电子传递潜力产生明显的抑制,致使珍贵树种PSII的光能转换效率降低。降香黄檀、闽楠、檀香、铁刀木和樟树经历2次低温胁迫后qP呈现出持续下降的趋势,表明珍贵树种PSII反应中心的开放程度降低。在2次低温胁迫间格木、降香黄檀、闽楠和铁刀木qN差异不显著(p>0.05),表明珍贵树种具有过剩光能耗散机制,保护光合机构免受破坏。相关性分析结果表明,Pn与Gs、Tr、Fv/Fm、qP、qN、ETR和Fm存在极显著相关关系(p<0.01),表明Gs、Tr、Fv/Fm、qP、qN、ETR和Fm均可作为衡量植物光合能力的参数。隶属函数法分析结果表明,6种珍贵树种抗寒能力依次为格木>樟树>闽楠=檀香=铁刀木>降香黄檀。总之,低温胁迫破坏了这6种苗木光合机构可能是导致植物光合能力降低的主要原因,且不同植物对低温胁迫的响应程度不同,这是扩大引种和栽培需要考虑的因素。
低温胁迫;气体交换参数;叶绿素荧光参数;抗寒能力评价;抗寒筛选
格木(Erythrophleum fordii)、降香黄檀(Dalbergia odorifera)、闽楠(Phoebe bournei)、檀香(Santalum album)、铁刀木(Cassia siamea)和樟树(Cinnamomum camphora)是热带、亚热带地区常见的珍贵树种,在林业生态工程建设中占重要地位。尽管已有对不同树种低温胁迫的生理生化响应的相关研究(王宁等,2013;林宁等,2012;贺磊等,2011;郭文福和贾宏炎,2006)。但对自然极速降温过程中珍贵树种光合生理生态变化的研究却少有涉及。本研究以格木、降香黄檀、闽楠、檀香、铁刀木和樟树6种珍贵树种苗木为研究对象,分析了自然降温过程中苗木叶片相对叶绿素含量、气体交换参数及叶绿素荧光参数等指标的变化,探讨了低温胁迫对6种珍贵树种苗木光合荧光特性的影响,分析了6种珍贵树种耐寒能力,以期为扩大引种和栽培提供理论依据。

表1 6种苗木的基本情况(平均值±标准误)Table 1 Characteristics of seedlings of six examined plant species (mean ± SD)
1 材料与方法
1.1 研究区概况
试验地设在广东省林业科学研究院(广州)苗圃内,位于23°14'N,113°23'E,海拔25 m,典型亚热带季风气候,年平均温度23 ℃,最低月平均气温13.3 ℃(1月),最高月平均气温38.1 ℃(8月),年降水量1638 mm,4~9月份的降水量占全年的80%,年平均湿度79%。
1.2 试验材料
6种参试苗木为:格木、降香黄檀、闽楠、檀香、铁刀木和樟树,均是热带、亚热带地区常见的阔叶乔木,喜温暖气候,对热量条件要求较高。选择生长健康的3 a生实生苗栽培于营养袋中,袋子高60 cm,袋口径为37 cm,袋中土为沙壤土,保持水肥管理一致。每个营养袋栽培1株实生苗,每个树种栽培3株(表1)。
1.3 研究方法
1.3.1 低温胁迫处理
2013年11月22日-24日平均气温为19.5 ℃,最低温为13 ℃,此温度为对照;2013年12月30日-2014年01月01日平均气温为12 ℃,最低温为4 ℃,此温度为低温胁迫Ⅰ(简称胁迫Ⅰ);2014年02月10日-14日平均气温为7.1 ℃,最低气温为4 ℃,此温度为低温胁迫Ⅱ(简称胁迫Ⅱ)。
1.3.2 相对叶绿素含量的测定
每株树选取3片冠层阳面的健康成熟的功能叶作为测试叶,分别挂牌标记,测定时保持叶片自然生长角度不变(下同)。采用美国CCM-200的手持式叶绿素计,对选定的苗木叶片进行叶绿素相对含量测定。每一处理重复测定3次。
1.3.3 气体交换参数的测定
在对照和低温胁迫(Ⅰ和Ⅱ)环境下,采用LI-6400便携式光合作用测定系统外加6400-2B LED红蓝光源测定了1500 μmol·m-2·s-1光合有效辐射强度下珍贵树种气体交换参数。指标包括:净光合速率(Pn,μmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)、蒸腾速率(Tr,mmol·m-2·s-1)、光合有效辐射(PAR,μmol·m-2·s-1)、大气CO2浓度(Ca,μmol·m-2·s-1)、大气温度(Ta,℃)、叶片温度(Tl,℃)、空气相对湿度(RH,%)等。叶片水分利用效率(WUE,μmol·m-2·s-1)由公式WUE=Pn/Tr计算。测定时间均为上午9:00~11:00,连续测定3 d。
1.3.4 叶绿素荧光参数的测定
在测定珍贵树种气体交换参数的同时进行叶绿素荧光参数的测定。日出前,待测叶片用锡箔包被进行暗处理。叶片暗适应20 min后利用配置调制荧光叶室(Li-6400-40)的便携式光合测定系统LI-6400XT(Li-COR,USA)对待测叶片进行测定,先测定初始荧光(Fo)、最大荧光(Fm)、最大PSⅡ的光能转换效率(Fv/Fm),然后打开活化光(1500 μmol·m-2·s-1)测定光化学淬灭(qP)、非光化学淬灭(qN)和表观量子传递速率(ETR)等。测定时间均为上午9:00~11:00,连续测定3 d。
1.4 数据处理与分析
数据统计分析和作图由Excel和SPSS16.0软件完成,用Duncan多重比较检验苗木各测量指标在不同温度处理间的差异显著性。各指标抗寒系数=
低温胁迫下的指标测定值/对照指标测定值(许桂芳等,2009)。抗寒性综合评价:采用数学分析隶属函数法(许桂芳等,2009;赵一鹤等,2012;亓白岩等,2013)对测定的各项指标进行转换和综合分析评价。
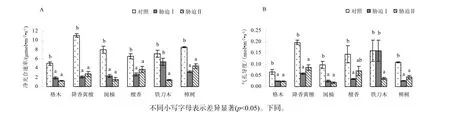
图1 低温胁迫对幼苗叶片净光合速率(Pn) 和气孔导度(Gs)的影响Fig.1 Effect of low temperature stress on the net photosynthetic rate (Pn) and stomatal conductance(Gs)in seedling leaves of the examined plants
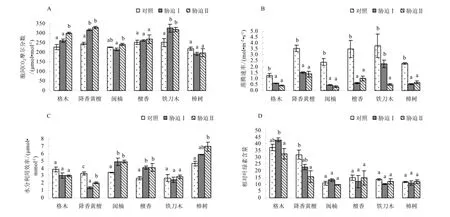
图2 低温胁迫对幼苗叶片胞间CO2摩尔分数(Ci)、蒸腾速率(Tr)、水分利用效率(WUE)和相对叶绿素含量的影响Fig.2 Effect of low temperature stress on the intercellular CO2, transpiration rate concentration(Ci), transpiration rate(Tr), water use efficiency (WUE) and relative chlorophyll content in seedling leaves of the examined plants
2 结果与分析
2.1 低温胁迫对珍贵树种气体交换参数的影响
与对照相比,低温胁迫Ⅰ和Ⅱ下6个树种的净光合速率(Pn)均有不同程度的下降(图1A),下降为对照的19.1%~75.4%;其中降香黄檀降幅最大,胁迫Ⅰ和胁迫Ⅱ分别下降为对照的19.1%和24.8%。方差分析表明,对照处理的格木、降香黄檀、闽楠、檀香和樟树Pn显著高于低温胁迫(Ⅰ和Ⅱ)(p<0.05),而在低温胁迫(Ⅰ和Ⅱ)处理间Pn差异不显著(p>0.05)。气孔导度(Gs)变化趋势与净光合速率基本一致,与对照相比均有所下降(图1B),其中,低温胁迫Ⅰ下格木、降香黄檀、闽楠、檀香和樟树的Gs与对照相比下降显著(p<0.05),分别下降为对照的36.4%、29.3%、25.4%、23.6%和 23.4%,而檀香和铁刀木Gs与对照相比差异不显著(p>0.05)。
从图2A可知,格木、降香黄檀、檀香和铁刀木胞间CO2摩尔分数(Ci)在低温胁迫期间呈上升趋势,其中,降香黄檀和铁刀木Ci显著高于对照(p<0.05),而闽楠和樟树Ci在不同处理间差异不显著(p>0.05)。受低温胁迫后,6种珍贵树种蒸腾速率(Tr)均有不同程度的下降。其中,格木、降香黄檀、闽楠、檀香、樟树的蒸腾速率(Tr)显著低于对照Tr(p<0.05),而在低温胁迫Ⅰ和Ⅱ间Tr差异不显著(p >0.05)(图2B)。在低温胁迫过程中,闽楠、檀香和樟树水分利用效率(WUE)呈上升趋势,而格木与降香黄檀WUE均有不同程度的下降(图2C)。
2.2 低温胁迫对珍贵树种相对叶绿素含量的影响
由图2D可知,闽楠、檀香、铁刀木和樟树的相对叶绿素含量(WUE)在对照、低温胁迫Ⅰ和低温胁迫Ⅱ间差异不显著(p>0.05),低温胁迫对以上树种叶片相对叶绿素含量影响不大;格木的相对叶
绿素含量在低温胁迫期间表现为先升高后下降的趋势,而降香黄檀相对叶绿素含量则呈现出持续下降的趋势。

图3 低温胁迫对苗木初始荧光(Fo)和最大荧光(Fm)的影响Fig. 3 Effect of low temperature stress on Foand Fmin seedling leaves of the examined plants
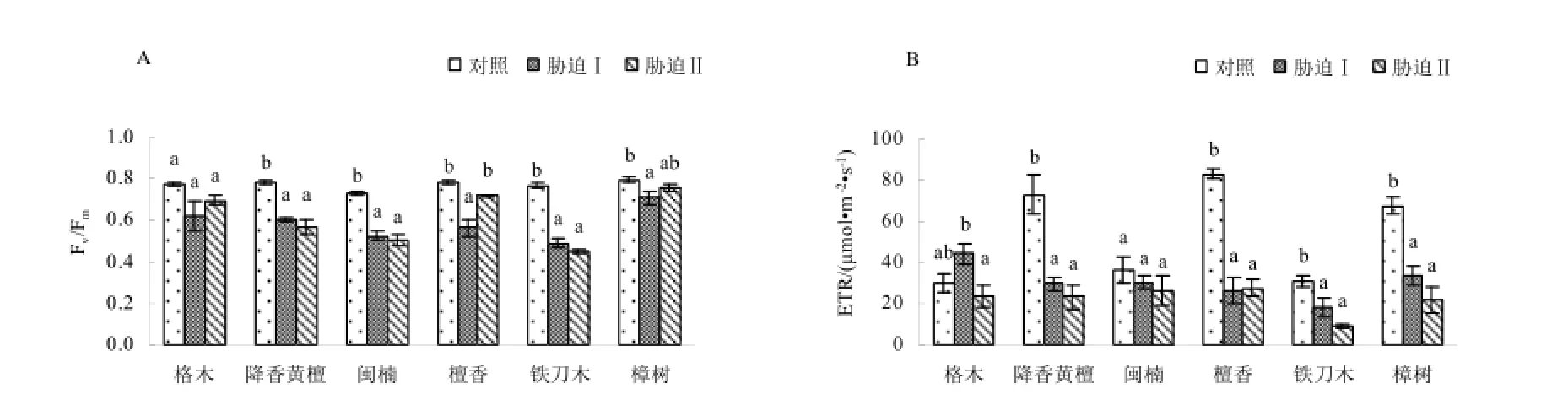
图4 低温胁迫对苗木最大PSⅡ的光能转换效率(Fm/Fv)和表观量子传递速率(ETR)的影响Fig.4 Effect of low temperature stress on Fm/Fvand ETR in seedling leaves of the examined plants
2.3 低温胁迫对珍贵树种叶绿素荧光特性的影响
由图3A可知,与对照相比,在低温胁迫Ⅰ下格木、降香黄檀、檀香、铁刀木和樟树的Fo均有不同程度的升高;其中檀香、铁刀木和樟树的Fo升高显著(p<0.5),增幅介于15.92%~37.00%之间;其中又数檀香增幅最高,表明PSⅡ反应中心受到不可逆转的失活或破坏(周蕴薇等,2006)。格木和降香黄檀的Fo上升不显著(p>0.5)。在低温胁迫Ⅱ下闽楠和铁刀木的Fo均有所下降,下降幅度分别为6.45%和25.07%,可能是低温使铁刀木叶片光合色素降解导致Fo下降(Mikkelsen,1996)。最大荧光(Fm)是已经暗适应的光合机构全部的PSⅡ中心都关闭时的荧光强度,通常叶片经暗适应20 min后测定,这时所有的非光化学过程都最小,是标准的最大荧光,可反映PSⅡ电子传递情况(Baker,2008)。与对照相比,低温胁迫下6种珍贵树种Fm均有不同程度的下降,这表明低温对珍贵树种苗木叶片PSII反应中心的电子传递潜力产生了明显的抑制。其中,降香黄檀、闽楠、铁刀木的Fm下降显著(p<0.05),低温胁迫Ⅰ和Ⅱ分别降为对照的68.24%和56.89%、60.22%和51.02%、57.64%和34.04%,而樟树Fm下降不显著(p>0.05),两种胁迫分别下降为对照的86.10%和95.82%(图3B)。
最大PSⅡ的光能转换效率(Fv/Fm)是表征健康植物叶片的指标(Maxwell和Johnson,2000)。从图4A可知,在对照下6种珍贵树种Fv/Fm介于0.73~0.79之间,说明在非胁迫条件下,Fv/Fm的变化极小,不受物种和生长条件的影响。在低温胁迫下,降香黄檀、闽楠和铁刀木的Fv/Fm显著低于对照(p<0.05),表现出持续下降的趋势,表明植物受到胁迫或由于某种原因猝灭了荧光,导致这一效率值明显下降(Maxwell和Johnson,2000);而降香黄檀、闽楠和铁刀木的Fv/Fm在低温胁迫处理间差异不显著(p>0.05)。格木、檀香和樟树经历2次低温胁迫后Fv/Fm表现为先降低后升高的趋势,但总体低于对照的Fv/Fm。6个树种的Fv/Fm与对照相比均偏低,表明低温胁迫使珍贵树种PSⅡ的光能转换效率降低。表观量子传递速率(ETR),即非循环电子传递速率,反映植物叶片总体光合能力(Baker,2008)。本试验发现,经历2次低温胁迫后降香黄檀、闽楠、铁刀木和樟树的ETR呈现出持续下降的趋势,而檀香和格木则表现为先升高后降低。方差分析结果表明,降香黄檀、檀香、铁刀木和樟树的ETR在低温胁迫处理间差异不显著(p>0.05),却显著低于对照(p<0.05),下降为对照的28.1%~58.9%,这表明各树种的光合能力降低(图4B)。
光化学淬灭(qP)反映的是PSⅡ天线色素吸收的光能用于光化学电子传递的份额,一定程度上反映了PSⅡ反应中心的开放程度(Maxwell和Johnson,2000)。从图5A可知,降香黄檀、闽楠、
檀香、铁刀木和樟树经历2次低温胁迫后qP呈现出持续下降的趋势,表明PSⅡ反应中心的开放程度降低。而格木qP表现为先升高后降低的趋势,这可能是格木受低温胁迫后通过增加qP来抵抗胁迫,但第2次持续胁迫qP降低,可见持续低温对格木PSⅡ反应中心的开放程度存在影响。与对照相比,降香黄檀、檀香和樟树的qP均显著下降(p<0.05),分别下降为对照的38.7%、39.4%和26.5%。而闽楠和铁刀木的qP在对照、低温胁迫Ⅰ和Ⅱ间差异不显著(p>0.05)。
非光化学淬灭(qN)是一种自我保护机制,对光合机构起一定的保护作用(周蕴薇等,2006)。与对照相比,6种珍贵树种经历2次低温胁迫后qN基本上呈下降的趋势,表现为对照下格木、降香黄檀、闽楠和铁刀木的qN显著高于低温胁迫(p<0.05),而2次低温胁迫处理间格木、降香黄檀、闽楠和铁刀木qN差异不显著(p>0.05)(图5-B)。此结果表明经历第1次胁迫时珍贵树种还未来得及启动防御机制,而随着胁迫时间的延长,光合器官的热耗散能力得到加强。。
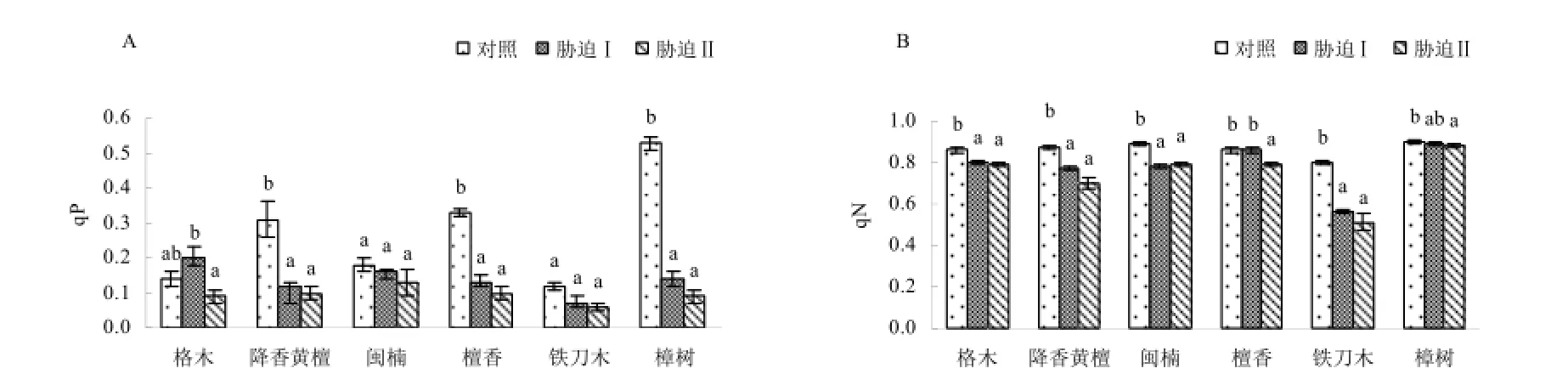
图5 低温胁迫对苗木光化学淬灭(qP)和非光化学淬灭(qN)的影响Fig.5 Effect of low temperature stress on qP and qN in seedling leaves of the examined plants

表2 各参数之间相关性分析Table 2 Correlation analysis between the indexes
2.4 各指标之间相关性分析
对光合参数和叶绿素荧光参数进行相关性分析(表2)。结果表明,Pn与Gs、Tr、Fv/Fm、qP、qN、ETR和Fm存在极显著相关关系(p<0.01),说明Gs、Tr、Fv/Fm、qP、qN、ETR和Fm均可作为衡量这几种植物光合能力的参数。本试验各树种受低温胁迫后Pn、Gs均降低而Ci升高,说明低温可能破坏了光合机构,导致CO2同化力下降,CO2大量积累,使Ci升高,Pn降低。Fv/Fm与qP、qN、ETR、Fm呈极显著正相关(p<0.01),Fv/Fm、qP、qN、ETR和Fm均有不同程度的下降。光合色素降解造成Fm下降(李磊等,2011),Fm下降可说明低温胁迫破坏了植物的光合机构。
2.5 利用隶属函数法评价苗木的抗寒性
隶属函数法已广泛应用于竹柳、花椒、珍珠菜等植物的抗寒性鉴定(许桂芳等,2009;吕小军等,2013;张捷等,2013)。本试验在测定多项生理指标的基础上,利用隶属函数法对植物的抗寒性进行综合评价。用每个树种各项指标隶属值的平均值作为树种抗寒能力综合鉴定标准,平均值越大,表明植物抗寒性越强(表3)。从平均值可知,6个树种抗寒能力依次为格木>樟树>闽楠=檀香和铁刀木>降香黄檀。
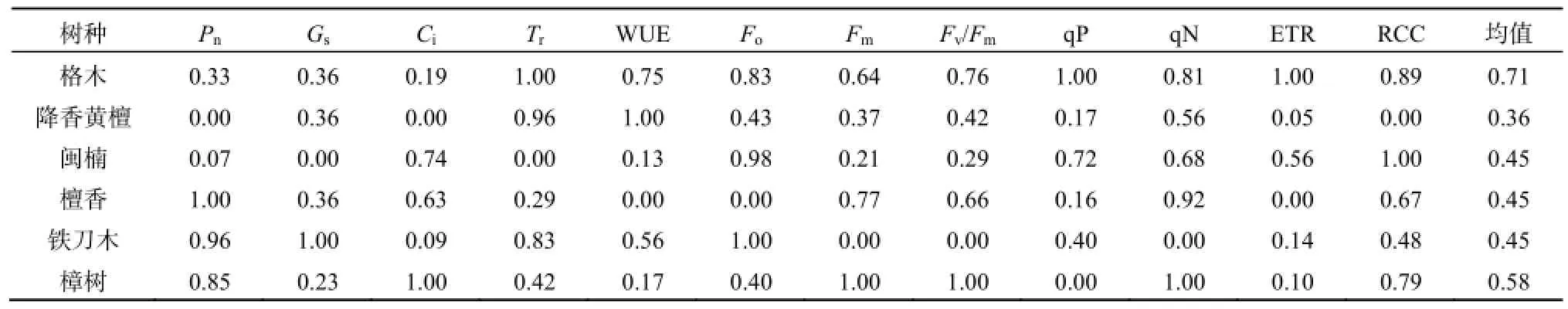
表3 6种苗木叶绿素荧光参数、光合参数和相对叶绿素含量的抗寒系数及隶属函数平均值Table 3 cold resistance coefficient of chlorophyll fluorescence parameters, photosynthetic parameters and relative chlorophyll content and their subordinative function mean value of 6 kinds of seedlings
3 讨论
3.1 幼苗抗寒性与光合参数
Pn下降主要由两个原因引起:气孔限制因素和非气孔限制因素。气孔限制因素引起的Pn下降表现为Gs和Ci均下降,非气孔限制因素引起的Pn下降表现为Gs下降而Ci升高(刘建等,2007)。本试验,6种珍贵树种Pn与对照相比均有不同程度的降低。在低温胁迫下6种树种Gs均低于对照,格木、降香黄檀、闽楠、檀香、铁刀木的Ci低温胁迫后升高,而樟树的Ci在低温胁迫后处于下降趋势,表明在低温胁迫下,格木、降香黄檀、闽楠、檀香、铁刀木净光合效率的下降很大程度上来源于非气孔因素抑制,而樟树净光合速率下降可能来源于气孔限制因素。本试验Tr的降低极有可能是由于气孔导度降低所致。植物能否适应当地的极限环境条件,关键在于它们能否很好地协调碳同化和水分耗散之间的关系,即植物水分利用效率(WUE)是其生存的关键因子(曹生奎等,2009)。格木和铁刀木的WUE变化不显著,降香黄檀WUE显著降低,而闽楠、檀香和樟树的WUE显著升高,这说明低温下闽楠、檀香和樟树能较好地协调碳同化和水分耗散之间的关系。
3.2 幼苗抗寒性与叶片相对叶绿素含量
低温胁迫期间格木和闽楠幼苗叶片的相对叶绿素含量上升后下降,其余幼苗叶片的叶绿素含量均下降。相对叶绿素含量下降,一方面可能是因为叶绿体色素合成酶活性降低,叶绿体合成受阻而造成;另一方面也可能是低温引起了叶绿体功能的紊乱,或加速了叶绿素的分解和叶绿体形态结构的受损使植物叶绿素含量减少(梁李宏等,2009)。此外,低温使植物体代谢缓慢,合成叶绿素的原料不足,也可能会造成叶绿素含量减少,相对叶绿素含量降低。
3.3 幼苗抗寒性与荧光参数
低温条件下,Fo的降低是PSⅡ的热耗散增加所致,Fo的增加表明PSⅡ反应中心已受到不可逆转的失活或破坏(王宁等,2013)。格木在低温胁迫前后,Fo变化不显著说明其受低温影响较小;其它树种都受到一定程度的低温影响。檀香和铁刀木Fo值在受胁迫前后差异显著,说明低温胁迫降低了其原初光能转化效率,引起了光抑制造成PSⅡ反应中心失活或伤害,这与杨猛等(杨猛等,2012)对玉米低温胁迫的研究结果一致。低温胁迫后,所有苗木的Fm均下降,说明6种珍贵树种光合机构的非光化学过程增大,电子传递潜力降低。Fv/Fm是近年来常用的研究植物对逆境响应的重要生理参数,在胁迫环境中Fv/Fm值下降(孔海云等,2011)。低温胁迫后所有植物Fv/Fm值均下降,其中格木、檀香和樟树的Fv/Fm值下降幅度较小,闽楠、降香黄檀和铁刀木的Fv/Fm值下降的幅度较大,可见闽楠、降香黄檀和铁刀木PSⅡ的最大光化学效率受低温胁迫的影响较大。低温胁迫对苗木的最大光能转化有较大的影响,主要是抑制了植物的光能转换效率。光化学淬灭系数qP愈大,则QA-重新氧化形成QA的量愈大,即PSⅡ的电子传递活性愈大;PSⅡ光化学淬灭系数qP越低,说明PSⅡ反应中心开放部分的比例越小,能够传递的电子数量也较少,单位时间内光合电子传递的速度较慢(周建等,2009)。除格木在低温胁迫后qP先升高后下降外,其它树种在受低温胁迫后qP持续下降,这说明除格木外5种珍贵树种苗木在低温胁迫下光合碳同化能力均受到影响。这与茶树(孔海云等,2011)、苜蓿(周建等,2009)、杨梅(刘辉等,2005)等植物方面的研究结果一致。本研究6种植物在低温胁迫后qN表现出显著下降趋势,说明严重的低温胁迫可以显著抑制PSII的热耗散。这与陈梅和唐运来(2012)对玉米低温胁迫研究的结果一致。除格木外,其它树种苗木的ETR低温胁迫后均呈下降趋势,说明低温损害了植物叶片的光合机构,导致光合电子传递效率下降。
广州属南亚热带季风气候,温度波动幅度较大,冬季常有极端低温的天气,特别是倒春寒,其特点是降温幅度大、降温速度快、降温持续时间长,这将对苗木的抗寒性产生直接的影响;本研究6种
珍贵树种苗木的光合作用对低温胁迫比较敏感,低温胁迫对珍贵树种叶片的光合机构造成一定的伤害。低温环境中,这几种植物叶片对光能的吸收传递光化学转换以及光合电子传递都受到低温的显著抑制,这必将影响到植物的生长。若要向冬季气温更低的地区引种这几种植物,应当优先考虑其抗寒性。
4 结论
(1)冬季低温胁迫下6种珍贵植物苗木净光合速率降低,格木、降香黄檀、闽楠、檀香、铁刀木净光合效率的下降很大程度上来源于非气孔因素抑制,而樟树净光合速率下降可能来源于气孔限制因素。
(2)冬季低温胁迫下6种珍贵植物苗木叶片相对叶绿素含量降低。
(3)冬季低温胁迫下6种珍贵植物苗木Fm、Fv/Fm、ETR、qP和qN等叶绿素荧光参数下降,植物叶片的光合机构受到一定的伤害。
(4)相关性分析表明Gs、Tr、Fv/Fm、qP、qN、ETR和Fm均可作为衡量这几种植物光合能力的参数,但无法从单一指标判断植物抗寒能力的强弱。利用隶属函数计算各个指标的隶属函数值,由隶属函数平均值可知抗寒能力强弱表现为格木>樟树>闽楠=檀香=铁刀木>降香黄檀。
BAKER N R. 2008. Chlorophyll fluorescence: a probe of photosynthesis in vivo[J]. Annual Review of Plant Biology, 59: 89-113.
GUILBAULT K R,BROWN C S,FRIEDMAN J M,et al. 2012. The influence of chilling requirement on the southern distribution limit of exotic Russian olive (Elaeagnus angustifolia) in western North America. Biological Invasions, 14(8): 1711-1724.
MAXWELL K, JOHNSON G N. 2000. Chlorophyll fluorescence-a practical guide[J]. Journal of Experimental Botany, 51(345): 659-668.
MIKKELSEN T N. 1996. Acceleration of leaf senescence in Fagus sylvatica L. by low levels of tropospheric ozone demonstrated by leaf colour, chlorophyll fluorescence and chloroplast ultrastructure[J]. Trees, 10: 145-156.
曹生奎, 冯起, 司建华, 等. 2009. 植物叶片水分利用效率研究综述[J].生态学报, 29(07): 3882-3892.
陈梅, 唐运来. 2012. 低温胁迫对玉米幼苗叶片叶绿素荧光参数的影响[J]. 内蒙古农业大学学报: 自然科学版, 33(3): 20-24.
邓化冰, 车芳璐, 肖应辉, 等. 2011. 开花期低温胁迫对水稻花粉性状及剑叶理化特性的影响[J]. 应用生态学报, 22(1): 66-72.
关雅楠, 黄正来, 张文静, 等. 2013. 低温胁迫对不同基因型小麦品种光合性能的影响[J]. 应用生态学报, 24(7): 1895-1899.
郭文福, 贾宏炎. 2006. 降香黄檀在广西南亚热带地区的引种[J]. 福建林业科技, 33(4): 152-155.
郭延平, 张良诚, 洪双松, 等. 2000. 温州蜜柑叶片气体交换和叶绿素荧光对低温的响应[J]. 植物生理学报, 26(2): 88-94.
何跃君, 薛立, 任向荣, 等. 2008. 低温胁迫对六种苗木生理特性的影响[J]. 生态学杂志, 27(4): 524-531.
贺磊, 游凯, 李远芳, 等. 2011. 5种典型热带引种园林植物幼苗抗寒性研究[J]. 中南林业科技大学学报, 31(2): 65-71.
姜卫兵, 李刚, 翁忙玲, 等. 2007. 五种木兰科树种在南京地区冬春季节的光合特征[J]. 应用生态学报, 18(4): 749-755.
孔海云, 张丽霞, 王日为. 2011. 低温与光照对茶树叶片叶绿素荧光参数的影响[J]. 茶叶, 37(2): 75-78.
李磊, 李向义, 林丽莎, 等. 2011. 两种生境条件下6种牧草叶绿素含量及荧光参数的比较[J]. 植物生态学报, 35(6): 672-680.
梁李宏,梅新,林锋,等.2009. 低温胁迫对腰果幼苗叶片组织结构和生理指标的影响[J]. 生态环境学报,18(1): 317-320
林宁, 刘昆成, 於艳萍, 等. 2012. 低温胁迫对格木幼苗生理特性的影响[J]. 北方园艺, (24): 87-90.
刘辉, 郭延平, 胡美君. 2005. 杨梅光合作用的低温光抑制[J]. 热带亚热带植物学报, 13(4): 338-342.
刘建, 项东云, 陈健波,等. 2009. 低温胁迫对桉树光合和叶绿素荧光参数的影响[J].桉树科技, 26(1).
刘建, 叶露, 周坚, 等. 2007. 夜间低温对2种桉树幼苗光合特性的影响[J]. 西北植物学报, 27(10): 2024-2028.
吕小军, 杨途熙, 何小红, 等. 2013. 冬季低温对花椒抗寒性生理指标的影响[J]. 西北农业学报, 22(7): 143-148.
亓白岩, 殷云龙, 於朝广, 等. 2013. 木兰科含笑属8种植物叶片解剖结构性状与抗寒性的关系[J]. 江苏农业科学, 41(4): 150-153.
邵怡若, 许建新, 薛立, 等. 2013. 低温胁迫时间对4种幼苗生理生化及光合特性的影响[J]. 生态学报, 33(14): 4237-4247.
王宁, 姚方, 袁美丽, 等. 2013. 自然降温对猴樟和芳樟叶绿素荧光参数的影响[J]. 河南农业大学学报, 47(4): 414-419.
王宁, 袁美丽, 苏金乐. 2013. 几种樟树叶片结构比较分析及其与抗寒性评价的研究[J]. 西北林学院学报, 28(4): 43-49.
王宇涛, 李春妹, 李韶山. 2013. 华南地区3种具有不同入侵性的近缘植物对低温胁迫的敏感性[J]. 生态学报, 33(18): 5509-5515.
许桂芳, 张朝阳, 向佐湘. 2009. 利用隶属函数法对4种珍珠菜属植物的抗寒性综合评价[J]. 西北林学院学报, 24(3): 24-26.
杨猛, 魏玲, 庄文锋, 等. 2012. 低温胁迫对玉米幼苗电导率和叶绿素荧光参数的影响[J]. 玉米科学, 20(1): 90-94.
张捷, 杨淑梅, 王玲. 2013. 低温胁迫下2个‘竹柳’品系的抗寒性[J]. 林业科学, 49(9): 158-164.
赵一鹤, 李建宾, 杨时宇, 等. 2012. 低温胁迫下3个甜角品种幼苗的抗寒性研究[J]. 林业科学研究, 25(2): 150-156.
周建, 杨立峰, 郝峰鸽,等. 2009. 低温胁迫对广玉兰幼苗光合及叶绿素荧光特性的影响[J]. 西北植物学报, 29(1): 0136-0142.
周蕴薇, 刘艳萍, 戴思兰. 2006. 用叶绿素荧光分析技术鉴定植物抗寒性的剖析[J]. 植物生理学通讯, 42(5): 945-950.
朱先灿, 宋凤斌, 徐洪文. 2010. 低温胁迫下丛枝菌根真菌对玉米光合特性的影响[J]. 应用生态学报, 21(2): 470-475.
Influence of low temperature stress on the photosynthetic fluorescence characteristics of 6 kinds of precious hardwoods seedling in winter
ZHANG Yilong1, ZHANG Weiqiang2*, GAN Xianhua2
1. College of Forestry, South China Agricultural University, Guangzhou 510642, China; 2. Guangdong Academy of Forestry, Guangzhou 510520, China
The gas exchange parameters, chlorophyll fluorescence parameters and the relative chlorophyll content of 6 kinds of precious hardwoods seedlings of Erythrophleum fordii, Dalbergia odorifera,Phoebe bournei, Santalum album, Cassia siamea, and Cinnamomum camphorawere were measured in winter using the LI-6400 portable photosynthesis system, and the obtained data were analyzed to explore the relationship between these parameters and cold resistance. The method of subordination function was used to evaluate cold resistance comprehensively. The net photosynthetic rate (Pn), stomatal conductance (Gs) and transpiration rate (Tr) of the six tree species seedlings declined in the process of low temperature stress comparing with controls. The decline of Pnof D. odorifera, S. album and C. siamea mainly caused by non-stomatal limitation, while of C. camphora caused by stomatal limitation. The water use efficiency (WUE) of P. bournei, E. fordii and C. siamea increased in low temperature stress, while WUE of E. fordii and D. odorifera decreased,this showed that P. bournei, E. fordii and C. siamea have better ability in coordinating the relationship between carbon assimilation and water dissipation. The maximum fluorescence (Fm) and Maximum quantum efficiency of PSII photochemistry(Fm/Fv)of each species decreased to some extent under low temperature stress which indicated that low temperature produced significant inhibition on the electron transfer potential of PSII reaction centers and reduced photochemical efficiency of PSII of the six tree species seedlings. The qP of D. odorifera, P. bournei, S. album and C. siamea showed a declining trend after 2 times low temperature stress which indicated that the degree of openness of PSII reaction centers of precious hardwood decreased. The qN of E. fordii, D. odorifera, P. bournei and C. siamea showed no significant difference (p>0.05) after 2 times low temperature stress which indicated that those species had excess energy dissipation mechanisms and protected the photosynthetic apparatus from damage. Correlation analysis showed that there were significantly positive correlations between Pnand Gs, Tr, Fv/Fm, qP, qN, ETR, Fm(p<0.01). Therefore, those parameters could be used as parameters to measure the photosynthetic capacity. The mean value of the subordinate function indicated that their ability of low temperature resistance from high to low E. fordii, C. camphora>P. bournei=S. album=C. siamea>D. odorifera. In short, low temperature stress destroy the 6 kinds of seedling photosynthetic organization which may be the reason that induce plant photosynthetic capacity reduction, and the degree of response of different plants to chilling stress are different, which should be taken into account when consider to expand the introduction and cultivation of those trees.
low temperature stress; gas exchange parameters; chlorophyll fluorescence parameters; cold tolerance evaluation; cold tolerance screening
Q945.78
A
1674-5906(2014)05-0777-08
广州市科技计划项目(12B541040007);国家林业公益性行业科研专项(201404301);广东省林业科技创新专项(2010KJCX013-02);佛山市生态景观林培育技术研究与推广示范和国家林业局广东东江源森林生态站联合资助
张毅龙(1988年生),男,硕士,研究方向为森林生态。E-mail: 908897597@qq.com *通信作者:张卫强(1976年生),男,高级工程师,博士,主要从事森林水文与植物生理生态。E-mail: happyzwq@sina.com
2014-03-03
张毅龙,张卫强,甘先华. 低温胁迫对6种珍贵树种苗木光合荧光特性的影响[J]. 生态环境学报, 2014, 23(5): 777-784.
ZHANG Yilong, ZHANG Weiqiang, GAN Xianhua. Influence of low temperature stress on the photosynthetic fluorescence characteristics of 6 kinds of precious hardwoods seedling in winter [J]. Ecology and Environmental Sciences, 2014, 23(5): 777-784.
植物光合作用与其所处生态环境密切相关,温度是自然界中限制植物光合作用以及地理分布最重要的生态因素之一(Guilbault等,2012;姜卫兵等,2007)。低温会影响植物生长发育、生理和光合特性,而热带或亚热带地区全年气候较为温暖(何跃君等,2008),因此分布于该区域的植物种类大多缺乏对低温环境的耐受性(王宇涛等,2013)。目前,针对低温胁迫对植物生理影响的研究很多,邵怡若等(2013)通过人工模拟低温环境,从叶绿素、渗透调节物质、抗氧化酶系统和光合特性等方面比较研究了盐肤木(Rhus chinensis)、假连翘(Duranta repens)、老鸭嘴(Thunbergia grandiflora)、葛藤(Pueraria lobota)的耐寒性强弱,结果表明,这4种植物的耐寒能力为葛藤>盐肤木>老鸭嘴>假连翘。周建等(2009)通过设置不同低温梯度研究低温胁迫对广玉兰(Magnolia grandiflora)幼苗叶片光合及叶绿素荧光特性的影响,结果表明低温胁迫除导致气孔受抑制外,还直接损伤光合机构使PSII反应中心失活,引起其光能原初捕捉能力和光能同化率减弱,提高了通过热辐射消耗的光能比例,最终导致广玉兰幼苗光合作用能力减弱。刘建等(2009)以人工低温处理尾叶桉(Eucalyptus urophylla)和邓恩桉(Eucalyptus dunnii)幼苗,研究低温对两种桉树光合和叶绿素荧光参数的影响,结果表明低温胁迫对尾叶桉的影响明显要大于对邓恩桉的影响。生理生态的研究方法已被广泛应用
于植物低温胁迫研究(王宇涛等,2013;关雅楠等,2013;邓化冰等,2011;朱先灿等,2010;Baker,2008),而利用叶绿素荧光参数和气体交换参数结合的研究方法比较少见(关雅楠等,2013;郭延平等,2000)。叶绿素荧光分析技术可以快速、灵敏和无损伤地研究和探测完整植株在胁迫下光合作用情况,评价植物光合机构的功能和环境胁迫对植物的影响(Baker,2008)。而利用叶绿素荧光参数和气体交换参数结合的研究方法能够获得植物对环境反应的全貌(Maxwell和Johnson, 2000)。
猜你喜欢
林业与生态(2022年5期)2022-05-23
中南林业科技大学学报(2020年5期)2020-06-06
天然产物研究与开发(2018年5期)2018-06-13
山东林业科技(2017年1期)2017-06-29
广东药科大学学报(2016年1期)2016-04-11
广西林业科学(2016年2期)2016-03-20
广西林业科学(2016年1期)2016-03-20
中国继续医学教育(2015年4期)2016-01-07
中国民族医药杂志(2015年7期)2015-07-13
意林(2011年21期)2011-05-14
