
北亚热带地区不同生境外来植物分布比较
2013-12-27 07:37郑景明刘艳红刘忠华
中南林业科技大学学报 2013年7期
袁 帅,郑景明,刘艳红 ,刘忠华
(北京林业大学 a.省部共建森林培育与保护教育部重点实验室;b.生物科学与技术学院,北京 100083)
北亚热带地区不同生境外来植物分布比较
袁 帅a,郑景明a,刘艳红a,刘忠华b
(北京林业大学 a.省部共建森林培育与保护教育部重点实验室;b.生物科学与技术学院,北京 100083)
亚热带地区是外来种入侵的活跃区,林改后人类活动增加可能会加大外来种入侵风险。对地处我国亚热带北缘地区的九江、星子两县不同生境的外来植物种组成丰富度和区系起源等情况进行了抽样调查。在森林、林缘、田边、路缘4类生境中共计调查发现外来植物28种,因为光照条件和人为干扰状况的不同,基于种数的方差分析结果显示外来植物种种数在林内、林缘、农田边缘和路边呈依次增高的趋势;盖度方差分析结果显示外来植物种数在林内、林缘、道边、农田边缘呈增高趋势,支持干扰促进外来种发生的假说。对各个生境内部本地种和外来种的相关性分析结果显示,基本上各个生境内部本地种种数和外来种种数呈显著的负相关;而本地种盖度和外来种盖度呈负相关性但不显著,支持丰富度高的群落入侵抵抗力强的假说。最后,分析了亚热带北缘地区各类生境中外来种组成的特征,对亚热带地区不同生境的外来种防控提出了建议。
外来植物;入侵;亚热带北缘地区;植物生境;相关性分析
全世界范围内,由于人类活动破坏天然植被及人类对土地的各种利用形式,加剧了外来植物种类的引入和传播,为外来植物种创造了新的定居环境。外来植物种已经成功入侵许多生态系统,这些入侵种对生态系统的结构和功能产生了很严重的负面影响。我国地域辽阔,南北跨越5个气候带,这样使得世界各地的外来种都能找到合适的栖息地,决定外来入侵植物物种数的主要的因素是气候及人类活动的强度[1]。我国的外来种共调查到800种,有80%是人为有意引入。整体分布不均衡,在东南沿海及南方分布较多而在西北、西部地区分布较少。亚热带地区是外来种种数仅次于热带和温带湿润区的我国第三大外来种分布区[2]。亚热带地区的外来植物数量较多,同时亚热带也是我国人类经济活动最为活跃的地区,容易发生入侵。我国各种生态系统包括农田、森林、灌丛、草原、城市和水生生态系统等都有外来种的分布。其中,农田、森林、和植物园等人类干扰强的生态系统含有80%以上的外来种[3]。
江西省九江市属于我国林改试点区,林区经营权归个人以后引发的许多问题中,由于人类活动对森林的干扰将有所增大,外来种入侵的风险是否会增加尤其需要加紧研究[4]。本文调查了亚热带北缘的江西省九江市的九江、星子两县不同生境类型中的外来植物分布情况,包括主要森林类型内部及其林缘、农田边缘、道路生境中外来植物种类组成、盖度等状况。同时,以往对单一生境的外来种情况研究较多,但对不同生境间外来种情况的横向比较较少,尤其是对亚热带森林生境中外来种种类和分布的研究少见报道。本研究旨在探究以下问题:(1)不同人类干扰程度的生境类型中外来种组成和数量有无差别?即是否符合干扰促进外来种入侵的假说;(2)不同生境中外来种丰富度与本地种丰富度之间是否存在负相关?即是否支持物种丰富度高的群落对外来种入侵抵抗力强的假说[5]。
1 试验地概况
调查地位于东经115.60~116.77,北纬29.13~29.87,属亚热带北缘,处东亚季风区,气候温和,四季分明,全年日照充足。年无霜期为239~266 d,年平均雾日在16 d以下,年平均湿度达75%~80%。多年平均气温16~17 ℃,山区略低一些。多年平均降水量1 400 mm,区域分布明显,垂直差异性大,而且雨季旱季比较分明,其中50%左右集中在春夏之交,雨量分配明显不均。有利于喜温作物和林木生长。适宜发展种植业和畜禽、水产等养殖业,物产丰富,生态环境良好[6-7]。
2 研究方法
2.1 样地布设
对不同类型森林内部及林缘、农田和路边等生境类型设立样地,调查其外来植物状况。森林样地的选取是根据调查范围内各样森林类型分布状况,随机抽样对各样森林类型进行林内、林外的外来植物调查。此次研究共调查各样森林样地34个,包括阔叶林样地9个、针阔混交林样地6个、毛竹林样地6个、马尾松林样地7个以及杉木林样地6个。同时在森林样地附近共布设31个林缘样地;农田以及路边生境方面,共计对39个农田边缘样地以及25个路边样地进行调查。其中森林样地面积20 m×30 m,调查林分乔木组成,采用5个5 m×5 m小样方调查灌木组成;样地里设置5个1 m×5 m的草本样方,调查草本组成。林缘样地即沿着森林样地附近的林缘处设置2 m×50 m的样地,在内部随机布设5个1 m×5 m的草本样方,调查草本组成。农田以地块为单位,在农田周围随机设置5个1 m×5 m的草本样方,调查草本组成;道路路缘样地设置以公路两侧2 m×50 m的路缘生境为单位,随机设立5个1 m×5 m的草本样方,调查草本组成。
2.2 野外调查
野外调查于2011年7月份进行,由于本次调查的外来植物都是草本,在每个生境样地中采用5个1 m×5 m草本小样方调查外来种和本地种数量,记录样方内外来种和本地种的物种名、盖度和平均高度,同时记录调查样地的经纬度、海拔、坡向、坡度、林内郁闭度和样地干扰程度,包括小路、除草痕迹、是否有倒树、动物的干扰、以及大石块等。
3 数据处理
对比森林类型之间,林内平均种数及平均盖度数据因其不符合正态分布,用非参检验K-W法进行显著性检验。对比各个不同生境类型,物种数符合正态分布,盖度数据用平方根转换后符合正态分布,都用单因素方差方法分析。各个生境之间,对外来种种数与本地种种数之间进行了相关分析。都由SPSS18.0完成。
4 结果分析
4.1 外来种区系组成
本次调查共调查草本植物132种,其中外来植物28种。其中菊科6种、十字花科1种、苋科5种、藜科4种、商陆科1种、锦葵科1种、大戟科1种、豆科2种、桑科1种、车前科1种、旋花科2种、马鞭草科1种、禾本科1种、蓼科1种。其中林下分布有外来植物14种,林缘外来植物27种,农田分布有20种,路边分布有24种。其中林内林缘共有种为14种,林内农田共有12种,林内道路共有种为14种,林缘农田共有19种,林缘道路边共有种24种,农田道路共有17种。本次调查的外来植物区系成分及起源如表1。
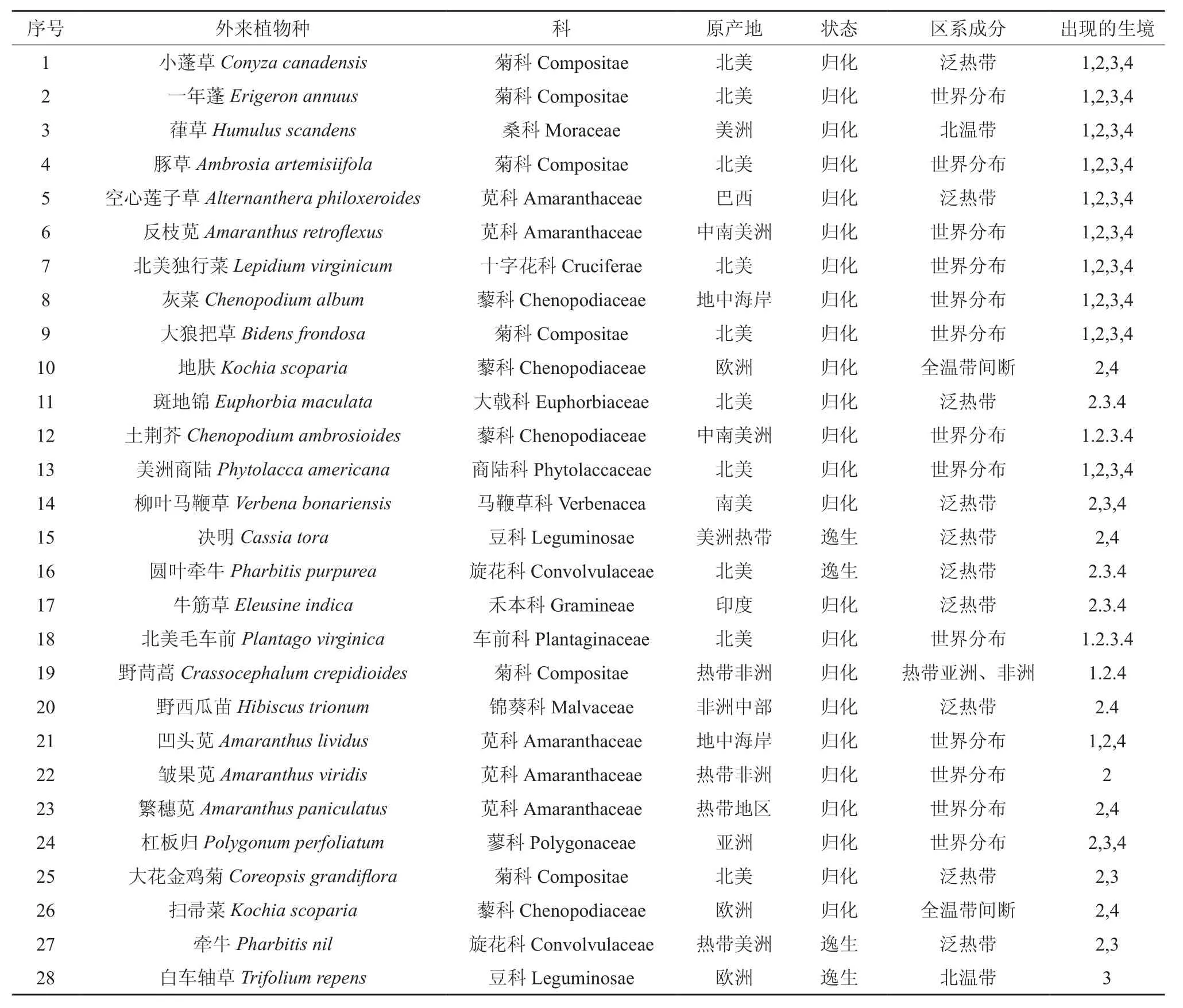
表1 外来种植物区系成分及起源†Table 1 Composition and origin of alien plants in sampling plots
从科的分布上看,菊科最多为6种,苋科为5种,藜科4种,桑科1种,十字花科1种,大戟科1种,商陆科1种,马鞭草科1种,豆科2种,旋花科2种,禾本科1种,车前科1种,锦葵科1种,蓼科1种。
外来种起源方面,源自北美的外来种最多为10种,热带非洲及美洲地区8种,源自欧洲、南美等其他地区10种,来自于北美洲的外来种最多,这是因为北美洲与亚洲同处于北半球且纬度相近。这也与国际贸易、旅游的发展和人们交往的增加以及全球环境的变化息息相关。
从归化状态上看,其中,决明、圆叶牵牛、牵牛和白车轴草4种植物处于逸生状态,即尚没有对当地构成危害;剩下的24种已经在当地建立稳定种群,部分已经分布十分广泛。
从区系成分上看,世界分布种为13个,其次为热带11种,温带起源仅有4种,表明当地主要外来种还是适应亚热带气候的。
从分布的生境类型看,小蓬草、一年蓬、葎草、豚草、空心莲子草、反枝苋、北美独行菜、灰菜、大狼把草、土荆芥、美洲商陆、北美毛车前12种外来种是四类生境共有的;除此以外,野茼蒿、凹头苋是林内、林缘和林内、道路共有的;林内和农田共有种即为四类生境共有种;斑地锦、马鞭草、圆叶牵牛、牛筋草、杠板归、大花金鸡菊、牵牛是林缘和农田共有的;地肤、斑地锦、马鞭草、决明、圆叶牵牛、牛筋草、野茼蒿、野西瓜苗、凹头苋、繁穗苋、杠板归、扫帚菜是林缘和道路共有的;斑地锦、马鞭草、圆叶牵牛、牛筋草、杠板归是农田和道路共有的外来种。
4.2 各样生境外来种组成比较
首先对不同森林类型的林内外来种的物种数和盖度进行差异显著性检验,因数据不符合正态,故采用K-W法(α=0.05),检验表明各类森林类型林内外来种数量无差异(种数P=0.476;盖度P=0.068);同时对林缘数据进行单因素方差分析检验,不同的森林类型林缘的外来种平均种数不存在显著差异(种数P=0.320;盖度P=0.393),因此,在后面的分析中,分别合并各类森林类型的林内样地及林缘样地,只作为林内和林缘两类生境处理。
对四类生境中外来种平均种数数据进行单因素方差分析,结果P<0.01;外来种平均盖度单因素方差的P<0.01。多重检验表明,在外来种平均种数方面,只有林缘和农田两生境差异不显著(P=0.513),剩下生境间皆显著(P<0.01);在外来种平均盖度方面,只有农田和道路生境间的差异不明显(P=0.387),余下生境之间的差异都非常显著(P<0.01)(见图1)。
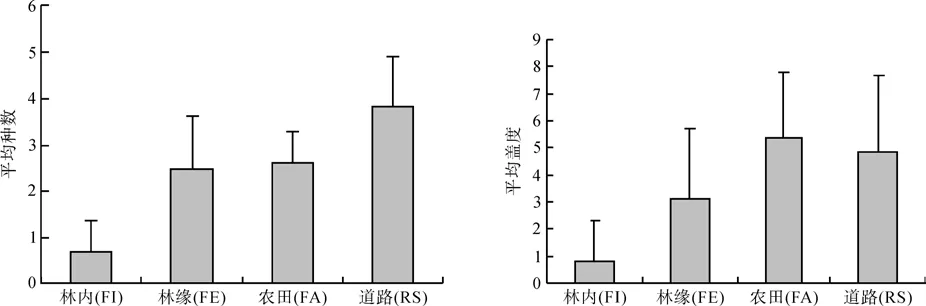
图1 不同生境外来种平均种数和盖度比较Fig. 1 Exotics species’ average and coverage among different type of habitats
4.3 生境内部外来种与本地种相关性分析
在林内调查中,总共发现93种草本植物,其中有14种外来植物;林缘样方总共128种草本植物,其中27种外来植物;路边样方总共93种草本植物,其中有20种外来植物;农田共发现76种草本植物,外来种有20种。对各个生境的1 m×5 m小样方的外来种物种数、本地种种数和外来种盖度、本地种盖度数据分别进行spearman相关性分析,见表2。
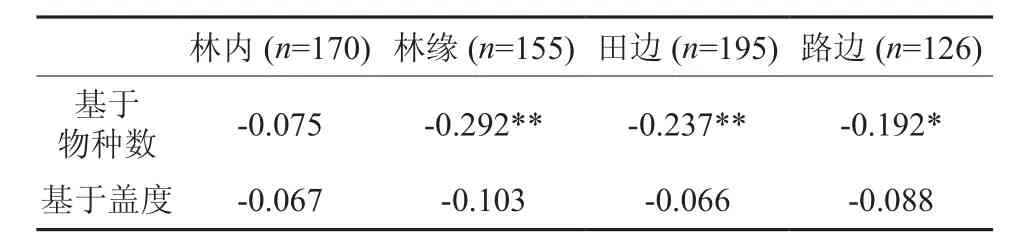
表2 不同生境中外来种和本地种的spearman相关系数†Table 2 Spearman correlation coefficients of alien and native plants among different type habitats
结果表明,在不同生境内,外来种数与本地种数呈负相关。基于物种数的相关分析中,除林内因外来种数过少而相关性不显著外,其余3类生境均为显著相关;而基于盖度的相关分析中,4种生境均为不显著,且相关系数都很低。
5 讨 论
根据以上结果,林内、林缘、农田、道路四样地内,平均种数和平均盖度的假设检验的结论均是林内、林缘、农田、道路边的外来植物种有显著差异。究其原因,首先,光对外来入侵植物的分布有重要影响,林内和林缘的外来物种差异不难理解,因为林缘样地因其处于林内和林外之间过渡的位置并且阳光照射相对充足,同时,外来物种大多都是阳性物种,在林内高郁闭的环境内很难生长,故林缘外来种更加丰富,林内的外来植物种在林缘均能找到。林内外来种植物种数和盖度最低正是因为光线条件的限制其外来种数相对较少,与以往研究结果类似[8],林缘生境比农田和路边生境的光照要少一些,故外来种数量也比农田田边和公路路缘相对较低。
其次,生境环境的扰动是造成外来种建群的主要原因之一,因为扰动导致的短期内资源的迅速增加为一些能较快利用这些资源的外来种提供了种群迅速增大的机会[9]。同时,人类活动可以提高受干扰生境的繁殖体压力,旅行者通过衣服、鞋和交通工具携带大量种子带入到被干扰的生境,几乎所有路边的植物的种子都能被人类携带进入新的生境[10-11]。所以人类干扰越频繁的地方就容易出现更多的外来种,这可以解释为什么林内、林缘、农田、道路边的外来植物丰富度呈由低到高的趋势。但是,在有些人类活动不频繁的森林内部也出现了外来植物,这就有可能是非人类干扰(动物、树倒等)因素的影响[9]。平均种数的检验中,林缘和农田之间的检验未呈现显著差异(P=0.513),这可能是因为调查到的当地农田多数为水稻田,这样其周围丰富的水养吸引一些喜湿喜水的外来种建立优势种群,进而降低了生境植物种数而导致农田外来种组成趋向相对单一。调查过程中发现,空心莲子草和牛筋草是农田生境中出现最频繁的外来种。盖度检验中,原本是道路的人为干扰要比农田要多,但农田盖度与道路盖度之间的差异不显著且农田的平均盖度超过了道路(P=0.387),这可能是因为农田因其丰富的水分和土壤养分致使其植物生长相对其他生境旺盛,故其外来植物种盖度相对要大。此外,路边、农田、林缘、林内四种生境的复杂程度是由低到高的,生境越复杂就越不容易被外来种入侵[12],这可能也是三种生境外来种的分布趋势的另一原因。
再者,外来种与本地生物环境间的相互作用是影响外来种分布的重要因素,作用包括种间竞争作用[13]和协作[14]作用等。土著种可以通过争夺光、水分、营养等抑制外来种,某些外来物种入侵新的生境,因为抢夺资源而获得优势[15]。本文对本地种和外来种之间关系的分析表明,整体上外来种与本地种之间种数与盖度的关系都呈负相关,即本地种和外来种存在着显著的竞争关系,本地种对外来种的生长有一定的抑制作用。在外来种和本地种种数之间关系的检验中,林内的检验并不显著,这可能是因为林内植物种稀疏并且林下的资源丰富,这样本地种的生长对外来种没有构成对水、营养物质等的激烈争夺,所以两者之间竞争作用相对较弱。盖度的检验中,农田外来种盖度和本地种盖度之间呈正相关,区别于其他生境检验。这可能是由于农田周围因其具有多于其他生境的土壤养分和水分,以至于其周围的植物生长繁茂,所以在盖度上没有体现出本地种与外来种之间的互相抑制关系。另外,盖度的检验中,四类样地P值都大于0.05,表明可能种间的竞争不是很剧烈。这可能是由于本次调查的外来种除了个别种如空心莲子草,美洲商陆,在生境中多数没有形成优势格局,尚未达到入侵程度,仅处于归化状态,因而可以与本地种的共存而没有发生明显的竞争排斥。
我国亚热带地区经济比较发达,人类活动频繁,对自然生态系统的干扰也比较严重,因此外来种入侵发生概率较大。森林这个陆地上最重要的生态系统,虽然由于其郁闭度大,物种丰富,通常很难造成大规模入侵,然而在人类干扰条件下,热带和亚热带森林同温带森林一样,都可能发生严重的外来种入侵[8,16-17]。所以,面对森林生态系统的入侵威胁防胜于治,尤其要重视森林(林缘、林内)的外来种植物的早期探测,加强外来种入侵预警监测和防治,一方面,对有可能入侵的外来种加强地区检疫等防范措施与并研制相应的应急控制技术[5];另一方面,对已经入侵的外来种进行监测和实施有效技术方法给予控制[18]。对于已经被入侵的森林的修复和外来种防除,国外有研究表明,林边人工围栏可阻止外来种侵入和促进本地种的生长[17]。其次,研究表明,亚热带地区的外来植物多以来自平原地区的农田杂草外来种为主,逐渐扩散到山地丘陵等其他区域[19-20]。亚热带地区的农业发达,农田生境因为其人类活动频繁、水养条件更适合外来种生存,也是外来种向其他生境(森林、草地等)的传播中转站,所以对农田田边地头荒地的外来种防控不能忽视。针对农田与人们生产生活直接相关等的特殊情况,可发展特殊的、可行的消灭或控制外来种的技术方法,制定最佳的治理、控制外来种的方案与技术。小面积入侵可进行高效的、紧急扑灭如机械防除等,对爆发性入侵要进行化学清除及一次性消灭[18]。再次,有关研究表明[21],道路作为景观廊道,不但增加了边缘效应,而且易造成外来种的入侵。道路对临近生境比如森林等生境外来种传播起很大作用[22]。路缘是一个外来种存在和传播的重要生境,针对道路的外来种的通道作用进行防控可能更重要[23-25]。
亚热带地区气候温暖湿润,人类干扰频繁,各类生境被入侵可能性高;九江地区,尤其是庐山保护区周围的九江和星子2县及九江市部分市区,因为毗邻庐山这个江西省自然保护区和国家地质公园及全国知名的旅游名胜地,面临较大的外来种入侵风险,特别是林改以后本地林农将可能增加对森林的经营管理,从而将促进外来植物繁殖体的传播和增强对自然环境的干扰,自然保护区的外来种防控任务将更为艰巨。本研究中所调查到的一些的外来种,如空心莲子草、土荆芥都列入我国政府的入侵种名录,鉴于其已经出现在林缘生境,对其可能出现更大范围的入侵和扩散应引起相关部门的重视并及时采取措施防治。
[1] 吴晓雯, 罗 晶, 陈家宽, 等. 中国外来入侵植物的分布格局及其与环境因子和人类活动的关系[J]. 植物生态学报, 2006,30(4): 576-584.
[2] 杨 博, 央金卓嘎, 潘晓云,等. 中国外来陆生草本植物:多样性和生态学特性[J]. 生物多样性, 2010, 18(6): 660-666.
[3] 丁 晖,徐海根, 强 胜, 等. 中国生物入侵的现状与趋势[J]. 生态与农村环境学报, 2011,27(3): 35-41.
[4] 张 颖, 刘 丹, 李 坦, 等. 林权改革对环境影响的统计分析研究[J]. 中国水土保持科学,2012,10(2): 83-87.
[5] 郑景明, 马克平. 入侵生态学[M]. 北京: 高教出版社, 2010.
[6] 赵 磊,葛 刚,刘以珍,等. 庐山保护区外来种子植物分析[J]. 江西科学,2008, 26(1): 155-169.
[7] 万慧霖,冯宗炜,庞宏东.庐山外来植物物种[J].生态学报,2008, 28(1): 103-110.
[8] 董红云, 李 亚, 汪 庆, 等. 江苏省3个自然保护区外来入侵植物的调查及分析[J]. 植物资源与环境学报, 2010,19(1):86-91.
[9] Hertling U M,Lubke R A. Assessing the potential for biological invasion : The case Ammophlia arenaria in the South Africa [J].South African Journal of Science,2000,96:520-527.
[10] Mack R N, Lonsdale W M. Humans as global plant dispersers:getting more than we bargained for [J]. Bio Science, 2001, 51:95-102.
[11] Schmidt W. Plant dispersal by motor cars [J]. Vegetatio, 1989,80: 147-152.
[12] 汤敏喆, 任明讯, 郑景明, 等. 加拿大一枝黄花对庐山自然保护区林地植物多样性及其季节动态的影响[J]. 植物科学学报,2012, 30(4): 366-373.
[13] 林金成, 强 胜. 空心莲子草对南京春季杂草群落组成和物种多样性的影响[J]. 植物生态学报, 2006, 30(4): 585-592.
[14] Vilá Montserrat, Jacob Weiner. Are invasive plant species better competitors than native plant species? – evidence from pair-wise experiments [J]. Oikos, 2004, 105(2): 229-238.
[15] Usher M B.Biological invasions of nature reserves:A search for generalization [J].Biological Conservation, 1988, 44: 119-135.
[16] 秦卫华, 王 智, 徐网谷, 等. 海南省3个国家自然保护区外来入侵植物的调查和分析[J]. 植物资源与环境学报, 2008,17(2): 44-49.
[17] Fine P V A. The invisibility of tropical forests by exotic plants[J]. Journal of Tropical Ecology, 2002, 18: 687-705.
[18] 万方浩, 郭建英, 王德辉. 中国外来入侵生物的危害与管理对策[J]. 生物多样性, 2002,10(1): 119-125.
[19] Alexander JM, Kueffer C, Daehler CC, et al. Assembly of nonnative floras along evelational gradients explained by directional ecological filtering [J]. Proceedings of the National Academy of Sciences, 2011, 108: 656-661.
[20] Keith L. Mcdougall, Anzar a. Khuroo, Lloyd L. Loope, et al. Plant Invasions in Mountains: Global Lessons for Better Management [J]. Mountain Research and Development, 2011,31(4): 380-387.
[21] 周 婷,彭少麟,林真光. 鼎湖山森林道路边缘效应[J]. 生态学杂志, 2009, 28(3): 433-437.
[22] Cara R Nelson, Charles B Halpern, James K Agee. Thinning and burning result in low-level invasion by nonnative plants but neutral effects on natives [J]. Ecological Applications,2008,18(3):762-770.
[23] 郑景明, 徐 满, 孙 燕, 等. 庐山自然保护区内外路缘外来植物对比[J]. 北京林业大学学报, 2011,33(3): 51-56.
[24] 曹福祥,祁承经,喻勋林,等.中国中部亚热带常绿阔叶林保育的重要性及其恢复重建策略[J].中南林业科技大学学报,2010, 30(11): 95-104.
[25] 李志辉,李柏海,祁承经,等.我国南方珍贵用林树种资源的重要性及其发展策略[J].中南林业科技大学学报,2012,32(11): 1-8.
Comparison of exotic plants’distribution among different habitats in northern subtropical region in China
YUAN Shuaia, ZHENG Jing-minga, LIU Yan-honga, LIU Zhong-huab
(a.Key Lab. for Silviculture and Conservation of Ministry of Education; b. College of biological science and technology, Beijing Forestry University, Beijing 100083, China)
Vegetation in subtropical is vulnerable to biological invasion. After the forest property reform, human activities in the forests increased and the natural vegetation was disturbed, these probably led to the higher risk of biological invasion in this region. Four types of habitats were randomly sampled in Jiujiang and Xingzi County in the north rim of subtropical region and the number and coverage of native and alien plant species were recorded. The statistical analysis based on species number and coverage were carried out. The results show that there were 28 alien plant species occurred in four types of habitats and there were signif i cant differences in number and coverage of alien species among forest interior, forest edge, farmland border and roadside. ANOVAs in terms of species number resulted in the increase of alien species from forest interior, forest edge, farmland border to roadsides, whereas the ANOVAs in terms of coverage led to an increase of alien plant from forest interior, forest edge, roadside, to farmland border. Overall, the ANOVAs results support the hypothesis that disturbance enhances biological invasion. The correlation analysis between native species and alien species showed that the number and coverage of native and alien plant species were negatively correlated, although p value was lower than 0.05 only for analysis based on species number. Thus correlation analysis supports the hypothesis that the species-rich community was more resistant than species-poor communities. The composition of alien plants species in the region was analyzed and some suggestions on prevention and management of alien plant invasion for different habitats were put forward, especially in area closed to the Nature Reserves.
exotic plants; invasion; northern edge of subtropical areas; plants habitats; correlation analysis
S718.53
A
1673-923X(2013)07-0025-06
2012-10-16
国家林业公益性行业科研专项(200904003);国家自然科学基金项目(30940006)
袁 帅(1987-),男,内蒙古赤峰人,助理研究员,主要从事入侵生态学的研究;E-mail:nmyuanshuai@126.com
郑景明(1971-),男,黑龙江鸡西人,副教授,主要从事入侵生态与森林生态研究; E-mail:Zhengjm@bjfu.edu.cn
[本文编校:吴 毅]
猜你喜欢
现代园艺(2021年13期)2021-07-05
南方农业·中旬(2020年9期)2020-11-30
黑龙江工程学院学报(2020年5期)2020-10-21
亚热带农业研究(2019年2期)2019-09-05
园艺与种苗(2019年1期)2019-03-13
文学教育(2018年27期)2018-08-24
绿色科技(2017年15期)2017-09-01
文理导航(2017年22期)2017-08-16
星星·诗歌原创(2017年5期)2017-08-13
环境与发展(2017年4期)2017-07-19
