
长双歧杆菌α-半乳糖苷酶的分离纯化及酶学性质
2014-02-13 01:26陈俊亮霍贵成张慧芸
食品科学 2014年7期
陈俊亮,田 芬,霍贵成,张慧芸
(1.河南科技大学食品与生物工程学院,河南 洛阳 471003;2.东北农业大学 乳品科学教育部重点实验室,黑龙江 哈尔滨 150030;3.杭州娃哈哈集团有限公司研究院,浙江 杭州 310018)
α-半乳糖苷酶(α-galactosidase,α-Gal,EC3.2.1.22)属外切糖苷酶类,能够专一性催化非还原末端α-半乳糖苷键的水解[1],它不仅能够水解含α-半乳糖苷的半乳甘露低聚糖和棉子糖族低聚糖[2-3],而且能够水解含该键的多糖[3-5],此外,它还能作用于含有α-半乳糖苷的糖蛋白和糖脂质[6],因此α-半乳糖苷酶在食品、医药、饲料工业等领域得到广泛的应用[7-9]。在食品行业中,大豆蛋白质是氨基酸平衡模式的典范,但是在豆制品中存在α-半乳糖苷寡糖类物质,由于人的消化系统中缺乏降解此类物质的α-半乳糖苷酶,当这些物质消化经过大肠时,微生物发酵会产生大量气体,从而导致腹胀,因此在豆制品中添加α-半乳糖苷酶可以分解这些寡糖,增加人体对豆类营养成分的吸收[10]。
α-半乳糖苷酶主要来源于微生物、植物和动物,其中微生物的产酶量较高,国内外学者已筛选出多种产α-半乳糖苷酶活性的微生物,主要包括短双歧杆菌、青霉菌、酵母菌、放线菌等[10]。为了能够满足食品加工的要求,开发耐热性α-半乳糖苷酶,寻找高产酶量的微生物成为研究热点。实验以1株产α-半乳糖苷酶长双歧杆菌KLDS2.0509作为研究对象[11],从酶的pH值稳定性、热稳定性、不同金属离子的影响,以及底物特异性等方面进行研究,为今后开发α-半乳糖苷酶的乳酸菌发酵剂的研究提供依据。
1 材料与方法
1.1 材料与试剂
长双歧杆菌KLDS2.0509 乳品科学教育部重点实验室工业微生物菌种保藏中心。
Trypticase peptone-yeast extract(TPY)培养基:酪蛋白胨12 g/L、大豆蛋白胨5 g/L、多聚蛋白胨(polypepton)3 g/L、葡萄糖5 g/L、酵母粉5 g/L、低聚果糖5 g/L、吐温-80 1 g/L、L-半胱氨酸盐酸盐0.5 g/L、K2HPO43H2O 0.2 g/mL、柠檬酸氢二氨0.2g/mL、乙酸钠0.5 g/mL、硫酸镁58 mg/mL、硫酸锰25 mg/mL,pH6.5;蜜二糖、棉子糖、水苏糖、pNPG(p-nitrophenyl-α-D-galactopyranoside)、牛血清白蛋白 美国Sigma公司;酵母粉、蛋白胨 英国Oxiod公司;其余试剂为国产分析纯。
1.2 仪器与设备
BCN1360型生物洁净工作台 上海佳胜实验设备有限公司;DU800核酸/蛋白质分析仪 美国Beckman公司;SPX-150B生化培养箱 上海智城分析仪器制造有限公司;HIRAYAMA HVE-50型高压灭菌器 日本Hirayama公司;Delta320pH计 瑞士梅特勒-托利多(上海)有限公司;离子交换色谱仪 美国Dionex公司;Simplicity超纯水系统 美国Millipore公司。
1.3 方法
1.3.1α-半乳糖苷酶的制备
将菌株KLDS2.0509在TPY培养基中连续两次增殖,体积分数5%活化发酵剂接种于200 mL添加1 g/100 mL葡萄糖和1 g/100 mL棉子糖的TPY肉汤培养基中,在37 ℃条件下厌氧培养48 h。在发酵12、24、36、48 h时分别取样50 mL,以6 000×g离心10 min收集菌体,用20 mL冷的50 mmol/L柠檬酸钠缓冲液(pH 5.5,4 ℃)清洗细胞沉淀,接着6 000×g离心10 min,重复以上清洗操作。然后用10 mL相同的缓冲液悬浮细胞沉淀,放置于冰浴中,超声波处理(间歇破碎80次,工作时间3 s,间隔4 s),最后12 000×g离心30 min除去细胞碎片,上述所有离心操作都在4 ℃进行,上层清液用作天然酶提取物[12]。
1.3.2 酶活力测定
取250 μL酶液与500 μL 5 mmol/L的pNPG混合,在37 ℃温育30 min,添加500 μL的0.2 mol/L碳酸钠终止反应,在400 nm波长测定其OD值,以p-NP的生成量表示酶活力。分别测定不同浓度对硝基苯酚标准品的OD400nm,根据测得的吸光度,绘制标准曲线。
酶活力单位(U/mL)定义为:在测定条件下每分钟每毫升pNPG释放1 μmol的p-NP所需的酶量[13]。
1.3.3 酶的纯化
向粗酶液中加入固体(NH4)2SO4至30%的饱和度,4 ℃盐析10 h,离心除去沉淀;然后取上清液继续加入固体(NH4)2SO4至饱和度为60%,4 ℃放置过夜,离心收集沉淀。将沉淀溶于0.01 mol/L的磷酸盐缓冲溶液(pH 6.5)中,然后装入透析袋中,4 ℃透析12 h,每隔3 h更换一次缓冲液;取适量上述透析液加入预先用0.01 mol/L的磷酸盐缓冲液(pH 6.5)平衡过的Sephadex G-100柱中进行洗脱,洗脱速率18 mL/h,分步收集洗脱液,分别测定每管的酶活力。
1.3.4 蛋白质分子质量和浓度的测定
采用LaemmLi法进行SDS-PAGE测定蛋白质分子质量[14];以牛血清白蛋白为标准,采用Lowry法测定蛋白质含量[15]。
1.3.5α-半乳糖苷酶酶学性质研究
1.3.5.1 酶的最适反应pH值及pH值稳定性
分别用pH 2.0~7.0的0.1 mol/L柠檬酸钠缓冲液和pH 8.0~9.0的0.1 mol/L的Tris/HCl缓冲液配制底物pNPG,将酶液分别加入进行酶促反应,测定酶活力,以确定酶的最适pH值;将酶液在不同pH值的缓冲液中于37 ℃条件下分别处理30 min,测定酶活力以研究酶的pH值稳定性[16]。按照1.3.2节方法测定酶活力,以最高酶活力为100%,计算相对酶活力。
1.3.5.2 酶的最适反应温度及热稳定性
在标准酶促反应体系中,在10~70 ℃温度条件下分别测定酶活力,确定此α-半乳糖苷酶的最适作用温度;然后将该酶在35、40、45、50 ℃条件下进行酶促反应,并分别在0.5、1.0、1.5、2.0、2.5 h时测定剩余酶活力[17]。按照1.3.2节方法测定酶活力,以最高酶活力为100%,计算相对酶活力。
1.3.5.3 不同金属离子对酶活力的影响
在酶促反应体系中分别加入金属离子Na+、Ca2+、Fe2+、K+、Li+、Zn2+、Mn2+、Mg2+、Hg2+、Cu2+、Ag+,使金属离子终浓度为1.0 mmol/L,在37 ℃条件下进行酶促反应,以未加入金属离子的酶液做对照,考察金属离子对酶活力的影响[18]。以未处理酶液的酶活力为100%,计算相对酶活力。
1.3.5.4α-半乳糖苷酶的底物特异性测定
将蜜二糖、棉子糖、水苏糖和瓜尔豆胶溶解于0.1mol/L McIlvaine缓冲液(pH 5.0)中,蜜二糖、棉子糖、水苏糖质量浓度各1 mg/mL,瓜尔豆胶为3 mg/mL。使用2 U的α-半乳糖苷酶分别与1 mL这些底物在40 ℃条件下作用24 h。使用离子交换色谱仪测定半乳糖的释放量[19]。
1.4 统计分析
所有数据均为3个重复样品的平均值,采用SPSS13.0软件进行统计分析,判断彼此间的差异显著性。
2 结果与分析
2.1 作用时间对酶活力测定的影响
根据对硝基苯酚标准品的标准曲线获得回归方程:y=0.018 2x+0.002 2(R2=0.999 8)。在酶反应的最初阶段,在反应开始后的2~10 min,反应速率最快,反应体系中对硝基苯酚的含量呈线性增加,继续反应,反应速率逐渐减缓,如图1所示。在酶化学反应过程中,酶反应的瞬时速率很难准确测定,并且反应后期反应速率下降,因此通常在酶反应的最初阶段测定初始反应速率。在本实验中,反应时间确定为10 min。

图1 酶反应的时间进程Fig.1 Time course of the α-galactosidase-catalyzed reaction
2.2 发酵过程中α-半乳糖苷酶酶活力变化曲线
菌株KLDS2.0509在pH6.0 TPY培养基中37 ℃条件下厌氧培养48 h,按照1.3.2节方法测定酶活力,以最高酶活力为100%,计算相对酶活力,其生长曲线和酶活力变化曲线如图2所示。菌株KLDS2.0509在培养4 h后进入指数生长期,在培养12 h后达到稳定生长期,α-半乳糖苷酶进入对数生长期后开始产生,进入稳定期后酶活力逐渐提高,在稳定期中期(24 h)酶活力到达最高值,继续培养酶活力呈现下降趋势,稳定期后期酶活力保持稳定。
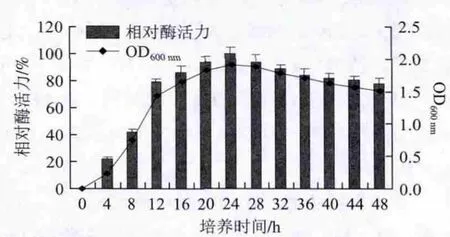
图2 在培养48 h期间KLDS2.0509所产α-半乳糖苷酶活力和OD600nm的变化Fig.2 Relative activity of α-galactosidase produced by KLDS2.0509 and its optical density at 600 nm as a function of culture time
2.3 酶分离纯化
粗酶液先经(NH4)2SO4盐析,再通过透析除杂浓缩,最后浓缩液经Sephadex G-100凝胶柱层析,结果如图3所示。整个洗脱过程出现3个蛋白峰,通过测定各管酶活力,确定α-半乳糖苷酶主要集中在第1个蛋白峰内,说明经过Sephadex G100凝胶柱层析除去部分小于其分子质量的蛋白。

图3 Sephadex G-100凝胶柱层析洗脱曲线Fig.3 Elution curves of Sephadex G-100 column chromatography
粗酶液经过(NH4)2SO4盐析、透析、Sephadex-G100层析处理后,经12.5%的SDS-PAGE电泳检测,结果如图4所示,纯化后仅有一条蛋白条带,说明纯化效果较好。根据相对迁移率分子质量的关系,可以计算出其分子质量约为45kD。

图4 α-半乳糖苷酶SDS-PAGE电泳图谱EFig.4 SDS-PAGE of α-galactosidase at different separation and purification steps
α-半乳糖苷酶的纯化结果如表1所示,粗酶液的酶比活力1.69 U/mg,酶活力回收率为20.6%,纯化倍数为29.3,纯化酶的酶比活力为49.6 U/mg。实验结果与已报道的1株短双歧杆菌的α-半乳糖苷酶活相近,其粗酶液酶比活力为1.76 U/mg[20],但是低于产α-半乳糖苷酶曲霉的粗酶液酶比活力(1.91 U/mg)[8],也低于产α-半乳糖苷酶青霉的粗酶液酶比活力(9.6 U/mg)[19],这与产α-半乳糖苷酶微生物的来源有关[21-22]。

表1 α-半乳糖苷酶的分离纯化Table 1 Purification of α-galactosidase from Bifidibacterium longum KLDS2.0509
2.4 α-半乳糖苷酶酶学性质
2.4.1α-半乳糖苷酶的最适pH值和pH值稳定性
如图5所示,该α-半乳糖苷酶的最适作用pH值为5.0,该酶在pH 4.5~6.0保持80%以上的相对酶活力。将该酶在不同pH值范围的缓冲液中37 ℃作用6 h后测其剩余酶活力,结果如图6所示,该酶在pH 5.0~6.0时最稳定,酶促反应6 h后酶活力仍保持在85%以上,随着pH值升高或者降低,稳定性均呈下降趋势。

图5 α-半乳糖苷酶最适作用pH值Fig.5 Optimum pH of the α-galactosidase
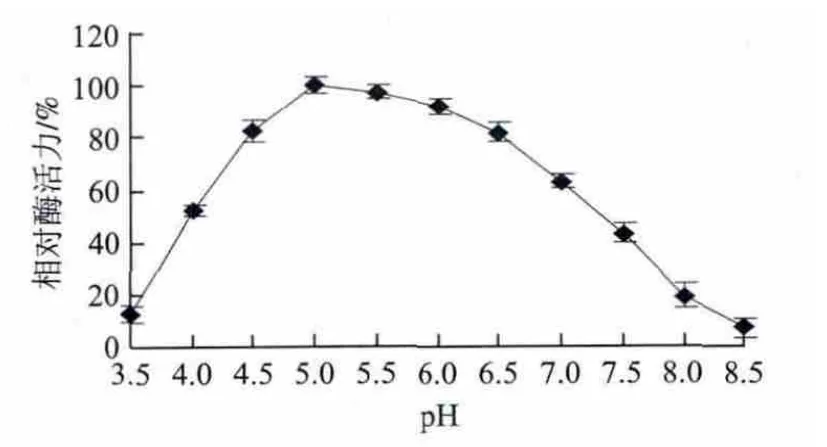
图6 α-半乳糖苷酶的pH值稳定性Fig.6 pH stability of the α-galactosidase
2.4.2α-半乳糖苷酶的最适温度和热稳定性
菌株LB21所产α-半乳糖苷酶的最适作用温度为42 ℃,如图7所示。随着酶促反应温度的升高,α-半乳糖苷酶酶活力逐渐增强,42 ℃达到最高值,温度继续升高,酶活力显著降低。

图7 α-半乳糖苷酶的最适作用温度Fig.7 Optimum temperature of the α-galactosidase
将该酶在不同的温度下作用2.5 h,然后测定剩余酶活力,结果如图8所示。该酶在40 ℃时,酶活力随着作用时间延长,呈现缓慢下降趋势,但是作用2.5 h后仍然保持65%的酶活力;当该酶在35 ℃时,作用2.5 h后,仍剩余82%的酶活力,表明该酶具有良好的热稳定性(P<0.05)。该酶在50 ℃条件下随着时间的增加,酶活力迅速下降,1.5 h时完全丧失酶活性。因此该酶对50 ℃以上的高温较为敏感,但是在42 ℃以下进行酶促反应时,具有较高热稳定性。

图8 α-半乳糖苷酶的热稳定性Fig.8 Temperature stability of the α-galactosidase
2.4.3 不同金属离子对α-半乳糖苷酶的影响
金属离子对该α-半乳糖苷酶酶活力的影响如图9所示,大多数的金属离子对α-半乳糖苷酶的活性没有明显影响,Fe2+、Mn2+、Ag+和Cu2+对其酶活性有一定抑制作用,Cu2+对其酶活性的抑制最显著,相对酶活力为21.2%,而Hg2+完全抑制α-半乳糖苷酶的活性,表明该酶的活性中心部位可能存在含巯基的氨基酸(如半胱氨酸)。

图9 各种金属离子对α-半乳糖苷酶活力的影响Fig.9 Effects of various metal ions on α-galactosidase activity
2.4.4α-半乳糖苷酶的底物特异性分析
如表2所示,底物特异性测定表明该酶可以有效降解含α-半乳糖苷键的蜜二糖、棉子糖和水苏糖,但不能降解末端含α-半乳糖苷键的瓜尔豆胶,对3种底物的降解能力依次为蜜二糖>水苏糖>棉子糖。结果表明该酶具有降解豆制品中α-半乳糖苷类的寡糖的潜力,可用于消除或降低此类寡糖的抗营养作用,从而提高豆制品的利用率,增加豆制品的可食用人群。

表2 α-半乳糖苷酶水解蜜二糖、棉子糖、水苏糖后半乳糖的释放量Table 2 Release of galactose from melibiose, raffinose and stachyose hydrolyzedd bbyy α-galactosiiddaassee
3 结 论
长双歧杆菌KLDS2.0509α-半乳糖苷酶分子质量约为45 kD,纯化后的酶比活力为49.6 U/mg,最适作用pH值为5.0,酶活力随着pH值升高或者降低,均呈下降趋势;最适温度42 ℃,温度继续升高,酶活力显著下降。大多数的金属离子对酶的活性无显著影响,Fe2+、Mn2+、Ag+和Cu2+能够对其产生抑制作用,而Hg2+完全抑制酶活性;酶的底物特异性实验结果表明,该酶可降解蜜二糖、棉子糖和水苏糖,但不能降解末端含α-半乳糖苷键的多糖。该酶在40 ℃条件下保温2.5 h后仍然保持65%的酶活力,表明该酶具有良好的热稳定性。
[1]CHROST B, SCHMITZ K.Purification and characterization of multiple forms ofα-galactosidase inCucumis meloplants[J].Journal of Plant Physiology, 2000, 156(4): 483-491.
[2]FUJIMOTO Z, KANEKO S, KIM W, et al.The tetramer structure of the glycoside hydrolase family 27α-galactosidase I fromUmbelopsis vinacea[J].Bioscience Biotechnology and Biochemistry, 2009, 73(10):2360-2364.
[3]梁素钰, 郑学勤.具血型转换功能的咖啡α-半乳糖苷酶基因的克隆与表达[J].遗传, 2005, 27(5): 759-764.
[4]KATROLIA P, JIA Huiyong, YAN Qiaojuan, et al.Characterization of a protease-resistantα-galactosidase from the thermophilic fungusRhizomucor mieheiand its application in removal of raffinose family oligosaccharides[J].Bioresource Technology, 2012, 110: 578-586.
[5]LILLA S D, NGUYEN Q D, DOMINIKA D L, et al.Effects of galactomannan as carbon source on production ofα-galactosidase byThermomyces lanuginosus: fermentation, purification and partial characterization[J].Enzyme and Microbial Technology, 2009, 45(5):367-371.
[6]SIMERSKA P, MONTI D, CECHOVA I, et al.Induction and characterization of an unusualα-D-galactosidase fromTalaromyces fl avus[J].Journal of Biotechnology, 2007, 128(1): 61-71.
[7]许尧兴, 李艳丽, 柳永, 等.黑曲霉变种RM48α-半乳糖苷酶的分离纯化及其酶学性质研究[J].浙江大学学报: 农业与生命科学版,2009, 35(2): 147-152.
[8]王剑锋, 王璋, 李江, 等.黑曲霉耐酸性α-半乳糖苷酶的分离及其酶学性质[J].食品科学, 2012, 33(21): 267-270.
[9]李苏红, 朱旻鹏, 李拖平.重组水稻α-半乳糖苷酶的分离纯化及酶学性质研究[J].食品科学, 2010, 31(21): 304-307.
[10]郝桂娟, 张凯, 王学智, 等.α-半乳糖苷酶的研究进展[J].中国畜牧兽医, 2013, 40(3): 149-154.
[11]陈俊亮, 杨丽杰, 霍贵成.产α-半乳糖苷酶乳酸菌的鉴定及其发酵性能研究[J].山东大学学报: 理学版, 2008, 43(7): 28-39.
[12]SCALABRINI P, ROSSI M, SPETTOLI P, et al.Characterization ofBifidobacteriumstrains for use in soymilk fermentation[J].International Journal of Food Microbiology, 1998, 39: 213-219.
[13]TSANGALIS D, SHAH N P.Metabolism of oligosaccharides and aldehydes and production of organic acids in soymilk by probiotic bifidobacteria[J].International Journal of Food Science and Technology, 2004, 39: 541-554.
[14]LAEMMLI U K.Cleavage of structural proteins during the assembly of the head of bacteriophage T4[J].Nature, 1970, 227: 680-685.
[15]LOWRY O H, ROSEBROUGH N J, FARR A L, et al.Protein measurement with the Folin phenol reagent[J].The Journal of Biological Chemistry, 1951, 193(1): 265-275.
[16]DONKOR O N, HENRIKSSON A, VASILJEVIC T, et al.α-Galactosidase and proteolytic activities of selected probiotic and dairy cultures in fermented soymilk[J].Food Chemistry, 2007, 104: 10-20.
[17]LEBLANC J G, SILVESTRONI A, CONNES C.Reduction of nondigestible oligosaccharides in soymilk: application of engineered lactic acid bacteria that produceα-galactosidase[J].Genetics and Molecular Research, 2004, 3(3): 432-440.
[18]CARRERA-SILVA E A, SILVESTRONI A, LEBLANC J G, et al.A thermostableα-galactosidase fromLactobacillus fermentumCRL722:genetic characterization and main properties[J].Current Microbiology,2006, 53: 374-378.
[19]密士军, 柏映国, 孟昆.一种来源于青霉的新的α-半乳糖苷酶的分离纯化及其酶学性质[J].微生物学报, 2007, 47(1): 156-160.
[20]肖敏, 刘树峰, 朱崇日, 等.短双歧杆菌α-D-半乳糖苷酶的纯化及性质[J].中华微生物学和免疫学杂志, 2001, 21(3): 307-311.
[21]李孝辉, 陈声明.生物源α-半乳糖苷酶的研究进展[J].微生物学通报, 2002, 29(2): 71-75.
[22]DONKOR O N, HENRIKSSON A, VASILJEVIC T, et al.α-Galactosidase and proteolytic activities of selected probiotic and dairy cultures in fermented soymilk[J].Food Chemistry, 2007, 104(1): 10-20.
猜你喜欢
食品工业(2022年2期)2022-03-09
吉林农业(2019年3期)2019-01-06
中成药(2018年5期)2018-06-06
中成药(2017年8期)2017-11-22
药学与临床研究(2015年4期)2015-06-05
中国当代医药(2015年20期)2015-03-01
中国药业(2014年12期)2014-06-06
中国药业(2014年19期)2014-05-17
天然产物研究与开发(2014年3期)2014-04-27
食品工业科技(2014年13期)2014-03-11
