
奖赏预期对面孔情绪加工的影响:一项事件相关电位研究*
2014-02-02 03:48康冠兰丁锦红郭春彦
心理学报 2014年4期
魏 萍 康冠兰,2 丁锦红 郭春彦
(1首都师范大学心理系, 北京市“学习与认知”重点实验室, 北京 100048)
(2北京大学心理学系, 北京 100871)
1 引言
当我们的某种行为是为了努力达到某个价值目标时, 就会出现激励性动机(incentive motivation)。激励可以是初级奖赏, 比如食物; 也可以是次级奖赏,比如金钱(Pessoa & Engelmann, 2010; Wise, 2004)。人类功能磁共振成像(functional Magnetic Resonance Imaging, fMRI)研究发现, 脑内的奖赏系统主要包含两条通路:边缘通路(mesolimbic pathway)和脑皮层通路(mesocortical pathway)(Pessoa & Engelmann, 2010; 魏萍, 康冠兰, 2012)。边缘通路是从腹侧被盖区(Ventral Tegmental Area,VTA)经内侧前脑束(Medial Forebrain Bundle)到伏隔核(Nucleus Accumbens); 脑皮层通路是连接腹侧被盖区和脑皮层(尤其是额叶皮层)的通路(Chiew &Braver, 2011)。例如, 动物研究发现, 腹侧被盖区多巴胺神经元对有奖赏预期的线索放电会增强(Schultz, Dayan, & Montague, 1997), 从而加强对额叶的信号输入。此外, 事件相关电位(Event-Related Potentials, ERPs)研究发现, 在视觉搜索任务中, 不同的奖赏预期诱发的 ERP成分不同。相对于低奖赏预期的目标, 高奖赏预期的目标所诱发的 N2pc成分出现得更早, 而且波幅更大(Kiss, Driver, &Eimer, 2009)。这些研究都说明, 对奖赏的期待能够提高被试脑内的准备状态, 从而促进被试对后续刺激的行为反应(Bijleveld, Custers, & Aarts, 2010;Engelmann & Pessoa, 2007; Locke & Braver, 2008)。
在人类的社会生存过程中, 获得奖赏常常跟愉快和满足感等正性情绪相关, 提示情感可能是奖赏的重要心理成分。近期一些研究试图考察情感是如何与奖赏相联系的。例如, Berridge和 Robinson(2003)发现, 奖赏能够产生积极的情绪体验。奖赏可能与正性情绪拥有共同的神经基础, 阅读故事并想象愉快的情景会激活脑内奖赏回路(Costa, Lang,Sabatinelli, Versace, & Bradley, 2010); 热恋期的人观看爱人的图片也可以激活奖赏回路(Aron et al.,2005)。功能磁共振成像研究发现, 加工情绪性信息的杏仁核在奖赏加工中也发挥重要作用(Murray,2007)。杏仁核对其他脑区有广泛的投射, 如腹侧纹状区(Ventral Striatum) (Alheid, 2003)、VTA (Amaral& Price, 1984)等区域。杏仁核在情绪性加工和奖赏信息的加工中均有激活, 说明奖赏与情绪之间可能是相互影响的(Baxter & Murray, 2002)。但是, 情绪加工与奖赏系统相互影响的认知和神经机制仍不清楚, 需要进一步的研究。
在人类的日常生活中, 情绪的表达可以通过很多种方式, 如身体姿势、头的姿势以及面孔表情等,其中最鲜明的情绪线索是面孔表情。情绪面孔包含了许多社会性信息, 快速准确地识别出面孔情绪表情, 对人类的生存十分重要(Darwin, 1998)。有研究表明, 人类(以及其他灵长类动物)对面孔表情的识别有先天的基础, 尤其是对负性情绪表情 (Öhman,1993)。情绪表情的加工可能是一种自动的前注意的加工(例如, Adolphs, 2002; Dimberg & Öhman,1996; Eastwood, Smilek, & Merikle, 2003)。正性情绪传递出一种友好的信息, 在日常生活中比较普遍。而负性情绪传达出敌意或者威胁信息, 在社会生存中非常重要。人们对正性情绪的偏向性加工存在个体差异(Fox, Russo, & Dutton, 2002)。但是, 情绪加工中的负性偏向, 即人们能够快速地探测负性的、威胁性的刺激, 得到了较为一致的实验结果(Hansen & Hansen, 1988; Morris, Öhman, & Dolan,1998; Öhman & Mineka, 2001; Pinkham, Griffin,Baron, Sasson, & Gur, 2010)。电生理学研究证据指出, 负性效价的图片刺激能够诱发更大的P1、LPP(Late Positive Potentials)成分(Ito, Larsen, Smith, &Cacioppo, 1998; Smith, Cacioppo, Larsen, & Chartrand,2003), 支持了情绪刺激加工的负性偏向。此外, 在情绪面孔的加工过程中, 左右半球存在对正性或负性情绪的单侧化加工优势(Adolphs, 2002)。情绪加工的效价理论认为, 右半球加工负性情绪有优势,左半球加工正性情绪有优势。例如, Adolphs, Jansari和 Tranel (2001) 的研究发现右半球损伤将降低对呈现在视野左侧的悲伤面孔的辨别准确率, 而左半球损伤将降低对呈现在右视野的高兴面孔的辨别准确率。
以往关注奖赏与情绪相互关系的研究都是单独地考察奖赏或者情绪对认知加工的影响, 在情绪加工中发现了奖赏相关的大脑区域(Aron et al.,2005; Costa et al., 2010), 或者在奖赏预期中发现了情绪相关的加工机制(Murray, 2007)。近期一些研究试图在同一个实验研究中同时操纵奖赏和情绪, 来考察奖赏和情绪之间的相互作用。例如, Wittmann,Schiltz, Boehler和Düzel (2008)利用功能磁共振成像技术考察了情绪效价和奖赏引起的中脑边缘系统的交互作用对记忆的影响。研究发现, 奖赏只增强了正性情绪条件下的记忆, 而对中性或负性情绪条件下的记忆没有影响。此外, 与奖赏相关的纹状区的激活增强了对正性刺激的表征, 而不是对负性刺激的表征。近期, Shigemune等人(Shigemune et al.,2010)利用正电子发射断层扫描(Positive Emission Tomography, PET)技术研究情绪和奖赏对情景记忆编码的影响。实验中使用了中性和负性刺激, 没有发现奖赏和情绪在行为表现上的交互作用。但是,影像学数据分析显示,在情景记忆编码过程中, 右侧海马可能整合了情绪和奖赏的效应。
综上所述, 奖赏和情绪都是具有情感显著性(affective significance)的刺激, 都会对认知控制产生影响(Pessoa, 2009), 同时奖赏和情绪又存在相互作用。那么, 一个重要的问题是, 奖赏预期是否能够直接影响人脑对情绪信息的加工?为探究这一问题, 本研究采用线索-目标范式和事件相关电位技术来考察奖赏预期影响情绪面孔识别的认知和神经机制。线索-目标范式能够有效地实现心理预期(Engelmann & Pessoa, 2007; Onoda et al., 2006;Veling & Aarts, 2010)。大量研究面孔表情加工的ERP实验主要探讨了对面孔加工敏感的N170、VPP(Vertex Positive Potential) 和N300成分在呈现不同面孔表情以及不同实验任务时表现出来的差异。
N170是在面孔刺激呈现后 170 ms左右出现的、分布在枕颞区的负成分, 该成分可能提供了对面孔结构快速编码的一种指示, 被认为是编码面孔结构的特异性成分(Bentin, Allison, Puce, Perez, &McCarthy, 1996; Itier & Taylor, 2004a, b, c)。研究指出, 相比左半球的N170, 右半球的N170波幅更大且潜伏期更短(Bentin et al., 1996)。而N170是否受面孔情绪表情的调节仍然存在很大的争议。有研究指出, N170会受到面孔情绪表情的影响, 相比中性面孔, 负性面孔诱发更大的 N170, 尤其是恐惧的表情(Batty & Taylor, 2003; Blau, Maurer, Tottenham,& McCandliss, 2007)。但也有研究发现, N170不受面孔情绪表情的调节(Eimer & Holmes, 2002;Ashley, Vuilleumier, & Swick, 2004)。这些不一致的研究结果, 可能受到刺激材料、面孔呈现方式等因素的影响。
VPP是在额中央电极点记录到的正成分, 其潜伏期类似于N170的潜伏期, 且与N170类似, 对面孔加工敏感(Eimer, 2000)。有研究认为, VPP与N170具有相同的皮层发生源, 是N170在额中央区的极性反转(Joyce & Rossion, 2005); 但是也有研究认为VPP和N170的皮层发生源是独立的(Bentin et al., 1996; Eimer, 2000)。VPP可能受到面孔情绪的调节, 情绪面孔比中性面孔诱发更大的 VPP波幅(Luo, Feng, He, Wang, & Luo, 2010; Williams,Palmer, Liddell, Song, & Gordon, 2006)。
N300是面孔刺激呈现300 ms左右的一个负成分。研究发现, 非面孔刺激并未诱发该成分或者诱发非常小的N300 (Botzel & Grusser, 1989), 且负性情绪性刺激诱发的 N300更大(Carretié & Iglesias,1995; Luo et al., 2010)。N300可能反映了对相关视觉刺激的更深入地加工(Carretié & Iglesias, 1995;Carretié, Iglesias, García, & Ballesteros, 1997; Luo et al., 2010)。
本研究利用线索-目标范式, 通过对线索类型(奖赏预期、无奖赏预期)的操纵实现对奖赏预期的操纵, 考察奖赏预期对目标情绪面孔识别的影响,并在被试反应之后给予相应的反馈(是否获得奖赏)。奖赏线索作为一种动机性因素会以一种“自上而下”的方式提高被试的准备状态并调节被试对后续目标的注意, 使得被试优先加工与奖赏相关的刺激。事件相关电位技术具有较高的时间分辨率, 能够考察线索引发的注意准备信号如何受到奖赏预期的影响, 以及不同的奖赏预期如何影响情绪面孔加工在各个时程上的特异性加工成分。
我们假设, 在行为水平上, 奖赏预期会提高被试在情绪辨别任务中的行为反应, 被试在奖赏预期条件下的反应时快于无奖赏预期条件下的反应时。情绪面孔加工可能具有优先性, 对情绪面孔的反应时快于对中性面孔的反应时。重要的是, 奖赏预期与情绪可能存在交互作用, 不同情绪条件下的奖赏效应(奖赏条件的反应时减去无奖赏条件下的反应时)存在差异。在神经机制水平上, 首先, 相对于无奖赏预期条件, 线索在奖赏预期条件下可能诱发更大的ERP波幅。其次, 在目标加工阶段, 由于之前的奖赏线索使得大脑进入不同的准备状态, 相对于无奖赏预期条件, 奖赏预期条件下大脑对目标面孔的神经反应会增强, 产生更大的波幅。根据以往对情绪面孔加工的 ERPs研究结果可预测, 目标面孔将会诱发枕颞分布的P1、N170, 以及额中央分布的VPP、N300等成分。由于奖赏引发的动机能够集中注意, 我们预期早期的 P1、N170、VPP可能会受到奖赏预期的影响。此外, 由于情绪加工过程中存在半球优势效应, 在左侧和右侧头皮位置记录到的情绪面孔诱发的 ERP成分也可能存在差异。在右侧头皮位置上, 情绪(正性和负性)面孔诱发的N170、VPP和N300波幅可能大于左侧头皮位置上记录到的ERP波幅。重要的是, 奖赏与情绪可能存在交互作用(Murray, 2007; Wittmann et al., 2008;Shigemune et al., 2010)。我们预期, 晚期的N300波幅可能会受到奖赏与情绪的交互影响。由于人们对负性面孔的加工偏向, 奖赏对负性面孔加工的促进效应可能大于对其他面孔加工的促进效应。
2 方法
2.1 被试
20名本科生或研究生参与了本实验(男性10人,年龄18~26), 所有被试都签署了实验知情同意书。被试均为右利手, 视力或者矫正视力正常, 身体健康, 无情绪障碍疾病病史。完成实验后, 被试均可获得一定的报酬, 包括基本报酬和可能的额外奖励。
2.2 实验材料和仪器
实验中共使用90张不同人的面孔的黑白图片,目标面孔图片来自中国情绪面孔图片系统(Chinese Facial Affective Picture System, CFAPS; 王妍, 罗跃嘉, 2005)。我们采用 CFAPS是为了避免中国被试使用西方人情绪面孔图片系统造成的文化差异。根据面孔表情的愉悦度、唤醒度和优势度分别选择了正性(高兴)、中性和负性(愤怒)面孔图片各30张,每种类型的面孔的性别男女比例是 1:1。目标面孔在明度、对比度方面都进行了匹配。正性和负性面孔图片在唤醒水平上进行了匹配, 两种类型的图片在唤醒度上没有差异[M
±SD
: Positive = 6.2 ± 0.75;Negative = 6.0 ± 1.10]。中性面孔图片的唤醒度为3.4 ± 0.31。三类面孔图片在效价这个维度上差异显著[M
±SD
: Positive = 6.6 ± 0.47; Negative = 2.9 ±0.39; Neutral = 4.6 ± 0.21,p
< 0.001]。此外, 我们招募了另外 17个被试(男性 6人, 年龄 20~28周岁)对面孔的表情强度进行了评价。结果显示, 愤怒面孔的表情强度(59%)与高兴面孔的表情强度(61%)无显著差异(p
> 0.1); 愤怒和高兴面孔的表情强度均显著大于中性面孔的表情强度(12%),t
(16) =19.92,p
< 0.001, 以及t
(16) = 17.23,p
< 0.001。实验刺激在17寸纯平CRT显示器上呈现, 分辨率为1024 × 768。实验刺激的呈现和被试反应数据的记录用 Presentation (http://nbs.neuro-bs.com/)软件实现。被试眼睛距离屏幕65cm。
2.3 实验程序
实验操控奖赏预期(奖赏预期、无奖赏预期)与目标面孔情绪(正性、中性、负性)两个因素, 构成6个实验条件:奖赏预期-正性面孔, 奖赏预期-中性面孔, 奖赏预期-负性面孔, 无奖赏预期-正性面孔,无奖赏预期-中性面孔, 无奖赏预期-负性面孔。正式实验之前, 被试需要完成一个练习, 共 24个试次。让被试完成练习实验, 一方面是为了让被试熟悉实验程序, 另一方面是为了获得每个被试对面孔情绪反应的平均反应时作为该被试的基线反应时(Small et al., 2005)。
正式实验时, 被试静坐于安静的实验室内, 眼睛注视屏幕中央。刺激是呈现在一个黑色的背景上。实验的刺激序列如图 1所示(实际实验中是用黑色背景, 线索及注视点均为白色)。每个试次的开始呈现一个中央注视点“+” (500 ms)。接下来是线索阶段, 中央线索(¥、#, 视角是 3°×3°)呈现时间1000 ms, 线索指示该试次的奖赏属性。当线索为¥,即, 奖赏相关试次, 说明对当前试次又快又好的反应与额外奖赏相关; 当线索为#, 即, 奖赏无关试次, 说明对当前试次的反应与额外奖赏无关。在练习后、正式实验前告知被试, 在奖赏相关试次中,当被试又快又好地(反应正确, 且比基线反应时快)完成的奖赏试次达到75 %, 即135个(奖赏试次总数为180个), 则可以获得额外的15元奖励。基线反应时是被试完成练习试次的平均反应时。中央线索呈现后600~1000 ms的可变间隔后, 屏幕中央呈现目标面孔 300 ms, 每张面孔的大小是 4.93°×5.99°。被试的任务是判断面孔的情绪表情, 进行按键反应, 三种表情对应键盘上的三个方向键:例如,正性面孔用右手食指按“左”方向键, 中性面孔用右手中指按“下”方向键, 负性面孔用右手无名指按“右”方向键。按键在被试之间平衡。并要求被试在所有条件下都要又快又准地进行反应。间隔1400~1800 ms后, 呈现反馈画面(500 ms)。在奖赏相关试次中, 被试反应正确且快于基线反应时, 反馈图像为“一元硬币图像”; 反应正确但慢于基线反应时,反馈图像为“实心的灰色圆”, 反应错误时, 反馈图像为为“空心的灰色环”。在无奖赏预期试次, 反应正确时反馈图像为“实心的灰色圆”, 反应错误时反馈图像为“空心的灰色环”。试次之间的间隔为1100~1600 ms。
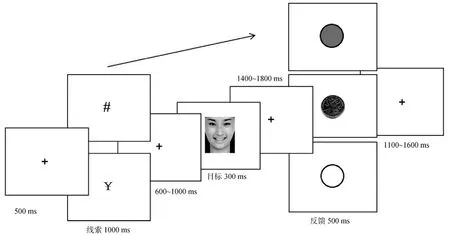
图1 正式实验流程图示例
正式实验共有360个试次, 每个子条件下有60个试次。这些条件平均分配在10个区组, 每个区组中各个实验子条件各有6个试次。每个区组共有36个试次, 以伪随机的方式呈现。练习阶段有24个试次, 刺激的呈现与正式实验相同, 只是在指导语中让被试忽视线索, 集中注意力对面孔情绪进行反应。反馈阶段只会出现正确反馈以及错误反馈, 不会出现奖赏反馈(一元硬币图像)。练习结束后, 屏幕上会呈现被试练习时的正确反应个数, 以及平均反应时。练习阶段记录到的平均反应时则作为正式实验中的基线反应时。
2.4 EEG的记录和分析
实验采用Neuroscan公司的ESI-64导脑电记录系统, 电极位置在国际 10-20系统基础上构成, 用Ag/AgCl电极帽记录62个头皮位置相应的EEG。左眼上下 2个电极记录垂直眼电(VEOG), 两眼外侧 2个电极记录水平眼电(HEOG)。参考电极置于左耳乳突处, 接地点在FPz和Fz连线的中点, 右耳乳突也放置一个电极。每个电极与头皮之间的电阻均小于5 kΩ, 连续记录时滤波带通为0.05~100 Hz,采样率为500 Hz。对测验阶段记录的EEG进行离线分析, 以左右乳突的代表平均为参考电压进行校正。截取每对刺激呈现前100 ms到呈现后1000 ms的脑电, 用-100~0 ms的平均波幅对基线进行校正。以回归法自动校正眼电伪迹, 并剔除波幅超过±75µV的试次, 滤波带通为0.05~40 Hz。
分别对每个条件下反应正确的 EEG活动进行叠加和平均。最终得到由奖赏期待(奖赏预期、无奖赏预期)和面孔情绪(正性、中性、负性)构成的6种类型的ERPs数据(即, 奖赏预期-正性面孔、奖赏预期-中性面孔、奖赏预期-负性面孔、无奖赏预期-正性面孔、无奖赏预期-中性面孔、无奖赏预期-负性面孔)。由于本研究假设, 奖赏线索作为一种动机性因素会以一种“自上而下”的方式调节被试的注意, 使得被试优先加工与奖赏相关的刺激。为了考察被试对奖赏预期线索和无奖赏预期线索的差异性加工, 首先分析线索阶段诱发的ERP成分。以刺激呈现为叠加零点, 分析时程为线索刺激出现后1000 ms, 基线为刺激出现前100 ms。每个被试每个条件至少有50次有效叠加次数。测量P1 (130~200 ms)、P2 (200~300 ms)和 P300 (300~600 ms)成分的平均波幅。P1、P2和P300的测量位点为Fz、Cz、Pz、F3、F4、C3、C4 、P3 和 P4。对测得的平均波幅进行两因素重复测量方差分析。两因素分别为奖赏期待(奖赏预期、无奖赏预期)和电极点位置(见成分的测量位点)。
对目标刺激诱发的ERPs的分析以目标刺激呈现为叠加零点, 分析时程为刺激出现后 1000 ms,基线为刺激出现前100 ms。对所有被试每个电极点位置的ERP成分的观察来看, N170成分在枕颞位置(PO7、PO8)最明显, VPP成分在额中央位置的波幅最大(Fz、Cz)。我们将在相应的电极点位置对以下成分的平均波幅进行重复测量方差分析:枕部(Oz 、O1/2)及枕颞位置(PO7/8)的P1 (80~130 ms),枕颞位置(PO7、PO8)的N170 (130~200 ms), 以及额中央位置(Fz、Cz、F3/4、C3/4)的VPP (130~200 ms)、N300 (200~400 ms)。对P1进行三因素重复测量方差分析, 三因素分别为奖赏预期(奖赏预期、无奖赏预期)、情绪(正性、中性、负性)以及头皮位置(中线:Oz; 左侧:O1、PO7; 右侧:O2、PO8)。其中左侧电压值为O1和PO7的平均电压值, 右侧电压值为O2和PO8的平均电压值。对N170的波幅进行三因素的重复测量方差分析, 三因素分别为奖赏预期(奖赏预期、无奖赏预期)、情绪(正性、中性、负性)以及相应的测量位点(PO7/8)。对 VPP、N300的平均波幅进行三因素的重复测量方差分析,三因素分别为奖赏预期(奖赏预期、无奖赏预期)、情绪(正性、中性、负性)以及头皮位置(中线:Fz、Cz; 左侧:F3、C3; 右侧:F4、C4)。其中, 中线位置的电压值是Fz和Cz的平均值, 左侧的电压值是F3和C3 的平均电压值, 右侧的电压值是F4和C4的平均电压值。
3 结果
3.1 行为结果
按照2(奖赏预期、无奖赏预期) × 3(面孔情绪:正性、中性、负性)的实验设计, 计算了每个子条件下的反应时和错误率(如表1)。其中, 2个被试的错误率超过 20%, 因此我们剔除了这两个被试的数据。另外剔除了每个被试每个子条件下的反应极端值(平均值±三个标准差以外的数据), 共剔除了1.6%的数据。接着我们将平均反应时进行2(奖赏预期、无奖赏预期) × 3(面孔情绪:正性、中性、负性)的重复测量方差分析。方差分析的结果显示:奖赏预期的主效应显著,F
(1, 17) = 36.19,p
< 0.001, 在奖赏预期条件下, 被试对面孔情绪的反应时 (601 ms)显著快于无奖赏预期条件下的反应时(651 ms)。面孔情绪的主效应显著,F
(2, 34) = 25.54,p
< 0.001,被试对正性面孔的反应时(566 ms)显著快于对中性面孔(677 ms)以及负性面孔(636 ms)的反应时(ps
<0.05); 对负性面孔的反应时显著快于对中性面孔的反应时(p
< 0.005)。此外, 我们并没有发现奖赏预期和面孔情绪的交互作用,F
(2, 34) < 1。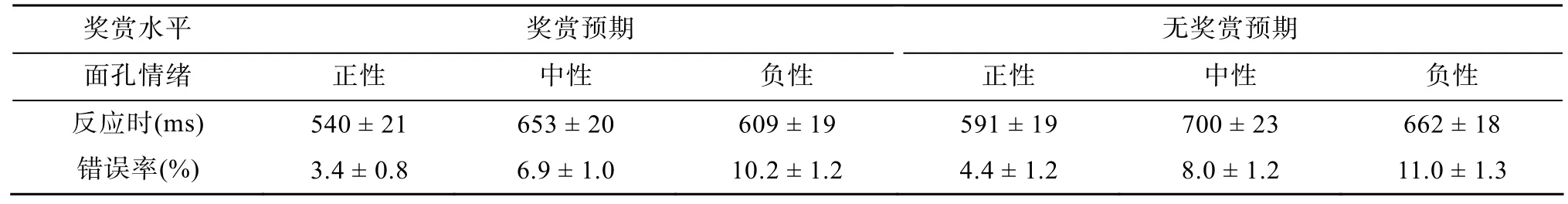
表1 实验各个子条件下反应时 ± 标准误(ms)以及错误率 ± 标准误(%)
对错误率进行了同样的重复测量方差分析显示, 只有情绪的主效应显著,F
(2, 34) = 11.61,p
<0.001, 对负性面孔反应的错误率(10.6%)显著高于对正性面孔反应的错误率(3.9 %)。3.2 ERP数据
3.2.1 线索引起的ERP成分
(1) P1 (130~200 ms)成分
P1波幅奖赏预期的主效应显著,F
(1, 17) =37.33,p
< 0.001, 奖赏预期条件下线索诱发的P1波幅显著大于无奖赏预期条件下的P1波幅(图2)。(2) P2 (200~300 ms)成分
P2波幅奖赏预期的主效应显著,F
(1, 17) =39.24,p
< 0.001, 奖赏预期条件下线索诱发的ERP波幅大于无奖赏预期条件下线索诱发的P2波幅(图2)。(3) P300 (300~600 ms)成分
P300波幅奖赏预期的主效应显著,F
(1, 17) =13.14, p
< 0.005, 奖赏预期条件下线索诱发的P300成分的波幅大于无奖赏预期条件下的P300波幅(图2)。产供集团目前也正为研究堆开发新型燃料。产供集团总裁纳塔利娅·尼基佩洛娃9月25日在一次会议上表示,对于产供集团,研究堆燃料市场的重要性不亚于商业反应堆燃料市场。产供集团在研究堆燃料领域与国外伙伴开展了大规模合作,根据客户的特定需求提供燃料。
3.2.2 目标刺激引起的ERPs成分
(1) P1 (80~130 ms)
对 P1波幅的重复测量方差分析显示, 奖赏预期与头皮位置的交互作用显著,F
(2, 34) = 3.83,p
<0.05。简单效应分析发现, 在左侧头皮位置(O1、PO7)上, 奖赏预期条件下的 P1显著大于无奖赏预期条件下的P1波幅(p
< 0.05); 右侧电极(O2、PO8)及中线电极(Oz)上, 奖赏预期条件与无奖赏预期条件下 P1波幅无差异。此外, 没有其他的主效应与交互作用。(2) N170 (130~200 ms)成分
对 N170波幅的重复测量方差分析表明, 奖赏预期的主效应显著,F
(1, 17) = 4.74,p
< 0.05, 奖赏预期条件下目标诱发的N170比无奖赏预期条件下的N170更正(图3)。奖赏预期与电极点位置的交互作用显著,F
(2, 34) = 3.06,p
< 0.05。进一步简单效应分析发现, 在左侧电极点PO7上, 目标在奖赏预期条件下诱发了更正的N170 (p
< 0.05); 在右侧电极点 PO8上, 奖赏预期与无奖赏预期条件下的N170波幅无显著差异。此外, 没有发现其他的主效应或交互效应。(3) VPP (130~200 ms)成分
对 VPP波幅的方差分析显示, 头皮位置的主效应显著,F
(2, 34) = 5.48,p
< 0.05, 进一步比较发现, 中线位置上的 VPP波幅显著大于左侧和右侧位置上记录到的VPP波幅(ps
< 0.05) (图3)。此外,没有发现其他的主效应与交互作用。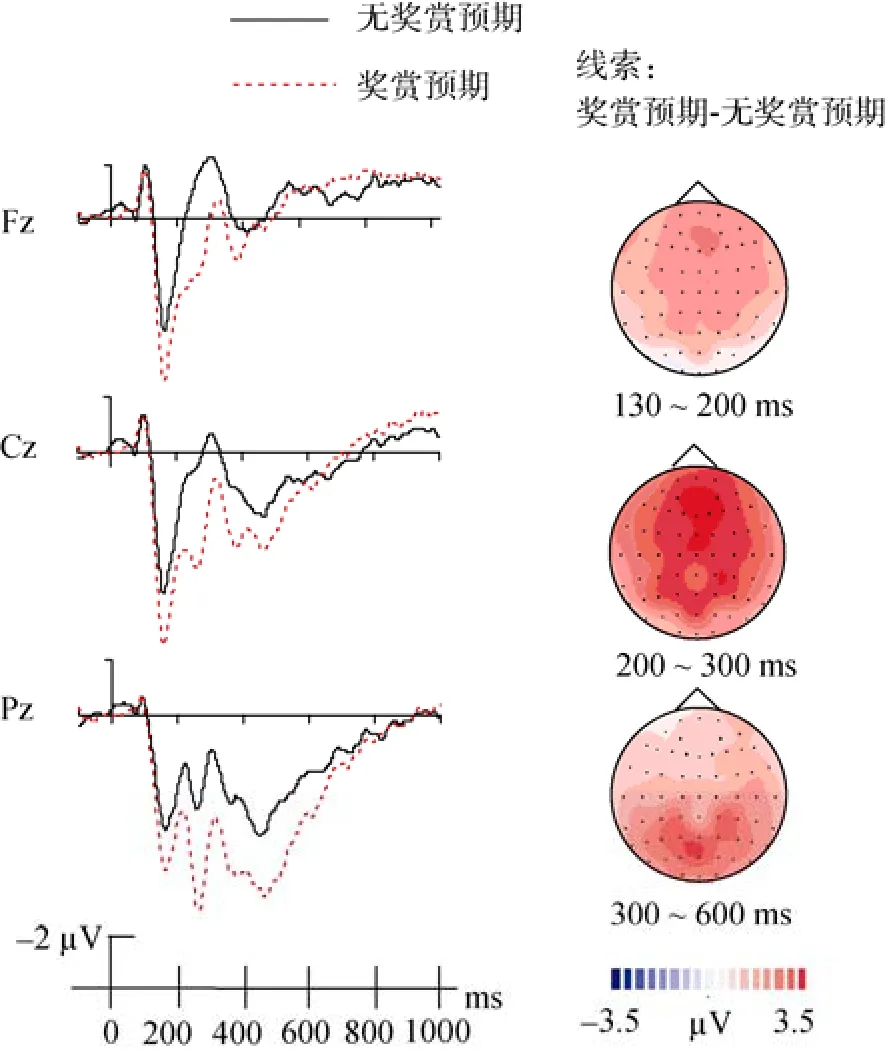
图2 Fz、Cz、Pz电极上记录到的线索诱发的ERP总平均图以及地形图
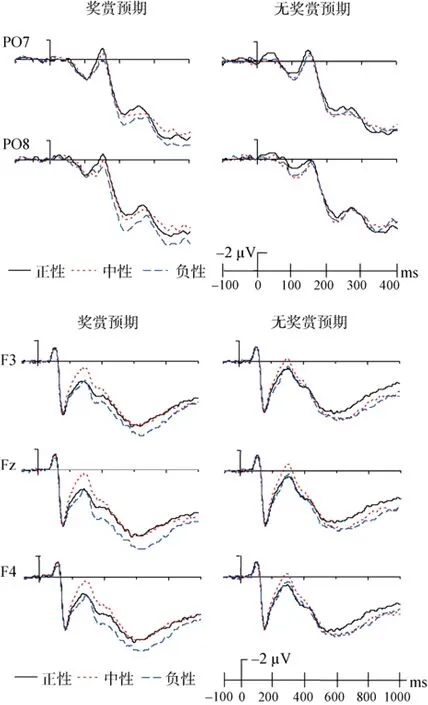
图3 奖赏预期和无奖赏预期条件下, 目标阶段正性、中性和负性情绪面孔诱发的ERPs。
(4) N300 (200~400 ms)成分
对 N300波幅的分析发现, 奖赏预期的主效应显著,F
(1, 17) = 28.64,p
< 0.001, 奖赏预期条件下目标诱发的N300比无奖赏预期条件下的N300更正。情绪的主效应显著,F
(2, 34) = 17.85,p
< 0.001,正性和负性面孔诱发的 N300比中性面孔诱发的N300 更正(ps
< 0.05)。头皮位置的主效应也显著,F
(2, 34) = 5.97,p
< 0.01, 中线位置记录到的N300比右侧头皮位置记录到的N300更正(p
< 0.05)。奖赏预期与头皮位置的交互作用,F
(2, 34) = 6.87,p
<0.005, 情绪与头皮位置的交互作用,F
(4, 68) = 3.51,p
< 0.05, 奖赏预期与情绪的交互作用,F
(2, 34) =5.47,p
< 0.01, 均显著。对奖赏预期与情绪的交互作用进行简单效应分析发现, 奖赏预期条件下, 情绪面孔(正性和负性)诱发的 N300比中性面孔诱发的 N300更正(ps
< 0.01)。同时, 负性面孔诱发的N300比正性面孔诱发的N300更正(p
< 0.005)。而无奖赏预期条件下, 情绪面孔(正性和负性)诱发的N300波幅比中性面孔诱发的N300更正(ps
< 0.05),正性与负性面孔诱发的N300波幅无显著差异(p
>0.05) (图 3)。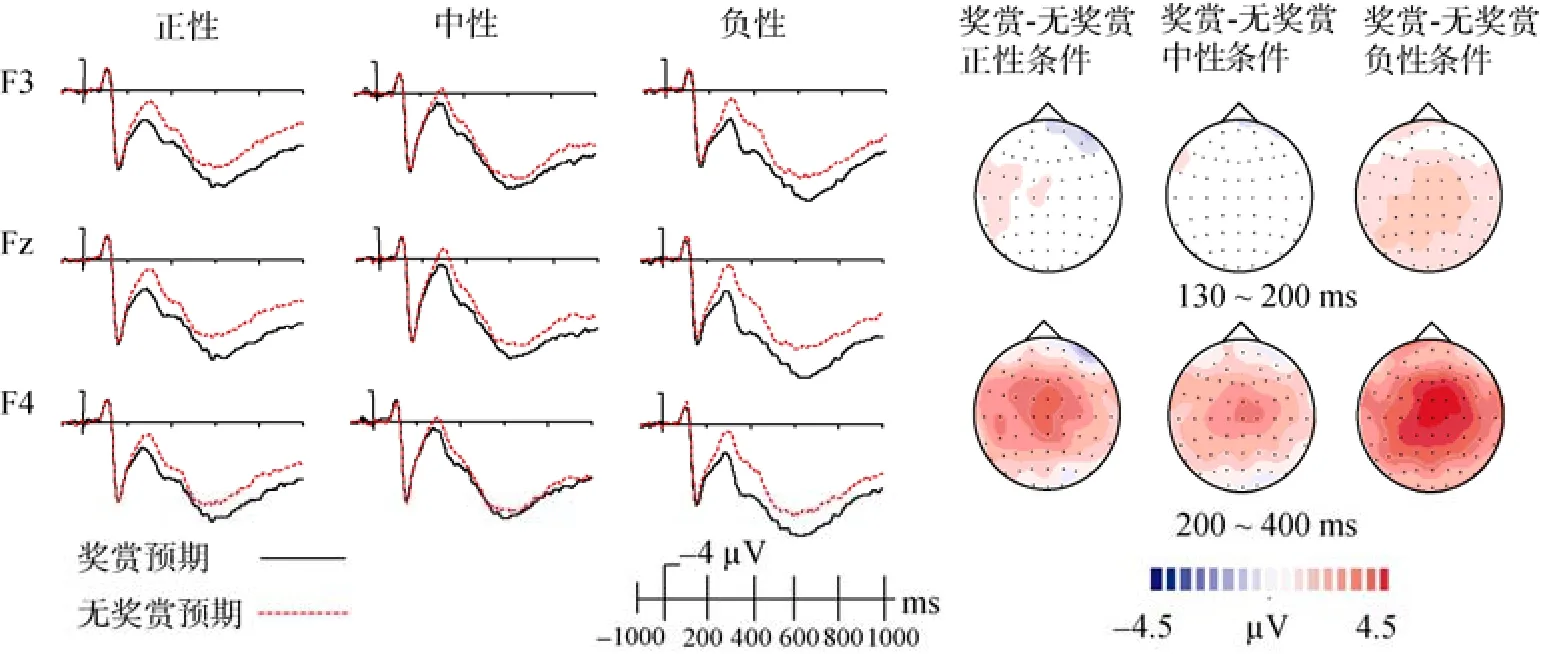
图4 目标阶段正性、中性和负性情绪面孔在不同奖赏预期条件下诱发的ERPs, 以及奖赏预期-无奖赏预期差异波在130~200 ms和200~400 ms的地形图
对正性情绪加工效应的分析结果显示, 头皮位置的主效应显著,F
(2, 34) = 4.19,p
< 0.05, 进一步比较发现, 中线位置的正性情绪加工效应大于左侧头皮位置的正性情绪加工效应,p
< 0.05。此外, 没有出现其他的主效应和交互作用。对负性情绪加工效应的分析结果显示, 奖赏预期的主效应显著,F
(1, 17) = 9.94,p
< 0.05, 奖赏预期条件下的负性情绪加工效应显著大于无奖赏预期条件下的负性情绪加工效应。头皮位置的主效应显著,F
(2, 34) = 4.65,p
< 0.05, 中线位置上的负性情绪加工效应大于左侧头皮位置上的负性情绪加工效应。奖赏预期和头皮位置的交互作用显著,F
(2,34) = 3.75,p
< 0.05。虽然在各个头皮位置上, 奖赏预期条件下的负性情绪加工效应均大于无奖赏预期条件下的该效应, 但是奖赏预期与无奖赏预期条件下的负性情绪加工效应的差异在中线位置上显著大于在左侧位置上的该效应差异(p
< 0.05)。对负性偏向效应的分析结果显示, 奖赏预期的主效应显著,F
(1, 17) = 6.60,p
< 0.05, 奖赏预期条件下的负性偏向效应显著大于无奖赏预期条件下的负性偏向效应。奖赏预期与头皮位置的交互作用显著,F
(2, 34) = 4.09,p
< 0.05。简单效应分析显示,在右侧和中线位置上, 奖赏预期条件下的负性偏向效应显著大于无奖赏预期条件下的负性偏向效应(ps
< 0.05); 在左侧头皮位置上, 奖赏预期与无奖赏预期条件下的负性偏向效应无显著差异(p
>0.1)。此外, 没有出现其他的效应。此外, 我们计算了奖赏效应(即, 不同头皮位置以及不同情绪条件下, 奖赏预期条件下的 N300波幅值减去无奖赏预期条件下的N300波幅值的差值)。对奖赏效应进行3(情绪:正性、中性、负性) ×3(头皮位置:左侧、中线、右侧)的重复测量方差分析。结果显示, 情绪的主效应显著,F
(2, 34) = 5.47,p
< 0.01, 进一步比较发现, 负性条件下的奖赏效应显著大于中性条件下的奖赏效应(p
< 0.05), 负性条件下的奖赏效应大于正性条件下的奖赏效应(p
=0.06), 中性与正性条件下的奖赏效应无显著差异(图4)。头皮位置的主效应显著,F
(2, 34) = 6.87,p
<0.005, 中线位置上的奖赏效应要显著的大于左侧和右侧头皮位置(ps
< 0.05)。此外, 没有发现头皮位置和情绪的交互作用。4 讨论
本研究采用ERPs技术考察了奖赏预期对面孔情绪加工的影响。行为结果显示, 被试在奖赏预期条件下的反应时快于无奖赏预期条件下的反应时;被试对正性面孔的反应时快于对负性面孔或中性面孔的反应时, 并且对正性面孔反应的错误率显著低于负性面孔或中性面孔的错误率。这种对正性情绪加工的有效性, 可能是由于正性面孔在社会生活中更常见、更熟悉, 容易产生知觉启动(Öhman,Lundqvist, & Estenvs, 2001)。同时对负性面孔的反应时显著快于对中性面孔的反应时, 体现了负性情绪的加工优势。ERPs数据显示, 相对于无奖赏预期条件, 在奖赏预期条件下线索诱发了更正的 ERP成分, P1、P2和P300波幅都出现了显著的奖赏预期主效应。在目标阶段, 枕部的 P1、枕颞位置的N170以及额中央位置的N300波幅均受到奖赏预期的调节, 目标在奖赏预期条件下诱发了更正的ERPs。P1、N170和VPP波幅没有受到面孔情绪的影响, 而晚期的 N300显示出情绪面孔与中性面孔加工的差异。重要的是, N300波幅出现了奖赏预期与情绪的交互作用。对N300的情绪效应分析发现,正性情绪加工效应并没有受到奖赏预期的影响, 不同奖赏预期条件下的正性情绪加工效应无显著差异。而负性情绪加工效应以及负性偏向效应均受到奖赏预期的调节, 奖赏预期条件下的负性情绪加工效应和负性偏向效应均显著大于无奖赏预期条件下的负性情绪加工效应和负性偏向效应。此外, 对N300奖赏效应的分析发现, 负性条件的奖赏效应显著大于正性以及中性条件下的奖赏效应。
在线索阶段, 线索类型指示不同的奖赏预期条件, 形成不同的动机状态。额区、中央区、顶区记录的P1、P2和P300波幅受到奖赏预期的调节, 奖赏预期条件下线索诱发的ERPs波幅显著大于无奖赏预期条件下的波幅, 这与近期的一些研究结果一致。例如, Hickey, Chelazzi和Theeuwes (2010)使用额外奇异项注意捕获范式发现, 当客体(目标或奇异项)具有与奖赏相联系的颜色属性时诱发了更大的P1和N2pc成分, 说明奖赏促进了对刺激的知觉登记和注意选择。P2成分指示任务相关刺激的注意加工, 任务相关的刺激比任务无关的刺激诱发更大的 P2 (Potts & Tucker, 2001; Potts, 2004)。Kaltwasser等人(Kaltwasser, Ries, Sommer, Knight, & Willems,2013)采用线索-目标范式也发现了奖赏线索比无奖赏预期条件下诱发了更大的P2。此外, 磁共振成像研究结果显示, 奖赏网络的皮层通路主要在额叶,如眶额皮层(Orbitofrontal Cortex, OFC)、前扣带皮层(Anterior Cingulate Cortex, ACC)等区域(O'Doherty,Kringelbach, Roll, Homak, & Andrews, 2001; Pessoa& Engelmann, 2010), 后扣带皮层(Posterior Cingulated Cortex, PCC)整合了动机与空间注意信息(Mohanty, Gitelman, Small, & Mesulam, 2008;Small et al., 2005)。在头皮额区、中央区以及顶区观察到的奖赏效应可能是因为奖赏线索激活了奖赏系统, 因而在相应的头皮部位产生了更强的神经元放电反应。大脑对奖赏预期线索产生了更强的神经反应, 有利于调动更多的注意资源, 使之进入更好的准备状态, 促进对目标的加工。
目标面孔诱发了早期的 P1成分。相对于其他类型的图片或随机面孔图片(scrambled faces), 面孔图片诱发的P1更大(Herrmann, Ehlis, Ellgring, &Fallgatter, 2005), 而 P1是否受到情绪的调节存在不一致的结果。一些研究发现, P1成分受到情绪信息的调节。例如, Batty和Taylor (2003)将六种基本情绪面孔与其他客体图片(汽车、飞机等)以随机顺序呈现给被试, 被试只需对其他客体图片按键反应。结果发现, 愤怒、高兴、悲伤、厌恶、恐惧等情绪面孔图片诱发的 P1显著大于中性面孔和惊奇面孔诱发的P1。Luo等人(2010)采用快速系列视觉呈现(Rapid Serial Visual Presentation, RSVP)范式和注意瞬脱(Attentional Blink)任务, 结果发现当目标二(T2)为恐惧面孔时比中性面孔和高兴面孔时诱发更大的 P1。但是, Rossignol等人(Rossignol,Philippot, Douilliez, Crommelinck, & Campanella,2005)采用 Oddball范式考察了高低焦虑人群对恐惧、高兴、中性面孔图片的ERP反应, 却没有发现P1受到焦虑人群或面孔情绪的调节。针对这些不一致的结果, 一个可能的解释是, P1反映了对情绪性信息或显著(salience)信息的快速提取(Vuilleumier& Pourtois, 2007)。在本研究结果中, P1虽没有受到面孔情绪的调节, 却受到了奖赏预期的调节, 目标面孔在左侧后部头皮位置上诱发的 P1波幅在奖赏预期条件下显著大于无奖赏预期条件下的波幅。这可能是由于, 此时奖赏预期具有更高的行为相关性(或称显著性), 被试对奖赏预期条件下的面孔快速提取信息, 导致早期的 P1成分主要受到奖赏预期的调节。
本研究利用情绪面孔作为目标刺激, 在枕颞位置记录到N170成分。N170是目标刺激出现后170 ms左右, 在后部外侧电极上记录到的负走向 ERP成分, 被认为是编码面孔结构的特异性成分(Itier& Taylor, 2004a, b, c)。本研究中N170波幅也未受到情绪的影响, 不同情绪面孔诱发的 N170无显著差异。可见, 早期的面孔结构编码加工没有受到情绪的影响, 这与之前的一些研究结果类似(Eimer &Holmes, 2002; Eimer, Holmes, & McGlone, 2003)。同时, 我们观察到奖赏预期对 N170波幅的影响,奖赏预期条件下目标面孔诱发了更正的 N170。在奖赏预期条件下, 面孔刺激得到了更有效的编码加工(Adcock, Thangavel, Whitfield-Gabrieli, Knutson,& Gabrieli, 2006)。而无奖赏预期条件下出现更负的N170可能是由于结构编码的难度增加, 被试为了获得奖赏, 而抑制了与奖赏无关的面孔的注意加工(Marini, Marzi, & Viggiano, 2011)。此外, 之前有研究指出, 相对于左半球, 右半球的 N170波幅更大,出现得更早(Bentin et al., 1996)。本研究并没有发现右侧头皮位置上N170波幅的优势。相反地, 与P1类似, 在左侧头皮位置出现了显著的奖赏效应。Shigemune 等人(2010)采用PET技术考察了高低奖赏对负性或中性图片再认成绩的影响, 发现奖赏的主效应激活了左侧的眶额皮层, 也支持左侧皮层对奖赏的优势化加工。然而, 关于大脑加工奖赏的单侧化优势机制还有待于进一步的深入研究。
尽管有研究认为, VPP与N170具有相同的皮层发生源, 是 N170在额中央区的极性反转(Joyce& Rossion, 2005), 但本研究中VPP与N170并没有得到汇聚性的结果。这与之前的一些研究结果类似,例如, Rossignol等人(2005)年的研究中采用oddball范式考察高、低焦虑组被试对情绪面孔的加工, 发现N100、P100和VPP均没有受到情绪以及焦虑水平的影响, 而N170显示出恐惧与高兴面孔的差异。研究者并没有对N170与VPP结果模式的差异进行解释。在本研究中, 与 N170的结果模式不完全相同, VPP既没有受面孔的情绪信息影响, 也没有受到奖赏预期的调节。这些不一致的结果可能是由于VPP和N170具有相互独立的皮层发生源(Bentin et al., 1996; Eimer, 2000)。此外, 本研究采用双侧乳突平均做离线参考, 可能将 VPP作为观测指标更合适, 若采用全脑平均参考可能将 N170作为观测指标更合适(Joyce & Rossion, 2005)。
在本研究中, 早期的面孔加工阶段(80~130 ms,130~200 ms)并没有出现情绪效应, 面孔情绪效应出现得较晚(200~400 ms), 中性面孔比情绪面孔诱发了更负的 N300, 说明个体在晚期对面孔情绪刺激的认知加工存在差异。然而, 过去研究结果中的N300在情绪性条件下(正性或负性)普遍比中性条件下更负, 研究者认为这体现了对情绪性刺激的精细加工(Carretié & Iglesias, 1995; Schutter, de Haan,& van Honk, 2004; Rossignol et al., 2005), 这与本研究结果不一致。同时, 在本研究中, 与N170趋势相同, 无奖赏预期条件下的 N300更负, 这与相关研究结果是一致的(Marini et al., 2011)。我们推测,当前研究中N300在情绪性面孔和中性面孔的差异与过去研究相反可能主要是受到了奖赏预期的影响。过去这些发现负性情绪诱发更负的N300的研究(Carretié & Iglesias, 1995; Schutter et al., 2004;Rossignol et al., 2005)均没有控制奖赏预期, 被试只是对目标面孔进行情绪辨别, 负性情绪易于吸引注意, 可能得到了更深入和更精细的加工。在当前研究中, 被试为获得奖赏, 加强了对反应速度和准确率的要求, 而中性面孔的显著性更低, 被试可能采用某种策略排除正性和负性面孔后才确认该面孔为中性面孔, 这可能造成了对中性面孔的反应时最长和引起的N300更负。今后研究应进一步考察情绪性信息引发的ERPs波幅如何受到奖赏预期的调节。
此外, 当前研究还发现 N300受到奖赏预期与情绪的交互影响。正性情绪加工效应并没有受到奖赏预期的影响, 不同奖赏预期条件下的正性情绪加工效应无显著差异; 而负性情绪加工效应以及负性偏向效应均受到奖赏预期的调节, 奖赏预期条件下的负性情绪加工效应和负性偏向效应均显著大于无奖赏预期条件下的负性情绪加工效应和负性偏向效应。可见, 对情绪刺激的加工过程受到动机的调节, 且不同情绪刺激加工效应受到奖赏预期的差异性影响, 这可能暗示奖赏与情绪存在互相影响的加工机制(Murray, 2007; Wittmann et al., 2008)。大量的研究指出, 动机显著性刺激能够促进注意的调节 (“fine-tuning”),使得注意偏向显著的刺激(Engelmann, Damaraju, Padmala, & Pessoa, 2009;Engelmann & Pessoa, 2007; Pessoa, 2009)。从这个方面来说, 负性面孔(如愤怒)传递出威胁的信号, 相比其他面孔更显著(salient)、能够更快地得到加工。人脑对负性刺激反应敏感, 而且这种负性偏向具有高度的一致性、稳定性(Delplanque, Silvert, Hot, &Segueira, 2005; Huang & Luo, 2006; Yuan et al.,2007; Yuan et al., 2012; 袁加锦, 李红, 2012)。奖赏期待调动注意资源使得被试对目标刺激更警觉, 负性刺激能够得到优先的注意, 使得奖赏预期条件下的负性情绪加工效应和负性偏向效应显著大于无奖赏预期条件下的效应。
然而, 最近一项 ERP研究(Kaltwasser et al.,2013)也采用线索-目标范式, 考察了不同奖赏预期条件下(奖赏、中性、惩罚)对目标词汇属性判断的(具体的或抽象的)ERP效应, 目标词可能是正性的、中性的或负性的。ERP结果没有发现目标词情绪效价与奖赏预期的交互作用, 作者认为奖赏与情绪加工依赖不同的神经机制。但是, 该研究与当前研究的显著不同是, 在当前研究中, 情绪效价是任务相关的, 而在 Kaltwasser等人(2013)的研究中,目标词的情绪效价是任务无关的。正因为奖赏预期能够促进注意集中在任务相关的刺激或刺激维度上, 当情绪效价与任务无关时, 则不会与奖赏发生交互作用, 进而不会影响被试的行为或神经反应。奖赏与情绪在神经机制上是否存在相互影响可能显著地受到任务情境的调节。
5 结论
本研究利用 ERP技术考察了动机性因素对情绪面孔加工的影响。首先, 奖赏预期增强了大脑的神经反应, 奖赏线索比无奖赏线索诱发了更正的P1、P2、P300, 这反映了动机性信息促进被试调动更多的注意资源, 准备对后续目标进行有效的加工。其次, 奖赏与情绪加工存在共同的神经机制。奖赏预期对不同面孔情绪加工效应(正性情绪加工效应、负性情绪加工效应、负性偏向效应)的差异性影响揭示了动机性因素对情绪加工的调节作用,动机性因素促进了个体在加工情绪面孔时的负性情绪加工效应以及负性偏向。
Adcock, R. A., Thangavel, A., Whitfield-Gabrieli, S., Knutson,B., & Gabrieli, J. D. E. (2006). Reward-motivated learning:Mesolimbic activation precedes memory formation.Neuron,50
, 507–517.Adolphs, R. (2002). Recognizing emotion from facial expression: Psychological and neurological mechanisms.Behavioral and Cognitive Neuroscience Reviews, 1
, 21–60.Adolphs, R., Jansari, A., & Tranel, D. (2001). Hemispheric perception of emotional valence from facial expressions.Neuropsychology, 15
, 516–524.Alheid, G. F. (2003). Extended amygdala and basal forebrain.Annals of the New York Academy of Sciences, 985
(1),185–205.Amaral, D. G., & Price, J. L. (1984). Amygdala-cortical projections in the monkey (macaca fasicularis).Journal of Comparative Neurology, 230
, 465–496.Aron, A., Fisher, H., Mashek, D. J., Strong, G., Li, H., &Brown, L. L. (2005). Reward, motivation, and emotion systems associated with early-stage intense romantic love.Journal of Neurophysiology, 94
, 327–337.Ashley, V., Vuilleumier, P., & Swick, D. (2004). Time course and specificity of event-related potentials to emotional expressions.Neuroreport, 15
, 211–216.Batty, M., & Taylor, M. J. (2003). Early processing of the six basic facial emotional expressions.Cognitive Brain Research, 17
, 613–620.Baxter, M. G., & Murray, E. A. (2002). The amygdala and reward.Nature Reviews Neuroscience, 3
, 563–573.Bentin, S., Allison, T., Puce, A., Perez, E., & McCarthy, G.(1996). Electrophysiological studies of face perception in humans.Journal of Cognitive Neuroscience, 8
, 551–565.Berridge, K. C., & Robinson, T. E. (2003). Parsing reward.Trends in Neurosciences, 26
, 507–513.Bijleveld, E., Custers, R., & Aarts, H. (2010). Unconscious reward cues increase invested effort, but do not change speed-accuracy tradeoffs.Cognition, 115
(2), 330–335.Blau, V. C., Maurer, U., Tottenham, N., & McCandliss, B. D.(2007). The face-specific N170 component is modulated by emotional facial expression.Behavioral and Brain Functions, 3
, 1–13.Botzel, K., & Grusser, O. J. (1989). Electric brain potentials evoked by pictures of faces and non-faces: A search for“face-specific” EEG-potentials.Experimental Brain Research, 77
, 349–360.Carretié, L., & Iglesias, J. (1995). An ERP study on the specificity of facial expression processing.International Journal of Psychophysiology, 19
, 183–192.Carretié, L., Iglesias, J., García, T., & Ballesteros, M. (1997).N300, P300 and the emotional processing of visual stimuli.Electroencephalography & Clinical Neurophysiology, 103
,298–303.Chiew, K. S., & Braver, T. S. (2011). Positive affect versus reward: Emotional and motivational influences on cognitive control.Frontiers in Psychology, 2
, 1–10.Costa, V. D., Lang, P. J., Sabatinelli, D., Versace, F., & Bradley,M. M. (2010). Emotional imagery: Assessing pleasure and arousal in the brain's reward circuitry.Human Brain Mapping, 31
(9), 1446–1457.Darwin, C. (Ed.). (1998).The expression of the emotions in man and animal
. New York: Oxford Press.Delplanque, S., Silvert, L., Hot, P., & Sequeira, H. (2005).Event-related P3a and P3b in response to unpredictable emotional stimuli.Biological Psychology, 68
(2), 107–120.Dimberg, U., & Öhman, A. (1996). Behold the wrath:Psychophysiological responses to facial stimuli.Motivation and Emotion, 20
, 149–182.Eastwood, J. D., Smilek, D., & Merikle, P. M. (2003).Negative facial expression captures attention and disrupts performance.Perception & Psychophsics, 65
, 352–358.Eimer, M. (2000). Attentional modulations of event-related brain potentials sensitive to faces.Cognitive Neuropsychology, 17
, 103–116.Eimer, M., & Holmes, A. (2002). An ERP study on the time course of emotional face processing.Neuroreport, 13
(4),427–431.Eimer, M., Holmes, A., & McGlone, F. P. (2003). The role of spatial attention in the processing of facial expression: An ERP study of rapid brain responses to six basic emotions.Cognitive, Affective, & Behavioral Neuroscience, 3
(2),97–110.Engelmann, J. B., & Pessoa, L. (2007). Motivation sharpens exogenous spatial attention.Emotion, 7
(3), 668–674.Engelmann, J. B., Damaraju, E., Padmala, S., & Pessoa, L.(2009). Combined effects of attention and motivation on visual task performance: Transient and sustained motivational effects.Frontiers in Human Neuroscience,3
(4), 1–15.Fox, E., Russo, R., & Dutton, K. (2002). Attentional bias for threat: Evidence for delayed disengagement from emotional faces.Cognition & Emotion, 16
(3), 355–379.Hansen, C. H., & Hansen, R. D. (1988). Finding the face in the crowd: An anger superiority effect. Journal of Personality& Social Psychology, 54
, 917–924.Herrmann, M. J., Ehlis, A. C., Ellgring, H., & Fallgatter, A. J.(2005). Early strages (P100) of face perception in humans as measured with event-related potentials (ERPs).Journal of Neural Transmission, 112
(8), 1073–1081.Hickey, C., Chelazzi, L., & Theeuwes, J. (2010). Reward changes salience in human vision via the anterior cingulate.Journal of Neuroscience, 30
, 11096–11103.Huang, Y. X., & Luo, Y. J. (2006). Temporal course of emotional negativity bias: An ERP study.Neuroscience Letters, 398
(1), 91–96.Itier, R. J., & Taylor, M. J. (2004a). Effects of repetition learning on upright, inverted and contrast-reversed face processing using ERPs.NeuroImage, 21
, 1518–1532.Itier, R. J., & Taylor, M. J. (2004b). N170 or N1?Spatiotemporal differences between object and face processing using ERPs.Cerebral Cortex, 14
(2), 132–142.Itier, R. J., & Taylor, M. J. (2004c). Source analysis of the N170 to faces and objects.Neuroreport: For Rapid Communication of Neuroscience Research, 15
(8),1261–1265.Ito, T. A., Larsen, J. T., Smith, N. K., & Cacioppo, J. T. (1998).Negative information weighs more heavily on the brain:The negativity bias in evaluative categorizations.Journal of Personality and Social Psychology, 75
(4), 887–900.Joyce, C., & Rossion, B. (2005). The face-sensitive N170 and VPP components manifest the same brain processes: The effect of reference electrode site.Clinical Neurophysiology,116
, 2613–2631.Kaltwasser, L., Ries, S., Sommer, W., Knight, R. T., &Willems, R. M. (2013). Independence of valence and reward in emotional word processing: Electrophysiological evidence.Frontiers in Psychology, 4
, 1–15.Kiss, M., Driver, J., & Eimer, M. (2009). Reward priority of visual target singletons modulates Event-Related Potential signatures of attentional selection.Psychological Science,20
, 245–251.Locke, H. S., & Braver, T. S. (2008). Motivational influences on cognitive control: Behavior, brain activation, and individual differences.Cognitive, Affective, Behavioral Neuroscience, 8
(1), 99–112.Luo, W. B., Feng, W. F., He, W. Q., Wang, N., & Luo, Y. J.(2010). Three stages of facial expression processing: ERP study with rapid serial visual presentation.Neuroimage, 49
,1857–1867.Marini, F., Marzi, T., & Viggiano, M. P. (2011).“Wanted!”The effects of reward on face recognition electrophysiological correlates.Cognitive, Affective, and Behavioral Neuroscience, 11
, 627–643.Mohanty, A., Gitelman, D. R., Small, D. M., & Mesulam, M.M. (2008). The spatial attention network interacts with limbic and monoaminergic systems to modulate motivation-induced attention shifts.Cerebral Cortex,18
(11), 2604–2613.Morris, J. S., Öhman, A., & Dolan, R. J. (1998). Conscious and unconscious emotional learning in the human amygdala.Nature, 393
, 467–470.Murray, E. A. (2007). The amygdala, reward and emotion.Trends in Cognitive sciences, 11
(11), 489–497.Onoda, K., Okamoto, Y., Shishida, K., Hashizume, A., Ueda,K., Kinoshita, A., Yamashita, H., & Yamawaki, S. (2006).Anticipation of affective image modulates visual evoked magnetic fields (VEF).Experimental Brain Research, 175
,536–543.O'Doherty, J., Kringelbach, M. L., Rolls, E. T., Homak, J., &Andrews, C. (2001). Abstract reward and punishment representations in the human orbitofrontal cortex.Nature Neuroscience, 4
(1), 95–102.Öhman, A. (1993). Fear and anxiety as emotional phenomenon:Clinical phenomenology, evolutionary perspectives and information processing mechanisms. In M. Lewis, & J. M.Haviland (Eds.),Handbook of emotions
(pp. 511–536).New York: Guildford Press.Öhman, A., & Mineka, S. (2001). Fears, phobias, and preparedness: Toward an evolved module of fear and fear learning.Psychological Review, 108
, 483–522.Öhman, A., Lundqvist, D., & Esteves, F. (2001). The face in the crowd revisited: a threat advantage with schematic stimuli.Journal ofPersonality and Social Psychology, 80
,381–396.Pessoa, L. (2009). How do emotion and motivation direct executive control?Trends in Cognitive Sciences, 13
(4),160–166.Pessoa, L., & Engelmann, J. B. (2010). Embedding reward signals into perception and cognition.Frontiers in Neuroscience, 4
, 1–8.Pinkham, A. E., Griffin, M., Baron, R., Sasson, N. J., & Gur, R.C. (2010). The face in the crowd effect: Anger superiority when using real faces and multiple identities.Emotion, 10
,141–146.Potts, G. F. (2004). An ERP index of task relevance evaluation of visual stimuli.Brain and Cognition, 56
, 5–13.Potts, G. F., & Tucker, D. M. (2001). Frontal evaluation and posterior representation in target detection.Brain Research.Cognitive Brain Research, 11
, 147–156.Rossignol, M., Philippot, P., Douilliez, M., Crommelinck, M.,& Campanella, S. (2005). The perception of fearful and happy facial expression is modulated by anxiety: An event-related potential study.Neuroscience Letters, 377
,115–120.Schultz, W., Dayan, P., & Montague, P. R. (1997). A neural substrate of prediction and reward.Science, 275
, 1593–1599.Schutter, D. J. L. G., de Haan, E. H. F., & van Honk, J. (2004).Functionally dissociated aspects in anterior and posterior electrocortical processing of facial threat.International Journal of Psychophysiology, 53
, 29–36.Shigemune, Y., Abe, N., Suzuki, M., Ueno, A., Mori, E.,Tashiro, M., Itoh, M., & Fujii, T. (2010). Effects of emotion and reward motivation on neural correlates of episodic memory encoding: A PET study.Neuroscience Research,67
, 72–79.Small, D. M., Gitelman, D., Simmons, K., Bolise, S. M.,Parrish, T., & Mesulam, M. M. (2005). Monetary incentives enhance processing in brain regions mediating top-down control of attention.Cerebral Cortex, 15
, 1855–1865.Smith, N. K., Cacioppo, J. T., Larsen, J. T., & Chartrand, T. L.(2003). May I have your attention, please: Electrocortical responses to positive and negative stimuli.Neuropsychologia, 41
, 171–183.Veling, H., & Aarts, H. (2010). Cueing task goals and earning money: Relatively high monetary rewards reduce failures to act on goals in a Stroop task.Motivation and Emotion, 34
,184–190.Vuilleumier, P., & Pourtois, G. (2007). Distributed and interactive brain mechanisms during emotion face perception: Evidence from functional neuroimaging.Neuropsychology, 45
, 174–194.Wang, Y., & Luo, Y. J. (2005). Standardization and assessment of college students’ facial expression of emotion.Chinese Journal of Clinical Psychology, 13
(4), 396–398.[王妍, 罗跃嘉. (2005). 大学生面孔表情材料的标准化及其评定.中国临床心理学杂志, 13
(4), 396–398.]Wei, P., & Kang, G. L. (2012). The brain mechanisms of reward cue in triggering and modulating fronto-parietal attentional network in visual search.Advances in Psychological Science,20
(6), 798–804.[魏萍, 康冠兰. (2012). 奖赏性线索启动和调控视觉搜索额顶网络的神经机制.心理科学进展, 20
(6), 798–804.]Williams, L. M., Palmer, D., Liddell, B. J., Song, L., &Gordon, E. (2006). The ‘when’ and ‘where’ of perceiving signals of threat versus non-threat.Neuroimage, 31
,458–467.Wise, R. A. (2004). Dopamine, learning and motivation.Nature Reviews Neuroscience, 5
(6), 483–494.Wittmann, B. C., Schiltz, K., Boehler, C. N., & Düzel, E.(2008). Mesolimbic interaction of emotional valence and reward improves memory formation.Neuropsychologia,46
(4), 1000–1008.Yuan, J. J., & Li, H. (2012). The human susceptibility to the valence strength of emotional stimuli: Neural mechanisms.Advances in Psychological Science, 20
(1), 10–18.[袁加锦, 李红. (2012). 人类对情绪事件效价强度的易感性及神经机制.心理科学进展, 20
(1), 10–18.]Yuan, J. J., Meng, X., Yang, J. M., Yao, G. H., Hu, L., & Yuan,H. (2012). The valence strength of unpleasant emotion modulates brain processing of behavioral inhibitory control:Neural correlates.Biological Psychology,89
(1), 240–251.Yuan, J. J., Zhang, Q., Chen, A., Li, H., Wang, Q., Zhuang, Z.,& Jia, S. (2007). Are we sensitive to valence differences in emotionally negative stimuli? Electrophysiological evidence from an ERP study.Neuropsychologia, 45
(12),2764–2771.猜你喜欢
健康体检与管理(2022年4期)2022-05-13
中国药学药品知识仓库(2021年18期)2021-02-28
初中生学习指导·中考版(2020年2期)2020-09-10
小资CHIC!ELEGANCE(2019年30期)2019-09-12
初中生世界·八年级(2019年8期)2019-08-29
价值工程(2018年25期)2018-09-26
中国现代医生(2017年24期)2017-10-17
女友·家园(2017年7期)2017-07-19
米娜·女性大世界(2016年8期)2016-08-17
科技视界(2016年6期)2016-07-12
