
三倍体毛白杨叶片水分特征及其对蒸腾的调节和影响
2014-01-03 09:06李广德贾黎明富丰珍
中南林业科技大学学报 2014年8期
李广德,贾黎明,富丰珍
(1. 国家开放大学 农林医药学院,北京 100039;2. 北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083;3.黑龙江省大庆市林业局,黑龙江 大庆 163002)
三倍体毛白杨叶片水分特征及其对蒸腾的调节和影响
李广德1,2,贾黎明2,富丰珍3
(1. 国家开放大学 农林医药学院,北京 100039;2. 北京林业大学 省部共建森林培育与保护教育部重点实验室,北京 100083;3.黑龙江省大庆市林业局,黑龙江 大庆 163002)
为探讨三倍体毛白杨叶片水分特征及其与蒸腾作用的相互关系,利用二氧化碳变量法、烘干法、压力平衡法等对三倍体毛白杨速生林叶片蒸腾速率、含水量、水势、水容等水分特征进行了连续2年的动态观测。结果表明,三倍体毛白叶片蒸腾速率(Tr)呈“单峰型”日变化,与之相反,其叶片绝对含水量(CLAW)、相对含水量(CLRW)、水势(Ψ)、水容(C)等呈“V”型。Tr与CLAW、CLRW、Ψ、C等具有一定的相关性,但不同时段其相关性及相关程度不同。由于树体水容充、放水特性,CLAW、CLRW、Ψ、C与Tr存在不同程度的滞后效应,与环境因素共同调节和影响树木蒸腾。
三倍体毛白杨;蒸腾;水势;水容
水作为重要的生态因子,对植物形态建成、生长发育、植被分布、群落演替等起着重要的作用[1],森林水循环作为全球物质循环中的重要环节,共同影响大气、土壤和植被的结构、功能、分布格局及其动态变化,在维持生物圈和地圈的生态平衡过程中起着重要作用[2]。森林作为功能强大的生态系统,在发挥其巨大水文效益的同时,也要消耗大量水分以维持自身生态系统的正常运转,存在着许多诸如水分输入输出矛盾冲突等问题。
植物蒸腾作用失水造成的水势梯度是植物水分吸收和运输的主要驱动力,蒸腾作用引起的水分上升也有助于根部吸收的无机离子以及根系中合成的有机物运转到植物体内以满足其生命活动的需要,同时,蒸腾作用可以调节叶片温度,避免高温灼伤。蒸腾速率是植物水分状况最重要的生理指标之一,对其日变化和季变化的研究有助于了解植物的蒸腾及水分利用特性[3]。树体组织含水量是反映树体水分状况的重要指标[4],绝对含水量是植物本身水分含量的反映,相对含水量是相对于饱和吸水时的水分状态,可以反映植物组织对水分的吸收能力。植物叶水势是表示植物水分状况或水分亏缺程度的一个直接指标,代表植物水分运动的能量水平[5]。水容是指单位水势变化引起植物组织含水量的变化,能反映植物组织和器官的含水量和水势变化关系,是导致叶水势与蒸腾速率存在滞后现象的主要原因[6],研究不同植物的水容特性与其蒸腾耗水之间的相互关系,有助于理解SPAC系统的水分运移机制[7]。
本研究以速生树种三倍体毛白杨为对象,对其两个生长季的蒸腾速率、叶片含水量、水势和水容进行了测定和分析,旨在揭示三倍体毛白杨叶片蒸腾速率及水分特征的日、季变化规律及其相互关系,为进一步研究和理解林木蒸腾速率影响和调控机制,以及田间水分管理提供一定依据。
1 研究区域概况
试验地位于山东省高唐县梁村镇盖洼村(36°58´N,116°14´E),海拔平均 27 m。暖温带半干旱季风区域大陆性气候,具有显著的季节变化和季风气候特性,光照充足,热量丰富。春季,降水少,风大,气候干燥,降水量占全年降水量的13.7%;夏季,温度高,湿度大,降水多,降水量占全年降水量的66.5%;秋季,气温急降,天气凉爽,降水量少,降水量占全年降水量的17.1%;冬季,低温寒冷,雨雪稀少,降水量只占全年降水量的2.7%。年降水量306.0~975.9 mm,年平均蒸发量1 880 mm,年平均温度12.0~14.1 ℃,极端最高气温达41.2 ℃,极端最低气温达-20.8 ℃。年日照总时数4433.5 h,无霜期204 d。
试验林地为潮土,土壤容重平均1.51 g·cm-3,平均有机质含量0.39%,全氮0.131%,全P 0.129%,K 0.612%,速效氮 14.486 mg·kg-1,速效P 1.752 mg·kg-1,速效 K 49.914 mg·kg-1。
2 材料与方法
2.1 试验材料
试验材料为三倍体毛白杨(hybrid triploids clones of Chinese White Popular (Populus tomentosaCarr.×Populus bolleanaLouche)×Populus tomentosaCarr.)。2005年春采用1年生根萌苗造林,带状配置,行向南北,密度1 m×2 m+6 m,试验林约3.4 hm2,2009年生长季末林分平均胸径8.90 cm,平均树高11.90 m。
2.2 蒸腾速率测定
选择试验林地中心位置,生长发育良好、无明显病虫害的三倍体毛白杨3株作为样树,使用便携式光合测定系统Licor-6400测定叶片蒸腾速率(Tr, mmolH2O·m-2·s-1)。测定时,将 Licor-6400放置于向阳处,每株样树剪取中上部枝条2根,一根用于测定蒸腾速率,另一根用于测定叶片水分特征,3次重复,自然光源。试验于2008和2009年生长季典型晴天进行(2008年测定日为5月26日,7月12日,8月24日,10月24日;2009年测定日为5月17日,6月30日,8月27日,9月26日,10月23日),日周期从6:00开始,18:00结束,时间步长为2 h。
2.3 叶片含水量、水势测定
与叶片蒸腾速率同步,将用于测定叶片水分特征的枝条下部放入盛有少量水的塑料桶中,迅速带回室内,“压力室法”(SKPM1400, UK)测水势(ψ, MPa),并扫描叶面积(S, cm2)及体积(V,cm3)(Color Scanner Expression 1680,Canada);另选3片叶子,编号,称重(Wr,g)后置于盛有蒸馏水的容器中在黑暗中浸泡24 h,使其充分吸水,称饱和重(Ws,g)并烘干至恒重(Wd,g)。
2008年试验期间分析叶片含水量与水势数据关系时发现,同海州常山Clerodendrum trichotomumThumb.[8]、 树 锦 鸡 儿Caragana arborescensLam.、 柠 条 锦 鸡 儿Caragana korshinskiiKom.和小叶锦鸡儿Caragana microphyllLam.[9]等树种,三倍体毛白杨叶相对含水量与水势呈极显著正相关(P<0.01),相关关系拟合曲线的决定系数都在0.85以上。由于“压力室法”测定叶片水势费时费力,有时一片叶片测定时间甚至达到或超过10 min,这样就很难保证叶片本身的水分状态不受干扰。因此,2009年叶片水势由2008年的拟合曲线(图1)通过测量相对含水量计算而来。
2.4 叶片含水量、水容计算
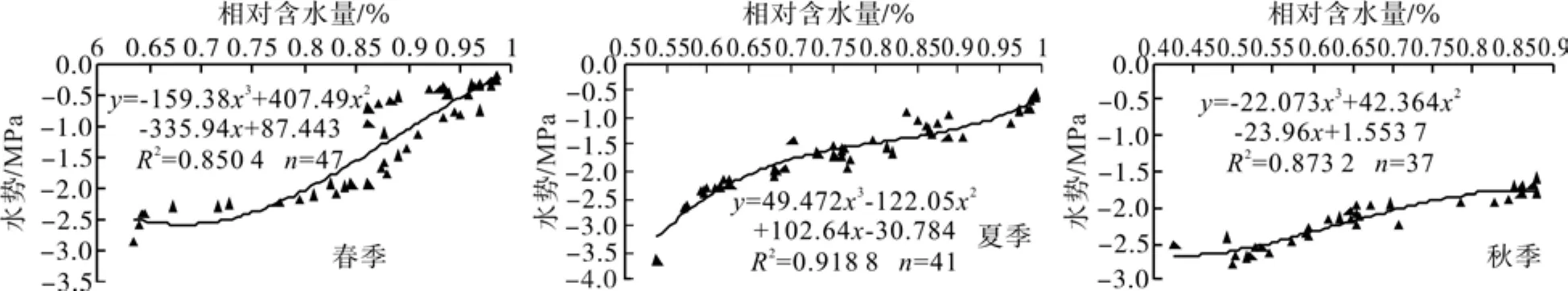
图1 三倍体毛白杨叶片水势与相对含水量相关关系曲线Fig. 1 Correlation between leaf relative water content and water potential in different seasons of triploid clones P. tomentosa
叶片绝对含水量(占鲜重)(Absoluteness Water Content,CLAW, %)、相对含水量(Relative Water content,CLRW, %)、 水 容(Leaf specif i c Water Capacitance,C, g·cm-3MPa-1)分别采用以下公式计算:
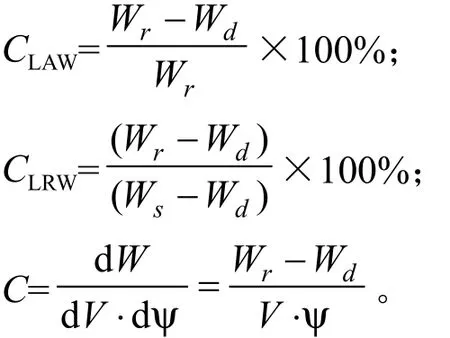
3 结果与分析
3.1 三倍体毛白杨叶片蒸腾速率日、季变化
三倍体毛白杨叶片蒸腾速率呈“单峰型”日变化(图2,a),春季和秋季于12:00~14:00达到峰值,夏季峰值出现在10:00左右。从峰值来看,2008年春季最大,为(7.83±0.55)mmol·m-2s-1;2009年夏末秋初最大,为(9.78±1.33)mmol·m-2s-1。

图 2 三倍体毛白杨速生林叶片蒸腾速率日、年变化特征Fig. 2 Diurnal and average daily Tr variation of triploid clones P. tomentosa in 2008 and 2009
从蒸腾速率日平均值来看(图2,b),不同年份随季节变化不同:2008年随季节变化呈逐渐减小趋势,2009年呈单峰型。2008整个生长季三倍体毛白杨叶片蒸腾速率日平均值为(3.90±0.49)mmol·m-2s-1,2009 年为(3.89±0.36)mmol·m-2s-1,两年蒸腾速率没有差异,但2009年的标准差更小,从一个侧面反映出2009年(造林第5年)的林分比2008年(造林第4年)更趋于稳定。
3.2 三倍体毛白杨叶片水分特征
3.2.1 三倍体毛白杨叶片含水量、水势、水容日变化
与蒸腾速率日变化相反,三倍体毛白杨叶片绝对含水量、相对含水量、水势、水容等随时间呈“V”型日变化(图3)。日出后,太阳辐射逐渐增强,气温不断升高,空气相对湿度下降,叶内外水汽压梯度增大,叶片蒸腾逐渐增大,绝对含水量随着失水的加快而逐渐减小,水分逐渐亏缺,导致叶细胞相对体积减小,相对含水量减小;为满足不断增加的蒸腾耗水的需求,叶片水势下降,以增强从土壤中吸收水分的能力,此时根系吸水速率小于蒸腾速率,树体组织由于水容作用持续放水,随着根系吸水速率和蒸腾速率差值的不断增大,水容逐渐降低。高辐射、高温、低湿的中午12:00~14:00随着蒸腾速率达到极值,叶片失水速率达到最大而导致CLAW、CLRW、ψ、C降低到一天中的最低值,叶片水分亏缺最为严重,叶片对自身水分耗散的调节和控制能力最弱,但吸水潜力最大。波谷出现之后,随着蒸腾作用的减缓,叶片失水减小,叶片水分条件逐渐改善,含水量、水势、水容开始回升。入夜后,三倍体毛白杨持续水分吸收和储存状态,叶片水分延续上升趋势,直到次日00:00~03:00或早上叶片蒸腾消耗大于其从土壤中吸收的水分时,叶片开始失水。

图 3 三倍体毛白杨速生林叶片水分特征日变化Fig. 3 Diurnal variation of triploid clones P. tomentosa leaf CLAW, CLRW, Ψ, and C in 2008 and 2009
3.2.2 三倍体毛白杨叶片含水量、水势、水容季节变化
三倍体毛白杨叶片绝对含水量随季节变化呈现“低-高-低”(图4)的变化趋势;相对含水量、水势、水容从春季到夏季到秋季逐渐递减,但春季和夏季差别不大,而从夏季到秋季则有个显著降低的过程,表明在春季和夏季三倍体毛白杨组织水分储存和释放能力较强,对树木蒸腾耗水的调节能力也较强。夏秋之交叶片相对含水量、水势、水容的突然降低可能是其生长变缓的一个具体表现,这也正好和试验地三倍体毛白杨部分叶片提前脱落(8月落叶)的现象相吻合,它们之间是否存在必然的联系,是否存在必然的因果关系,还有待进一步研究和考证。

图 4 三倍体毛白杨速生林叶片水分特征年变化Fig. 4 Average daily values of triploid clones P. tomentosa leaf CLAW, CLRW, Ψ, and C in 2008 and 2009
2008和2009生长季三倍体毛白杨叶片CLAW、CLRW、ψ、C分 别 为(61.88%±2.00%)和(67.41%±1.41%)、(74.23%±3.42%) 和(73.58%±4.73%)、(-2.00±0.23)和(-1.71±0.25)MPa、(4.48±0.73)×10-3和(5.19±1.22)×10-3g·cm-3MPa-1。
3.3 三倍体毛白杨叶片蒸腾速率与水分特征的相关性
表1为2008年、2009年及两年综合的三倍体毛白杨叶片蒸腾速率与水分特征的相关性分析表。结合图2~3可以发现,三倍体毛白杨叶片蒸腾速率日变化与叶片CLAW、CLRW、ψ、C相反,呈现出一定的相关性。
三倍体毛白杨叶片蒸腾速率与CLAW、CLRW、ψ、C在2008年和2009年呈现一定的负相关,而从2年综合数据来看,Tr除与CLAW呈负相关外,与CLRW、ψ、C都呈现正相关,但只有C达到显著。造成这种相关性相反的原因可能与数据分析有关,2008年与2009年各指标的相关性系数是分别从这两年各测点得到的数据进行相关分析得出的,而综合指标的相关性系数则是通过2年测定日平均值的相关性分析得到的。表明在日变化尺度上,叶片蒸腾速率与CLAW、CLRW、ψ、C等呈负相关,而在大的时间尺度如年变化上,叶片蒸腾速率则与叶片水分条件呈正相关,随着叶片水分条件的变化而相应变化。说明在叶片蒸腾速率的日变化过程中,叶片水势、含水量、水容等并不是引起叶片蒸腾速率增加或降低的原因,而是结果。但是从较大的尺度上来看,叶片蒸腾速率的大小则在一定程度上受控于叶片的水分状态,水分状态好有利于叶片蒸腾,水分条件较差则可限制叶片的蒸腾作用。这也体现了植物对环境条件的反馈调节作用和能力,当环境条件较为优越时,植物体各项生理功能便会得到最大的发挥,而当环境条件比较恶劣时,植物体便会通过各种途径来减少蒸腾对水分的损失,保持自身良好的水分平衡。

表1 三倍体毛白杨叶片蒸腾速率与水分特征相关性分析表Table 1 Correlations between Tr and CLAW, CLRW, Ψ, and C of triploid clones P. tomentosa
虽然相关程度不同,三倍体毛白杨叶片CLAW、CLRW、ψ、C之间都存在一定的正相关关系,表明植物体随外界环境条件变化的协同性和整体性,即随着环境条件和自身生理条件的变化,三倍体毛白杨叶片水分含量、水势、水容呈现出协调一致的变化趋势和方向,以此来适应环境条件的变化。
4 讨 论
植物蒸腾作用是一个复杂的生理和物理过程,受树种自身及环境等多种因素影响和控制[10-11]。叶片水势不仅是反映植物缺水最灵敏的指标,而且可以反映SPAC系统水流动力。与大多数植物一样,三倍体毛白杨蒸腾速率呈“单峰型”日变化[11-13],叶水势呈“V”型[5,11],日出后随蒸腾的加强而下降,正午前后达到最低,气孔关闭,气孔导度下降,通过调节蒸腾有效控制叶片水势,影响气孔的开关,以免过度蒸腾造成水分传输结构的损伤[14-15]。有研究表明,山杏、板栗、山桃、花椒、君迁子5种苗木蒸腾速率与叶水势显著或极显著相关,蒸腾速率的变化导致叶水势的变化[16]。本文结果也显示,在较小的时间尺度上(日进程),三倍体毛白杨叶片蒸腾速率与叶水势呈显著或极显著负相关,蒸腾速率的变化导致叶水势的变化;在大的时间尺度上(年变化),叶片蒸腾速率则与叶水势呈正相关但未达到显著水平,叶水势和其它因素共同决定和影响其蒸腾速率。
黄明斌和邵明安[6,17]在研究冬小麦和玉米叶片蒸腾速率与叶水势关系时发现,两种作物叶水势与蒸腾速率之间存在滞后现象,上午蒸腾速率增大速率大于叶水势降低速率,根系吸水速率小于蒸腾速率,组织放水,蒸腾速率等于根系吸水量和组织放水量之和,下午则相反,并认为当环境条件一致时,这种滞后现象主要由植物组织水容独特的充、放水特性所决定的。同冬小麦和玉米,三倍体毛白杨叶片Tr与CLAW、CLRW、ψ、C等水分特征也存在一定的滞后现象,如表2所示,除2008年5月26日和8月24日外,其余测定日Tr峰值出现时间都早于各水分特征值2~4 h。

表2 三倍体毛白杨叶片Tr与CLAW, CLRW、ψ、C等水分特征峰值出现时间Table 2 The daily peak-time of Tr, CLAW, CLRW, Ψ, and C of triploid clones P. tomentosa in 2008 and 2009
水容是植物固有的生物物理性状,树木水容在调节树木日季水分的动态平衡中发挥非常重要的作用[18]。2008和2009生长季三倍体毛白杨叶片水容分别为(4.48±0.73)×10-3和(5.19±1.22)×10-3g·cm-3MPa-1,低于国槐[19]。早上蒸腾开始前叶片水势、水容都处于较高水平,随着叶片蒸腾的加强,根系水分吸收速率逐渐小于蒸腾耗散,树体组织水容发挥作用而逐渐减小;当树体组织净水分储量减少到一定程度时,水容也降低到最低,其调节作用也逐渐减小,进一步导致SPAC 系统水势梯度加大,根系在较高的水势梯度作用下加快吸水以满足蒸腾需求,中午后随着蒸腾变缓,根系水分吸收速率逐渐高于蒸腾耗散,进入树体水分相比叶片蒸腾出现盈余,组织开始储水,水容回升。正是由于树体组织水容的存在,树木才得以根据环境条件的变化有效地调节自身对水分的吸收和蒸腾耗散,维持相对稳定的生理活动和生长节律[20]。
[1] 李俊清,等.森林生态学[M]. 北京: 高等教育出版社, 2006.
[2] 刘世荣,等.中国森林生态系统水文生态功能规律[M]. 北京:中国林业出版社, 1996.
[3] 刘奉觉, 郑世锴, 巨关升, 等. 树木蒸腾耗水测算技术的比较研究[J]. 林业科学, 1997, 33(2): 117-126.
[4] 张建国, 李吉跃, 沈国舫. 树木耐旱特性及其机理研究[M].北京: 中国林业出版社, 2000.
[5] 李继文, 王进鑫, 张慕黎, 等. 干旱及复水对刺槐叶水势的影响[J]. 西北林学院学报, 2009, 24(3): 33-36.
[6] 黄明斌, 邵明安. 不同有效土壤水势下植物叶水势与蒸腾速率的关系[J]. 水利学报, 1996, (3): 1-6.
[7] 奚如春, 马履一, 王瑞辉, 等. 元宝枫等3个树种枝干水容特征[J]. 中南林学院学报, 2006, 26(4): 22-26.
[8] 魏 娟, 谢福春, 王华田, 等. 水分胁迫对海州常山抗逆生理特性的影响[J]. 山东农业大学学报: 自然科学版, 2009, 40(3):371-376.
[9] 方向文, 李凤民, 张海娜, 等. 树锦鸡儿、柠条锦鸡儿、小叶锦鸡儿和鹰嘴豆干旱适应能力比较[J]. 生态学报, 2011,31(9): 2437-2443.
[10] Aston M J,Lawlor D W.The relationship between transpiration, root water uptake and leaf water potential[J]. Exp Bot, l979, 30: 249-263.
[11] 康绍忠,刘晓明,王振镒.冬小麦叶片水势、气孔阻力、蒸腾速率与环境因素的关系[J].灌溉排水,1991,10(3):1-6.
[12] 王孟本, 李洪建, 柴宝峰, 等. 树种蒸腾作用、光合作用和蒸腾效率的比较研究[J]. 植物生态学报, 1999, 23(5): 401-410.
[13] 孟德悦, 吴海勇, 刘光斌, 等. 不同种源无患子光合与蒸腾速率差异分析[J]. 经济林研究, 2013,31(2): 48-53.
[14] S. Silim, R. Nash, D. Reynard,et al.Leaf gas exchange and water potential responses to drought in nine poplar (Populusspp.) clones with contrasting drought tolerance[J]. Trees, 2009,23(5): 959-969.
[15] 周翠鸣, 赵 平, 倪广艳, 等. 基于树干液流和土壤-叶片水势梯度分析荷木干湿季整树水分利用特征[J]. 生态学杂志,2011, 30(12): 2659-2666.
[16] 董玉峰, 张 恒, 韩 俊, 等. 几个经济树种蒸腾作用对连续土壤水分变化过程的响应[J]. 江西农业大学学报, 2007,29(5): 797-800.
[17] 黄明斌, 邵明安. 冬小麦叶水势-蒸腾速率关系的滞后效应[J]. 科学通报, 1995, 40(12): 1137-1139.
[18] Fabian G S, Sandr J B, Guillermo G,et al.Biophysical properties and functional significance of stem water storage tissues in Neotropical savanna trees[J]. Plant, Cell and Environment, 2007,30:236-248.
[19] 李广德, 王晓辉, 贾黎明, 等. 国槐枝叶水分特征及其对树干边材液流的影响[J].中南林业科技大学学报,2010,30(1):23-28.
[20] 奚如春, 马履一, 樊 敏, 等. 油松枝干水容特征及其对蒸腾耗水的影响[J]. 北京林业大学学报, 2007, 29(1): 160-165.
[21] 任忠秀, 聂立水, 张 强, 等. 水氮耦合下毛白杨无性系材积生长规律及品种选择[J]. 中南林业科技大学学报, 2011,31(8): 187-193.
Regulating role and inf l uences of water characteristics of triploid clones Populus tomentosa on leaf transpiration
LI Guang-de1,2, JIA Li-ming2, FU Feng-zhen3
(1. Faculty of Agro-forestry & Medicine, Open University of China, Beijing 100039, China; 2. Key Lab. for Silviculture and Conservation of State Education Ministry, Beijing Forestry University, Beijing 100083, China; 3. Daqing Forestry Bureau, Daqing 163002, Heilongjiang, China)
To clarify the characteristics and relationships of leaf absoluteness water content (RAWC), relative water content (RRWC), water potential (ψ) and water capacitance(C) and its impacts on transpiration rate(Tr) of Triploid clonesPopulus tomentosa, the dynamic observations of 2 consecutive years onTr,RAWC,RRWC,ψ, andCwere carried out by using Licor-6400 portable photosynthesis system,oven drying method, portable pressure chamber, and so on. The results show that the diurnal variations of Triploid clonesP. tomentosa Trwas a single-peaked curve, but a “V-shape” curve forRAWC,RRWC,ψ, andC; there was a correlation betweenTrandRAWC,RRWC,ψ,C,but the correlation coeff i cients was different at different time; there exists fi fferent lag effects betweenTrandRAWC,RRWC,ψ,C, which regulated and affected the tree transpiration together with the environmental factors.
Triploid clonesPopulus tomentosa; leaf water features; leaf transpiration; water potential; water capacitance
S792.117
A
1673-923X(2014)08-0009-06
2013-10-31
国家林业公益性行业科研专项经费重大项目(201004004);北京林业大学“985”优势学科创新平台开放基金(000-1108003)
李广德(1980-),男,山西武乡人,博士,讲师,主要从事森林培育与远程教育研究;E-mail:guagndeli123@163com
贾黎明(1968-),男,山西偏关人,博士,教授,主要从事用材林和能源林培育、城市林业等研究;
E-mail:jlm@bjfu.edu.cn
[本文编校:吴 彬]
猜你喜欢
科学养鱼(2021年11期)2022-01-11
干旱地区农业研究(2021年6期)2021-12-08
新农业(2021年15期)2021-08-23
热带农业科技(2020年3期)2020-12-15
新农民(2020年21期)2020-12-08
农村实用技术(2020年9期)2020-11-19
河北林业科技(2020年3期)2020-01-10
湖北林业科技(2019年4期)2019-09-16
食品与生活(2019年12期)2019-05-15
东坡赤壁诗词(2018年1期)2018-03-31
