
综合处理措施对解除青钱柳种子休眠的影响
2014-01-04 09:47尚旭岚方升佐徐锡增
中南林业科技大学学报 2014年1期
尚旭岚,方升佐,徐锡增
(南京林业大学 资源与环境学院,江苏 南京 210037)
综合处理措施对解除青钱柳种子休眠的影响
尚旭岚,方升佐,徐锡增
(南京林业大学 资源与环境学院,江苏 南京 210037)
研究了综合处理措施对青钱柳种壳结构和透性的影响,并对休眠解除过程的果皮浸提液进行了生物测定。结果表明:浓硫酸处理可在一定程度上破坏青钱柳种子木质坚硬的种壳结构;在酸蚀处理基础上再进行GA3浸泡和GA3拌沙层积处理,随着层积时间的延长,种壳表面出现孔洞且逐渐变大,种胚含水量和种子呼吸速率显著提高,果皮浸提液抑制作用显著降低,裂口萌动种子中可能具抑制作用的物质种类减少。说明综合层积处理能有效破坏青钱柳种子坚硬的种壳结构,改善种皮透性,促进果皮中抑制物质的降解,从而加速种子休眠的解除。
青钱柳;酸蚀处理;拌沙层积处理;种壳结构;透性;抑制物质
青钱柳Cyclocarya paliurus(Batal.)Iljinskaja,又名青钱李、摇钱树等,系胡桃科Juglandaceae青钱柳属植物,是我国特有的单种属植物。青钱柳广泛分布于江西、浙江、江苏、安徽、福建、湖北、湖南、四川和贵州等地海拔420~2 500 m的山区、溪谷或石灰岩山地[1]。青钱柳集药用、材用和观赏等多种价值于一身,是一种具有极高保护和开发利用价值的珍贵树种[1-2]。青钱柳种子具深休眠习性,在一般播种育苗条件下隔年才能萌发,严重影响了其产业化进程。目前对其休眠原因已有了较深入的研究[3-6],也有学者对解除其休眠的措施进行了探索,结果表明酸蚀后再进行GA3浸泡和GA3拌沙层积的综合处理措施能有效解除青钱柳种子的休眠[7-8],但对其作用机理还不完全清楚。本文中通过研究酸蚀后GA3浸泡和GA3拌沙层积的综合处理措施对青钱柳种皮结构和透性的影响,并对解除休眠的种子果皮浸提液进行了生物测定和GC-MS鉴定,旨在探讨青钱柳种子休眠解除的机制并为生产上采取相应的技术措施提供理论依据。
1 材料和方法
1.1 材料和试验设计
供试种子采自云南昆明植物园(25°10′N,102°44′E,海拔1 943 m)单株青钱柳(树高,14 m;胸径,26 cm),种子千粒重为85.25 g,饱满率为41%。
酸蚀处理:种子去翅后用98%浓硫酸进行酸蚀处理,浸泡2.5 、5.0 和7.5 h后搓去碳化层,用流水冲洗干净,阴干待用。以未经处理的完整干种子作为对照(干CK)。
层积处理:分自然层积处理和综合层积处理两种方案。自然层积处理是将青钱柳干种子清水浸种48 h后,清水拌沙后置于室外层积,种沙比约为1︰3。分别在层积1个月、2个月、3个月和4个月定时取样进行指标测定,以未经处理的完整干种子作为对照(记为0)。综合层积处理是将种子进行酸蚀5.5 h后用500 mg·kg-1GA3浸种48 h,再用400 mg·kg-1GA3拌沙,按种沙比1︰3混匀后置于室外层积。取样时间设置为层积2个月(记为层2月)、层积3个月(记为层3月)、种子萌动裂口(记为裂口)、胚根刚伸出种皮(记为胚根伸出)。以未经处理的完整干种子作为对照(干CK)。
1.2 测定方法
1.2.1 不同酸蚀时间种壳结构的观察
取2.5、5和7.5 h 3个酸蚀处理时间和干CK种子,在扫描电子显微镜下观察种子表面结构并进行记录和拍照。过程如下:取各处理种子10颗,在乙二胺中恒温浸泡24 h,软化种皮,使种子容易解剖,然后将种子置于4%戊二醛中固定,0.1 M PBS清洗,乙醇系列脱水(30%、50%、70%、90%、100%),转入乙酸异戊酯置换,CPD-2临界点干燥仪干燥、粘台、离子溅射仪喷金,PHILIPS SEM-505扫描电子显微镜观察下观察、拍照。
1.2.2 综合层积处理过程种壳结构的观察
取综合层积处理30 、45 和60 d的种子各10颗,在扫描电子显微镜下观察种子表面结构并进行记录和拍照,其方法同1.2.1。
1.2.3 种胚含水量和种子呼吸速率的测定
按试验设计在自然层积和综合层积各阶段取样进行种胚含水量和种子呼吸速率的测定。呼吸速率测定采用小篮子法[9]。每个处理各取种子100粒,3次重复。含水量测定采用高温烘干法,取样后分离出种胚,在130℃恒温条件下,烘至恒质量。
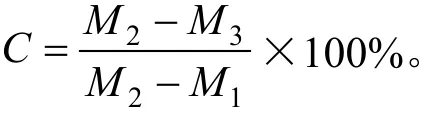
式中:C为含水量(%);M1为样品盒和盖的质量(g);M2为样品盒和盖及样品的烘前质量(g);M3为样品盒和盖及样品的烘后质量(g)。
1.2.4 层积过程中果皮甲醇浸提液的生物测定和萌动种子浸提液GC-MS鉴定
取综合层积处理不同时段的果皮干粉末10 g置于100 mL锥形瓶中,加入 4倍体积80%甲醇,混匀后放入冰箱,在1~4 ℃的条件下密闭浸提。其间多次取出摇动使其充分浸提,24 h后过滤,重复浸提2次后,混合3次所得的浸提液,减压浓缩后定容到1 g·mL-1。参考尚旭岚等[5]的方法对各取样时段的果皮浸提浓缩液稀释至0.2 g·mL-1进行白菜种子生物测定。另将裂口种子1 g·mL-1果皮浸提液送中国林科院林产化工研究所仪器分析室,参考尚旭岚等[5]的条件进行GC-MS鉴定。
1.3 数据分析
采用SPSS 13.0进行单因素方差分析,并用Duncan法检验其差异显著性。结果以平均数±标准偏差表示。
2 结果与分析
2.1 不同酸蚀时间对种壳结构的影响
青钱柳种子经酸蚀处理后,不仅种子变小,种壳变薄,而且其种壳表面出现了很多变化。不同酸蚀时间处理后种壳结构的电镜观察结果显示,未经处理的青钱柳种子种壳表面角质化(见图1A);浓硫酸处理2.5 h后搓表面变得坑坑凹凹,出现少量腐蚀坑(见图1B);酸蚀5 h和7.5 h后种壳表面的腐蚀坑越来越多,且越来越大(见图1C和1D)。
2.2 综合层积处理对种壳结构的影响
与干种子角质化表皮结构(见图2A)相比,综合层积处理30 d时种壳表面出现了小洞(见图2B),而且这些孔洞随层积时间的延长而变大(见图2C和图2D),这种特殊的结构变化有利于种子吸水和透气,从而加速种子休眠的解除。
2.3 不同层积处理对种子透性的影响
2.3.1 不同层积处理过程种胚含水量变化
水分是控制种子萌发的最重要因素,只有在细胞内自由水增多的条件下,种子的贮藏物质才能被动员,同时使酶活化或合成而起催化作用。

图1 不同酸蚀时间处理后种壳结构变化Fig. 1 Changes of ultra-structure of husk scarif i ed in sulphuric acid for different times

图2 综合处理措施层积过程种壳结构变化Fig. 2 Changes of husk treated by comprehensive treatments during stratif i cation
从图3可看出,随着层积时间的延长,综合层积处理的种胚含水量呈现不断上升的趋势,干种子种胚的含水量为4.97%,层积至刚裂口时含水量达到35.01%,比干种子增加了6倍。方差分析和多重比较结果表明,层积处理各阶段种胚的含水量差异达到极显著水平。对于自然层积种子来说,层积第1个月种胚含水量有一个较大的提高,但随着层积时间的延长其种胚的含水量变化不大,到层积4个月时含水量仍只有18.94%。多重比较结果也表明,层积4个月间的种胚含水量未达到极显著差异水平。
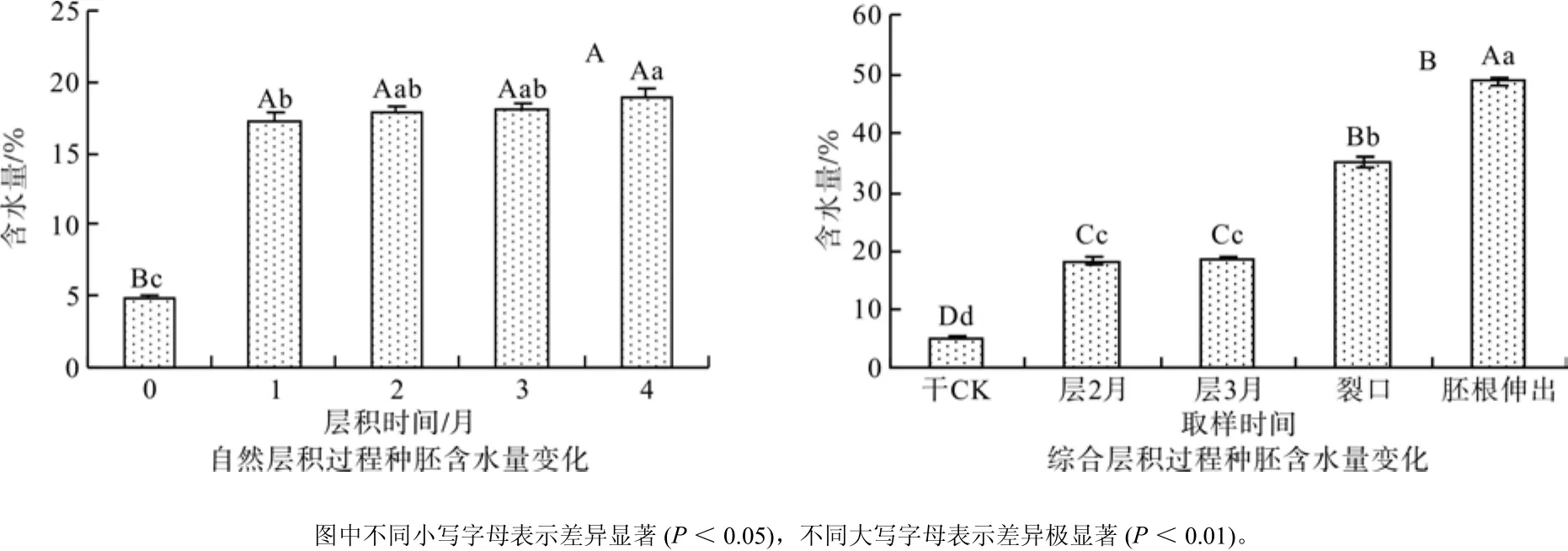
图3 不同层积处理过程种胚含水量变化Fig.3 Changes of water content in embryos under different treatments
2.3.2 不同层积处理过程种子呼吸速率变化
一般干燥种子的呼吸都被抑制在最低水平上,但随着种子的吸胀,呼吸作用会不断增强。通过对不同层积处理过程中呼吸速率的比较,来阐明青钱柳种子休眠解除过程的呼吸生理。
从图4中可知,随着综合层积处理时间的延长,青钱柳种子的呼吸速率不断提高,层积到第3个月时提高到28.1 mg·(100 g)-1,这说明综合层积处理可在一定程度上改善种皮的透气情况,至种子裂口萌动时,其呼吸速率达到干种子的9.96倍,到胚根伸出时进一步增加到干种子的15.21倍,达到94.17 mg·(100g)-1。方差分析和多重比较结果表明,综合层积处理各阶段的种胚呼吸速率差异达到极显著水平。随着自然层积时间的延长,青钱柳种子的呼吸速率有所提高,但整个层积过程的呼吸速率变化都较小。方差分析和多重比较结果也表明,自然层积4个月时的呼吸速率才与干种子达到极显著差异水平。说明自然层积过程种子内部的代谢活动很微弱,这可能就是青钱柳种子在自然条件下通常需要两冬一夏才能萌发的原因之一。
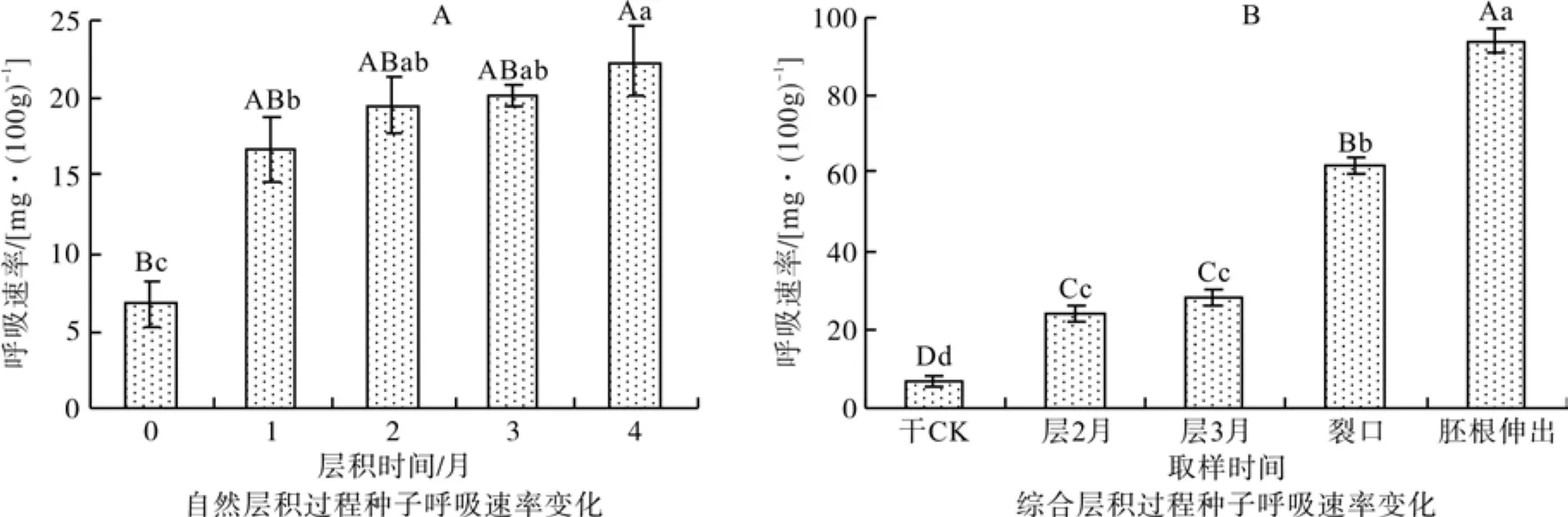
图4 不同层积处理种子呼吸速率变化Fig. 4 Changes of respiratory rate in seeds under different treatments
2.4 果皮甲醇浸提液对白菜苗高和根长的影响
综合层积处理种子休眠解除过程的种子果皮甲醇浸提液生物测定结果(见图5)表明,从干种子至裂口萌发,白菜种子的苗高和根长逐渐增加,说明果皮浸提液对白菜种子的抑制作用随着层积时间的延长逐渐降低。方差分析结果表明,不同层积时间果皮浸提液对苗高和根长的差异均达到极显著水平(苗高F=41.500,P<0.01;根长F=48.683,P<0.01)。多重比较结果显示,干种子和水浸12 d的苗高和根长与对照及其它层积时间之间均有极显著差异,说明干种子的抑制作用最强,而果皮中的抑制物质可能随休眠的解除而不断降解,因此到种子裂口萌动时抑制作用最弱。
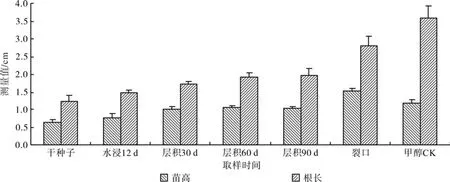
图5 休眠解除过程中果皮甲醇浸提液对白菜生长的影响Fig.5 Effects of extracts from pericarp during breaking dormancy on cabbage growth
2.5 裂口萌动种子果皮甲醇浸提液GC-MS鉴定
将已裂口萌动种子的果皮甲醇浸提液进行GC-MS鉴定,其总离子流程见图6。具有发芽抑制作用的有机化合物的种类及相对含量列于表1。
裂口萌动种子果皮甲醇浸提液中可能具抑制作用的有机化合物的种类和含量较干种子果皮明显减少[5],而且未发现抑制作用较强的酚酸类化合物及其衍生物,这可能正是萌动种子果皮浸提液抑制作用降低的原因。
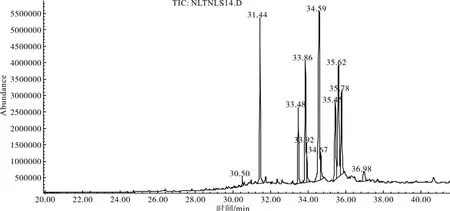
图6 萌动种子果皮甲醇浸提液GC-MS总离子流色谱Fig. 6 Total ion chromatogram of methanol extract from pericarp of germinating seeds

表1 萌动种子果皮甲醇浸提液的GC-MS分析结果Table 1 Results of GC-MS analysis for methanol extract from pericarp of germinating seeds
3 讨 论
解除种子休眠的方法很多[10-13],如化学药剂处理、激素处理、层积及一些物理方法,通常需要针对种子休眠的原因来寻找适宜的破眠方法。许多林木种子休眠的原因并不是单一的,因此需要采用综合的处理措施来解除其休眠。如深休眠的秤锤树种子在酸蚀2 d的基础上经500 mg·kg-1GA3溶液浸种预处理可获得40.1%的出苗率[14]。对大果冬青种子采用浓硫酸处理1.5 h后经250 mg·kg-1GA3和0.2 % KNO3混合溶液浸种24 h,再进行冷变温层积的综合处理措施可提高其发芽率[16]。王艳华等[12]研究表明,采用浓硫酸处理40 min+400 mg·kg-1GA3浸种+3~5℃低温层积30 d的综合处理措施可快速打破大山樱种子的休眠。前期研究结果表明,青钱柳种子经酸蚀后再进行GA3浸种和GA3拌沙室外变温层积能有效解除青钱柳种子休眠[8],但此综合处理措施的破眠机制尚不明晰。
董厚德等[17]对结缕草种子休眠解除前后的种皮结构进行扫描电镜观察,发现经处理的种子脂类物质及颖孔盖脱落,颖孔被打开,构成颖的纤维网络清晰可见,因此种子内外的水、气交换可顺利进行,减少胚生长的阻力,发芽率达到潜在生活力水平。Lin等[18]研究结果表明,热水浸泡等处理可促进苦瓜种子萌发和幼苗生长,通过对其种皮表面结构电镜扫描观察,发现采用热水浸泡等处理可使种皮细胞更大,细胞间隙也变大,透水透气性得改善,这是种子休眠解除的原因之一。黄丹等[19]对美国桂花种子的研究结果也表明,随着低温湿沙层积时间的增加,美国桂花种壳中的种孔直径有明显增大、数量不断增多的趋势。陈昕等[20]对层积处理过程中黄山花楸种子表皮和子叶细胞进行了透射电镜观察,结果表明黄山花楸种子萌发率和休眠解除效果与种皮角质层破裂解体、子叶细胞结构变化及储藏物质降解程度密切相关。本研究通过电镜扫描观察,结果表明酸蚀处理不仅可以有效破坏青钱柳种子表面的角质层,而且适当的腐蚀还可使种皮明显变薄,这样不仅可增加种皮的透性,又可减轻胚根突破种皮时的机械阻力。在酸蚀处理基础上再进行GA3浸种和GA3拌沙的综合层积处理,种皮表面出现了明显的小孔洞,而且这些孔洞随层积时间的延长而变大。从青钱柳种子层积各阶段种胚含水量和种子呼吸速率测定结果来看,自然层积处理的变化较小,而综合层积处理的种子随层积时间的延长至种子萌动均达到极显著差异水平,这与其种壳表面结构的变化是基本一致的,表明综合层积处理措施有效促进了种皮透性的改善。
Wróbel等[21]研究表明,休眠的葡萄种子中含有较高含量的儿茶酸和没食子酸,以及少量的香豆酸等酚酸类物质,层积后这些物质含量都大大降低。Soltani等[22]对东方山毛榉种子进行研究后发现,休眠解除过程酚类物质含量也呈现不断降低的趋势。本实验结果表明,青钱柳果皮浸提液的抑制作用随休眠的解除而不断减弱,与未经处理的种子相比[5],裂口萌动种子果皮甲醇浸提液中可能具发芽抑制作用的有机化合物的种类减少,这说明综合层积处理措施有效促进了青钱柳种子中香草酸等酚类物质的降解,从而使种子休眠较快得以解除。
综上所述,酸蚀处理可在一定程度上破坏青钱柳种子木质坚硬的种皮结构,减轻其机械障碍和透性障碍。在此基础上进行GA3浸种和拌沙综合层积处理,能够促进种皮透性的改善和果皮中抑制物质的降解,从而加速种子休眠的解除。因此,在生产上采用酸蚀处理结合GA3浸种和GA3拌沙的综合层积处理措施是快速有效的,但值得注意的是,在进行酸蚀处理时,应根据种子千粒重大小以及种皮厚度确定最佳的酸蚀时间,否则容易出现因酸蚀时间不当导致种子难萌发或种胚受伤的情况。
[1] 方升佐, 洑香香. 青钱柳资源培育与开发利用的研究进展[J].南京林业大学学报:自然科学版, 2007, 31(1): 95-100.
[2] 方升佐, 杨万霞. 青钱柳的开发利用与资源培育[J]. 林业科技开发, 2003, 17(1): 49-51.
[3] 杨万霞, 方升佐. 青钱柳种皮甲醇浸提液的生物测定[J]. 植物资源与环境学报, 2005, 14(4): 11-14.
[4] 王家源, 方升佐. 青钱柳种子层积过程中贮藏物质含量及酶活性的变化[J]. 南京林业大学学报: 自然科学版, 2007, 31(1):111-113.
[5] 尚旭岚, 徐锡增, 方升佐. 青钱柳种子休眠机制[J]. 林业科学,2011, 47(3): 68-74.
[6] 尚旭岚, 孙 容, 徐锡增, 等. 青钱柳种子不同部位发芽抑制物质的测定[J]. 林业科技开发, 2011, 25(5): 29-32.
[7] 史晓华, 徐本美, 黎念林, 等.青钱柳种子休眠与萌发的研究 [J]. 种子, 2002, 21(5): 5-7.
[8] Fang S Z, Wang J Y, Wei Z Y, et al. Methods to break seed dormancy in Cyclocarya paliurus (Batal) Iljinskaja [J]. Scientia Horticulturae, 2006, 110: 305-309.
[9] 黄学林主编. 种子生理实验[M]. 北京: 农业出版社, 1990:99-100.
[10] 杨晓玲,郭守华,张建文,等.层积和激素处理对山楂种子生理生化的影响[J].经济林研究,2009,27(1):76-79.
[11] 杨晓玲,郭守华,关学敏,等.山楂种子层积过程中抑制物质的变化[J].经济林研究,2009,27(2):44-48.
[12] 游双红,钟 诚,涂淑萍.变温层积过程中圆齿野鸦椿种子内含抑制物的生理活性变化[J].经济林研究,2013,31(3):41-47.
[13] 洑香香,周晓东,刘红娜.山茱萸种子休眠机理与解除方法初探[J].中南林业科技大学学报,2013,33(4):7-12.
[14] 徐本美, 冯桂强, 史 华, 等. 从秤锤树种子的萌发论酸蚀处理效应[J]. 种子, 1999, 119(5): 45-47.
[15] 徐本美, 史晓华, 孙运涛, 等. 大果冬青种子的休眠与萌发初探[J]. 种子 , 2002, 122(3): 1-2.
[16] 王艳华, 高述民, 李凤兰. 大山樱种子休眠机理的探讨[J]. 种子, 2005, 24(5): 12-16.
[17] 董厚德, 宫莉君. 结缕草种子(颖果)解除休眠前后的颖显微结构[J]. 草地学报, 2001, 9(4): 243-247.
[18] Lin J M, Sung J M. Pre-sowing treatments for improving emergence of bitter gourd seedlings under optimal and suboptimal temperatures [J]. Seed Sci. & Technol., 2001, 29: 39-50.
[19] 黄 丹,许岳香,胡海波. 美国桂花种子低温层积过程中种子结构的变化[J]. 中南林业科技大学学报, 2010,30(4):101-106.
[20] 陈 昕,曹珊珊,张红星.黄山花楸种子休眠解除技术及其解剖学观察[J]. 南京林业大学学报:自然科学版, 2011, 35(4):23-26.
[21] Wróbel M, Karama M, Amarowicz R, et al. Metabolism of phenolic compounds in Vitis riparia seeds during stratification and during germination under optimal and low temperature stress conditions [J]. Acta Physiologiae Plantarum, 2005, 27: 313-320.
[22] Soltani A. Phenolics in the seed coat of oriental beech (Fagus orientalis lipsky) as an autotoxic factor [C]//Proceedings of the Fourth International Iran and Russia Conference, 2004: 1048-1055.
Effects of comprehensive treatments on breaking dormancy of Cyclocarya paliurus seeds
SHANG Xu-lan, FANG Sheng-zuo, XU Xi-zeng
(College of Forest Resources and Environment, Nanjing Forestry University, Nanjing 210037, Jiangsu, China)
Cyclocarya paliurusseeds remain deep dormancy characteristics, and the seeds usually take two years to germinate in natural condition. The effects of comprehensive treatments to the structure and permeability of husk were studied, and the bioassays of the pericarp extract during breaking dormancy were measured. The results show that the chemical scarif i cation with concentrated sulphuric acid (H2SO4) could destroy the hard structure of husk to a certain extent, therefore could alleviate the mechanical and permeability barriers; after chemical scarif i cation, the seeds were soaked in gibberellic acid (GA3) and then stratif i ed in the sand moistened with GA3;while the stratif i cation time of comprehensive treatments was prolonged, some pores appeared in the surface of husk and gradually became larger; meanwhile, the water content on embryo and seed respiratory were signif i cantly enhanced, however the inhibitory effects of pericarp extract were signif i cantly reduced. The Gas Chromatography-Mass Spectrometry (GC-MS) identif i cation results for methanol extract from pericarp showed that there were fewer potential inhibitors in the germinating seeds than those in the untreated seeds. In conclusion, the comprehensive treatments combined with chemical scarif i cation, exogenous application of GA3and stratif i cation could destroy the hard structure of husk, promote the exchange of gas and water, and accelerate the degradation of inhibitors, therefore it could alleviate dormancy ofC. parliurusseeds in a relatively short period of time.
Cyclocarya paliurus; acid etching; mixing sand stratif i cation; seed husk structure; permeability; inhibitory substance
S718.43;Q945.35
A
1673-923X(2014)01-0042-07
2013-08-10
国家林业局林业科学技术推广项目﹙[2011]18 号);国家林业公益性行业科研专项(编号:200904046)
尚旭岚(1978-),女,四川乐山人,讲师,主要从事森林培育研究;E-mail:shangxulan@njfu.edu.cn
方升佐(1963-),男,安徽歙县人,教授,主要从事人工林定向培育研究;E-mail:fangsz@ njfu. edu. cn
[本文编校:谢荣秀]
猜你喜欢
安徽农业科学(2022年18期)2022-10-13
花卉(2020年24期)2021-01-04
防护林科技(2020年11期)2020-12-30
东北农业大学学报(2020年11期)2020-12-17
潍坊学院学报(2020年6期)2020-11-22
农业技术与装备(2020年7期)2020-08-18
中国食品学报(2019年3期)2019-01-13
天津农业科学(2018年3期)2018-03-24
江苏农业科学(2017年12期)2017-09-16
中成药(2017年6期)2017-06-13
