
植物对硒的吸收、转化及生理功能研究综述
2014-02-27 09:00万海英向极钎杨永康殷红清
安徽农业科学 2014年21期
万海英,向极钎,杨永康 ,殷红清,马 进
(1.湖北省农业科技创新中心鄂西综合试验站,湖北恩施445000;2.恩施清江生物工程有限公司,湖北恩施445000)
硒是动物和人体必需的微量元素,缺乏或过量都引起各种疾病。研究表明,地方性疾病如克山病、大骨节病、克汀病、地方性癌症等都是由缺硒造成的[1-2]。由于缺硒会使谷胱甘肽过氧化物酶(GPX)的活性降低,组织细胞的抗氧化损伤能力减弱,影响细胞的分裂、繁殖等,从而干扰核酸、蛋白质、糖和酶的合成及其代谢[3]。
通过植物对硒的吸收进行补硒是最经济、最安全的途径。植物作为自然界中硒循环的重要载体,可以将土壤中的无机态硒经过吸收、同化转变为有机硒,进而被人和动物所摄取以达到补硒的目的。硒虽然对人和动物的正常生理活动有益,但不是越多越好,满足的范围较狭窄,过量的硒会造成人和动物的硒中毒。
1 植物对硒的吸收
植物对硒元素的吸收是其进入食物链的有效途径。低浓度的硒有助于植物生长,而高浓度的硒则对植物生长有抑制作用[4]。研究表明,对胡萝卜叶面喷施硒盐,其硒含量可增加到2 μg/g(干基),与对照间差异达 0.05 显著水平[5]。在贯叶连翘上进行叶面施硒试验,发现植物器官中硒含量达到1 000 ~12 000 μg/g,比对照提高了 100 倍[6]。利用水稻的生物富集和转化作用,在水稻上施用硒肥,把非生物活性和毒性高的无机硒转化为毒性低、安全有效的活性有机硒。这是改善和满足食物链中硒水平不足的廉价且可行的方法之一[7]。富硒玉米籽粒中的硒以有机结合态硒为主,占总硒的比例接近90%,是其主要的硒存在形式;在有机态硒中蛋白质硒含量较高,多糖中硒的含量较低,在蛋白质硒中以碱溶蛋白质中硒的含量为最高,占总硒的50.62%[8]。根据不同植物对硒的吸收和富集能力的不同,将植物划分为聚硒植物和非聚硒植物。聚硒植物如黄芪(Ast ragalus)硒含量高达10 g/kg,可以作为硒的指示植物,而大部分农作物是非聚硒植物,聚硒能力低,一般含量在0.001~1.000 μg/g。植物对硒的富集能力依次为十字花科>黑麦草>豆类>谷类,对蔬菜作物富硒能力最强的是大蒜。
土壤中的硒按价态可分为元素态硒、硒化物、亚硒酸盐、硒酸盐、有机态硒化物和挥发态硒,其中有机态硒化物是土壤有效硒的主要来源,挥发态硒在土壤微生物的作用下转化成气态烷基化合物。植物对硒的吸收可以通过根部和叶片,吸收的主要形态是Se4+和Se6+[9-10]。根吸收的Se4+是被动吸收,在植株体内积累的浓度低于根外土壤环境的硒浓度,而Se6+能主动地在植物体内累积,使其浓度高于外部环境硒浓度[11]。由于Se4+和Se6+在土壤中迁移转化方式的不同,其在土壤中移动性、生物有效性和毒性大小存在差异。有研究表明,在水培条件下,植物对硒酸盐和亚硒酸盐的吸收速率的研究结果一致[12];但是,在土培条件下,硒酸盐更易被吸收和转运,因而生物有效性更高[13-14],而亚硒酸盐更易被土壤表面所吸附[15],也更易累积在植物的根部[16],所以在生产上常选用硒酸盐作为肥料[17],但是在田间喷施上由于六价硒必须首先在叶中转化为二价硒再转化为有机硒,而四价硒不需要酶催化就能直接转化为有机硒而进入籽粒,故多选用亚硒酸盐作为喷施硒肥[18]。但是,也有研究表明,植物对Se4+的吸收过程并不是完全的被动吸收。试验中,加呼吸抑制剂时20%的Se4+的吸收受到抑制,因此至少有部分的吸收是与根部代谢相关联的[19]。另外,非聚硒植物的累积的硒含量与植物生长年限、时间长短有很大关系[20]。
2 硒在土壤-植物系统中的转化
在土壤的环境条件下,Se0、Se4+、Se6+在微生物氧化还原的条件下相互转换,其中Se4+在高亲和磷酸转运蛋白,Se6+在高亲和硫酸酯转运蛋白的作用下由土壤中进入植物的根系中,Se4+在根部部分转化为有机硒,然后转换为二甲基硒化物(DMSe)以气态的形式排出根外。在植物的根部Se6+可以转化为Se4+,Se4+可以转化为Se0,但是具体的转运途径目前尚不清楚。其中,进入根部的Se6+经过木质部可以进入植物的叶绿体中,至于转运机制目前仍尚未清楚。Se6+被吸收进入植物叶绿体细胞后在5-磷硫酸腺苷(APS)的作用下转化为Se4+,然后经过硒代半胱氨酸甲基转移酶的作用下转化成硒代半胱氨酸(SeCys),部分SeCys在一种半胱氨酸脱硫酶(CpNifs)和硒代半胱氨酸裂合酶(SL)的作用下转化成Se0,部分SeCys在胱硫醚-γ-合成酶作用下转化成硒代胱硫醚,进而转化成硒代高半胱氨酸(Sehocys),此时硒代半胱氨酸在未知的转运机理的条件下进入细胞质中。部分SeCys可直接进入细胞质中,但进入机理也尚未清楚。进入细胞质的Sehocys进一步转化成硒代蛋氨酸(SeMet),SeMet在蛋氨酸甲基转移酶(MMT)的作用下转化成硒甲基蛋氨酸(SeMM),进而转化成二甲基硒化物(DMSe),以气态的形式排出植物体外。部分直接进入细胞质中的SeCys有2种转运途径,一种是在硒代半胱氨酸裂合酶(SL)的作用下转化成Se0,另一种是在硒代半胱氨酸甲基转移酶(SMT)的作用转化成甲基硒代半胱氨酸(MeSecys),进而转化成二甲基二硒化物(DMDSe),以气态的形式排出植物体外。其中,部分有机硒可以保存在种子中。目前,植物对硒的挥发性的由于其可以作为一种有效的植物修复土壤硒污染的方法而得到广泛的关注[21](图 1)。
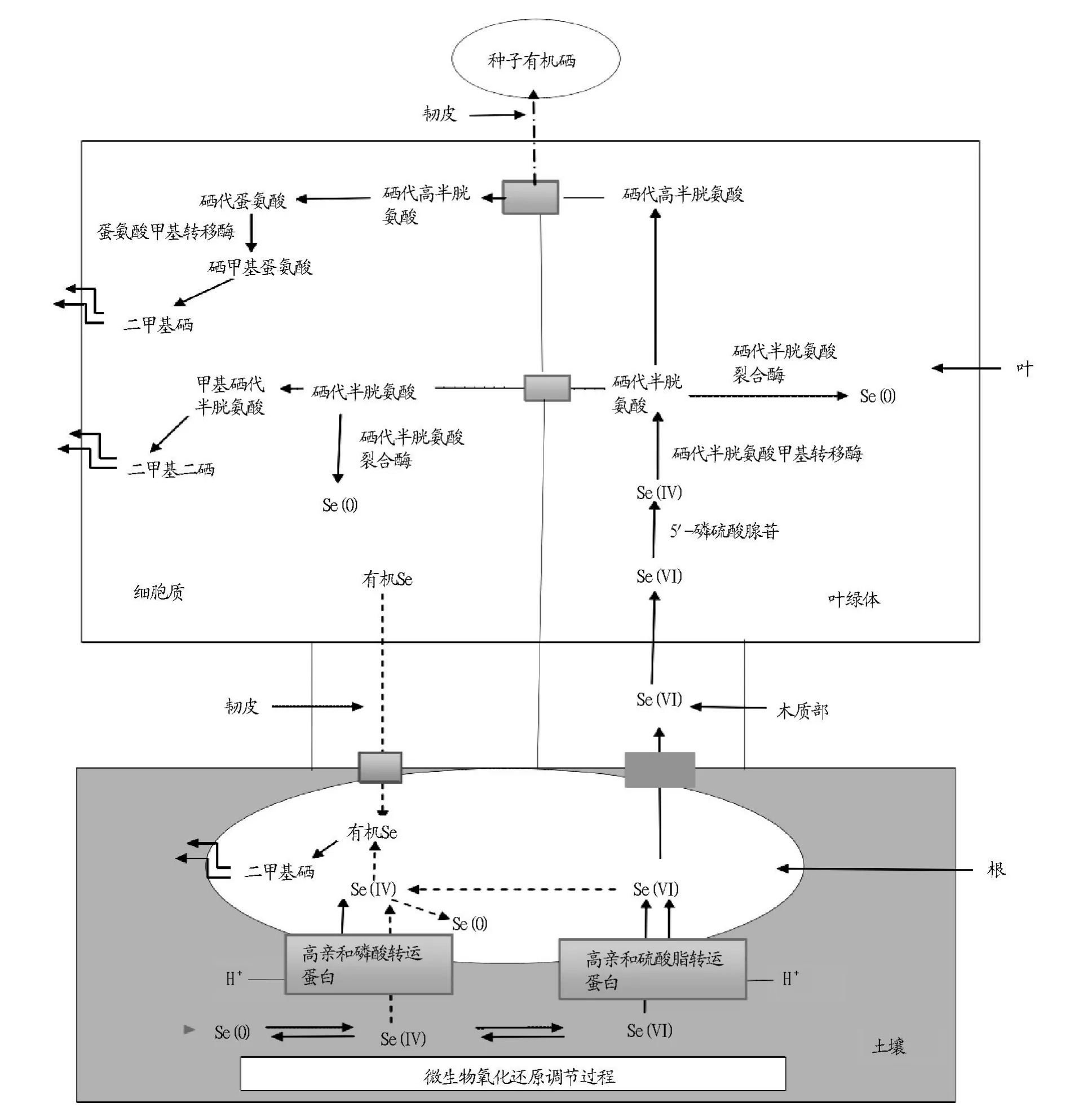
图1 硒在土壤-植物中的新陈代谢图
3 硒对植物生理功能的影响
硒对植物生理功能有重要影响。在植物中,除了发现硒代半胱氨酸和硒代蛋氨酸外,还有多种低分子量有机硒化物(表1)存在于机体,多数是从植物中得到非蛋白硒代氨基酸衍生物[22]。
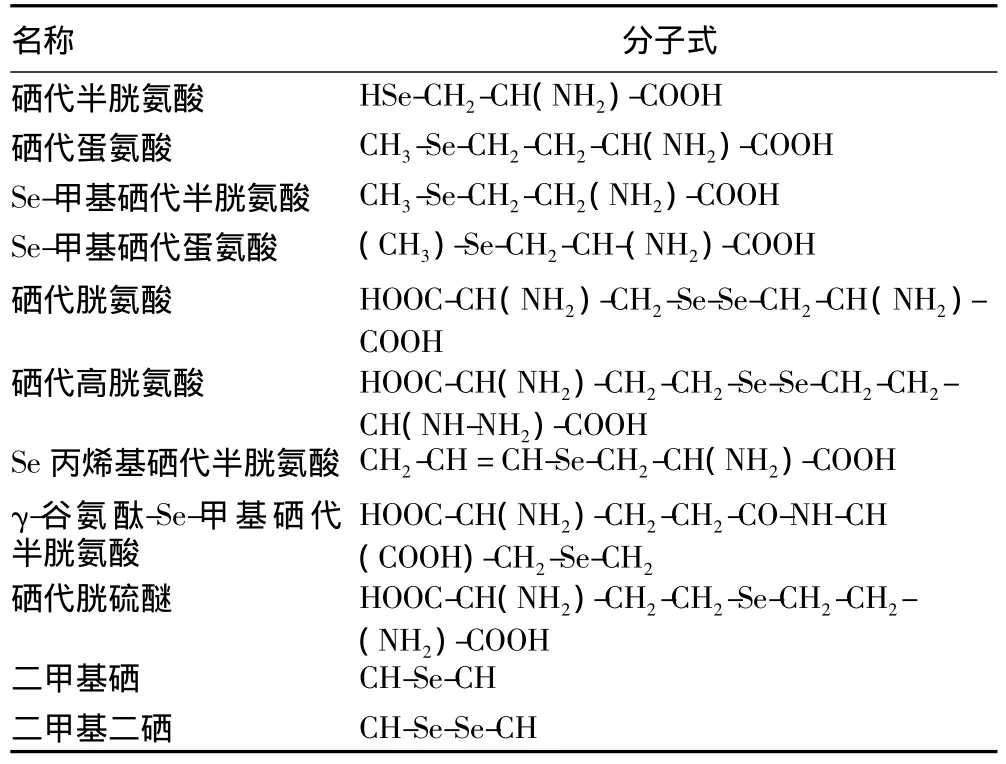
表1 植物中发现的低分子量有机硒化合物
硒元素不仅对植物生理功能的实现有重要的意义,而且对种子萌发、根系活力、叶绿素含量等有显著影响。不同类型的植物种子萌发所需适宜的硒浓度也有所差异。适宜浓度的硒处理种子能够促进其萌发。浓度高的硒处理反而具有毒害作用,不利于种子的萌发。研究表明,用0.05 mg/L的硒浓度处理大豆种子的萌发率最高,其发芽率、发芽势、发芽指数和活力指数也相应的提高,但浓度过高时对大豆种子发芽有明显的抑制作用[23]。不同价态的硒在低浓度时均能促进油菜、白菜、萝卜种子的萌发,提高发芽势和发芽率。硒对种子萌发的影响与价态无关。硒促进种子萌发可能与影响种子的酶活性和物质代谢有关[24]。硒对根系活力的影响同样与硒浓度有关。研究发现,当硒浓度低于21.0 μg/ml时,油菜根系活力随硒浓度上升而增加,但浓度过高时引起根系活力降低[25]。在烤烟叶片中,当硒浓度为4 μg/g时,叶绿素a、叶绿素b、叶绿素a+b、类胡萝卜素含量分别比对照增加5.37%、17.53%、6.56%、18.75%,当硒浓度为8 μg/g时其叶绿素含量开始下降,当硒浓度为16 μg/g时烤烟叶片中叶绿素和类胡萝卜素含量与对照相比显著下降(P<0.05)[26]。另外,硒还能够对植物体内多种酶的活性具有一定的影响。低浓度Se可以提高烟株体内活性氧防御体系,使得MDA酶含量下降;高浓度Se则对其有抑制作用,对活性氧防御体系产生胁迫效应,MDA含量上升,SOD、POD酶活性下降[26]。硒是非金属元素,能增强植物的抗氧化性和对重金属的耐受性。有研究发现,施用硒酸钠溶液能提高地上部分含砷量7%~45%,也能提高蜈蚣草中的硫醇含量和谷胱甘肽含量,抑制脂类的过氧化反应。这是因为硒激活了植物的保护机制,减轻了其氧化压力,提高了蜈蚣草对砷的吸收[27]。
水培试验结果表明,硒能提高水稻幼苗叶片超氧化物歧化酶、过氧化物酶活性,降低过氧化氢酶活性[28]。硒能促进小麦幼苗体内谷胱甘肽过氧化物酶合成,提高幼苗体内谷胱甘肽过氧化物酶活性。随着硒处理时间的增加,硒对谷胱甘肽过氧化物酶合成的促进作用逐渐增强[29]。硒能影响植物体内谷胱甘肽过氧化物酶、过氧化物酶、多酚氧化酶、超氧化物歧化酶、过氧化氢酶等多种酶活性。
4 结语
硒是人和动物生命中必需的微量元素,同时对植物的生长发育有显著的影响。一定浓度的硒对植物生长发育有着积极的促进作用。在增加产量、提高品质方面,硒有着显著的功效。在植物体内,硒的适用范围很狭窄。一般,低硒或适量硒对植物具有促进作用,而高浓度和过量硒反而造成毒害。目前,硒对植物的毒害机制机理是过多的硒代半胱氨酸替代半胱氨酸参与蛋白质的合成,但是否具有其他毒害机制仍需进一步探讨。目前,市场上的绝大多数富硒农产品是通过外源施硒肥获得的,但是长期施用硒肥不仅成本较高,而且易造成环境污染,因此筛选出具有一定富硒能力的农产品并通过一定的育种措施选育出富硒植物新品种是今后硒研究的方向之一。
[1]中国科学院地理研究所环境与地方病组.我国低硒带与克山病、大骨节病病因关系的研究[J].环境科学,1986,7(4):89.
[2]谭见安译.环境硒与健康[M].北京:人民卫生出版社,1989:219-224.
[3]WORTZMAN M S.Effect of dietary selenium on the in-teraction between 2-acetylaminofluorene and rat liver DNA in vi-vo[J].Cancer Res,1980,40:2670.
[4]HARTIKAINEN H.Biogeochemistry of selenium and its impact on food chain quality and human health(Review Article)[J].Journal of Trace Elements in Medicine and Biology,2005,18(4):309-318.
[5]KAPOLNA E,HILLESTRØM P R,LAURSEN K H,et al.Effect of foliar application of selenium on its uptake and speciation in carrot[J].Original Research Article Food Chemistry,2009,115(4):1357-1363.
[6] MATEJA G,VEKOSLAVA S,SAMO K.Selenium concentration in St.John’s wort(Hypericum perforatum L.)herb after foliar spraying of young plants under different UV-B radiation levels[J].Food Chemistry,2009,117(9):204-206.
[7]周鑫宾,施卫明,杨林章.叶面喷硒对水稻籽粒硒富集及分布的影响[J].土壤学报,2007,44(1):73-77.
[8]向东山,翟琨,刘晓鹏.富硒玉米子粒中硒赋存形态研究[J].玉米科学,2007,15(5):93-95.
[9]NOWAK J,KAKLEWSKI K,LIGOCKI M.Influence of selenium on oxidoreductive enzymes activity in soil and plants[J].Soil Biology and Biochemistry,2004,36(10):1553-1558.
[10]BANUELOS G S,LIN Z Q.Phytoremediation management of seleniumladen drainage sediments in Santhe Luis Drain:a greenhouse feasibility study[J].Ecotoxicology and Environmental Safety,2005,62(3):309-316.
[11]EUSTIEE D C,FOSTER I,KULL F J,et al.In vitro incorporation of selenome thionine into protion by vigna radiate[J].Polysomers Plant Physiol,1980,66:182-186.
[12]杨兰芳,丁瑞兴.恩施烟地土壤和烤烟团棵期硒含量与分布的研究[J].湖北民族学院学报,1998,66(1):27-29.
[13]LI H F,LOMBI E,STROUD J L,et al.Selenium speciation in soil and rice:influence of water management and se fertilization[J].J Agric Food Chem,2010,58:11837-11843.
[14]CARTES P,GIANFREDA L,MORA M L.Uptake of selenium and its antioxidant activity in ryegrass when applied as selenate and selenite forms[J].Plant and Soil,2005,276(1/2):359-367.
[15]吴雄平,梁东丽,鲍俊丹,等.Se(Ⅳ)和Se(Ⅳ)对小白菜生长及生理效应的影响[J].环境科学学报,2009,29(10):2163-2171.
[16]SUSANNE EICH-GREATOREX,TRINE A,SOGN A.Plant availability of inorganic and organic selenium fertilizer as influenced by soil organic matter content and pH[J].Nutr Cycl Agroecosyst,2007,79:221-231.
[17]王松山,吴雄平,梁东丽,等.不同价态外源硒在石灰性土壤中的形态转化及其生物有效性[J].环境科学学报,2010,30(12):2499-2505.
[18]BROADLY M R,WHITE P J,BRYSON R J.Biofortification of UK food crops with selenium[J].Proceedings of the Nutrition Society,2006,65(2):169-181.
[19]ARVY M P.Selenate and selenite uptake and translocation in bean plant(Phascolus vulgaris)[J].J Exp Bot,1993,44(263):1083-1087.
[20]KESKINEN R,MARJA TURAKAINEN M,HARTIKAINEN H.Plant availability of soil selenate additions and selenium distribution within wheat and ryegrass[J].Plant and Soil,2010,33(1/2):301-313.
[21]ZHU Y G,PILON-SMITS E A H,ZHAO F J,et al Selenium in higher plants:understanding mechanisms for biofortification and phytoremediation [J].Trends in Plant Science,2009,14(8):436-442.
[22]陈铭,谭见安,王五一.环境硒与健康关系研究中的土壤化学与植物营养学[J].土壤学进展,1994,22(4):1-10.
[23]李会芳,白云生,樊文华,等.不同浓度的硒对大豆种子发芽率及幼苗生长的影响[J].山西农业大学学报:自然科学版,2006,26(3):256-258.
[24]孙汉文,李朝昉,梁淑轩,等.硒对铬胁迫下蔬菜幼苗生长的影响[J].农业环境科学学报,2006,25(1):63-68.
[25]张驰,周大寨,吴永尧,等.硒对油菜苗期生长和生理生化指标的影响[J].湖北农业科学,2007,46(3):363-365.
[26]高家合,张晓海.硒对烤烟的生态毒理效应及临界指标研究[J].云南环境科学,2006,25(4):12-14.
[27]FENG R W,WEI C Y,TU S X,et al.Effects of Se on the uptake of essential elements in Pteris vittata L[J].Plant Soil,2009,325(1/2):123-132.
[28]吴秀峰,陈平.硒对水稻幼苗生长和生理特性的影响[J].农业与技术,2004,24(5):76-79.
[29]董卫华,李贞霞,夏金富.亚硒酸钠对小麦幼苗硒积累和谷胱甘肽过氧化物酶活性的影响[J].食品与生物技术学报,2006,35(2):1-5.
猜你喜欢
国际放射医学核医学杂志(2020年2期)2020-05-30
光谱学与光谱分析(2020年2期)2020-02-25
山东青年(2018年2期)2018-06-23
天然产物研究与开发(2016年6期)2016-06-05
北方文学·下旬(2016年6期)2016-05-14
分析测试学报(2015年6期)2016-01-13
东北电力大学学报(2015年4期)2015-11-13
集美大学学报(自然科学版)(2015年4期)2015-02-28
应用化工(2014年8期)2014-08-08
山东工业技术(2014年12期)2014-05-03

- 安徽农业科学的其它文章
- 水肥处理对苹果幼树树体结构、叶片及光合作用的影响