
四川盆地钉螺生境指示植物研究
2013-12-27 07:37蒋俊明周金星费世民刘国华
中南林业科技大学学报 2013年7期
蒋俊明,周金星,费世民,刘国华
(1.中国林业科学研究院,北京 100091;2.四川省林业科学研究院,四川 成都 610081)
四川盆地钉螺生境指示植物研究
蒋俊明1,2,周金星1,费世民2,刘国华2
(1.中国林业科学研究院,北京 100091;2.四川省林业科学研究院,四川 成都 610081)
文章以四川仁寿山丘疫区157个样方钉螺密度、草本层特征调查数据为基础,采用方差、X2分统计度量、Ocihiai、Dice、Jaccaed、Pccc等系列技术分析法,研究了钉螺生境指示植物及其种间联结度。结果显示:在85个草本群落中,与钉螺有极显著正连接的有水芹、水金凤、毛茛、拉拉藤、空心莲子草、广布野豌豆6种,显著正连接的有五朵云、蜈蚣草、碎米荠、葎草等4种;有极显著负连接的有野蔷薇、苔草、金星蕨等3种,显著负连接的种类有凤尾蕨1种;Pearson相关分析表明,土壤水分含量、土地利用类型是决定钉螺分布密度的两大关键因子;Ocihiai、Dice、Jaccaed、Pccc种间系数达到一定阈值才能达到正连接或负连接的水平,四个系数与X2拟合均呈线抛物线关系;钉螺生境可分划分为极度适生、一般适生与不适生三类,后者是前者退化与灭螺改造的结果;林业血防生态工程改造效果可用钉螺不适生来描述,其标志为正连接指示植物消失而负连接指示植物出现,钉螺指示植物可用于作钉螺生境识别,也可用作林业血防生态工程模式林的效果评价。
钉螺;钉螺生境; 指示植物;山丘疫区;种间连结性;血防林评价指标
林业血防的兴起,改变了传统灭螺方式,30多年在我国水网型和湖沼型疫区的实践证明,该方法是行之有效的。2006年林业血防工程作为一项重大生态项目在全国疫区实施,但该项生态控螺技术应用于山丘型疫区还有很多问题需要解决,主要集中于:其一,山丘区林业血防建设是以是小流域为单元,层层营造抑螺防病林带,对钉螺实施阻断、隔离和杀螺等[1],其治理对象不单纯是控制钉螺,而是对疫区破碎生态系统的整治和修复;单一钉螺密度和有螺面积的变化指标不能反映对钉螺生境的改造效果;其二,钉螺个体较小[2],查找钉螺不仅费工、费时,能否通过钉螺环境指示植物进行评估,这方面国内一些学者已通过3S技术在宏观尺度对钉螺生境的识别[3-7],这证明了这种想法的可行性。山丘区钉螺生境识别依靠3S技术只能是河流库塘等湿地大类型的边缘进行评估,但很多小生境夹杂在很多生境的间隙或中央,是生境不均一的结果,无法借助远距离测试手段来精确找寻或识别钉螺生境。
钉螺与植被、土壤因子均存在显著的关联性[8-11],这些研究表明钉螺存在一个相互作用、相互依赖的草本生态系统。种间连接(Inter-specif i c association)是指不同物种在空间分布上的相互关联性,反映了生态位的重叠与分离度。基于钉螺栖息于草本群落生境,即这些草本层为钉螺营造的一种适生环境,同时也为钉螺提供了食源,这些草本群落和种类多为湿地植物,即这些草本和钉螺有一个湿生环境,两者对生境条件的要求具有共性,往往指示植物在群落中为优势种类时,此种优势种与钉螺之间更容易建立一种正连接关系,更容易在景观尺度上识别或测定钉螺生境面积。这些与钉螺呈现正连接的物种能构成钉螺分布的指示种类,且具有统计学上的概率显著性。
基于上述问题,开展对山丘型疫区钉螺及生境因子调查和研究,采用种间连接分析法探讨山丘区与钉螺生存的共生植物、共存的环境特性,寻找钉螺生态系统同生态位种团,一则深入认识钉螺生境特性,二则找寻钉螺识别的指示植物,三则建立血防林效果评估的指标。
1 材料与方法
1.1 研究样地概况
试验区位于四川省仁寿县,属四川盆地紫色丘陵区。该区气候为亚热带湿性气候,累年平均气温17.4 ℃,全年气温以7月最高,为26.5 ℃,1月最低,为6.8 ℃,全年极端最低温为-3.8 ℃,总正积温6 360 ℃,≥10 ℃的积温为5 532.8 ℃;全年无霜期310 d左右,最长356 d,最短282 d。历年平均降雨量1 009.6 mm,最低765.6 mm,年际间变幅644.6 mm。累年平均蒸发量1 355.7 mm,以4~8月为最大,月蒸发量超过150.0 mm,3月和9月次之,亦大于100 mm。历年平均日照1 196.6 h,年日照率27%,太阳总辐射89千卡/cm2,日照时数和总辐射为全国低值区。典型植被为马尾松、柏木林,混生有青冈、慈竹、苦楝等树种,林下草本植物为茅草、蕨类,灌木为油茶等,流域内大多为农地。
仁寿县是四川省最早报告发现血吸虫病人的地方,属山丘型重度疫区。全县血吸虫病流行乡镇31个,人口85.27万人。流行村302个,人口39.75万人。历史钉螺面积1 590.56万m2,历史血吸虫病病人30 772人,人群感染率17.3%。
1.2 调查方法
(1) 植被调查:林分调查:进行每木调查,实测林木的胸径(cm)、树高(m)、冠幅(m×m)和枝下高(m);林下灌木及草本设置小样方进行群落学特征调查;草本样方调查:按土地利用类型等,按等距离(10 m)选择样点、每样方按1 m×1 m调查植物种类、株(丛)数、植株高度、盖度、生活力等。
(2) 钉螺调查:依据钉螺调查方法,结合样带、样地及样方调查,螺框面积33 cm×33 cm;记录统计查螺框数、有螺框数、活螺数、死螺数、活螺框出现率与活螺密度等。
(3) 土壤调查:土样采集:根据土地利用/覆盖分类的不同,对典型植被类型挖土壤剖面,详细记录剖面信息;环刀采集原状土,混合采集表土(0~20 cm)样品1 kg左右,供室内分析测定用。烘干法测定土壤含水量,环刀法测定土壤容重或紧实度。在样地调查时,用土壤水分速测仪(HH2)测定土壤温度、土壤水分和土壤电导率。
1.3 数据分析方法
以钉螺为目标种,统计共存的种类与相离的种类样方数,N为样方总数,a为两物种共同出现的样方数,d为两物种都不出现的样方数,b,c为分别仅有一个物种出现的样方数。
种间关联可用四种成对种间匹配系数计算[12-13]:
①Jaccard(JI)系数,即共同出现百分率PC(percentage co-occurrence):JI=a/(a+b+c);
②Dice系数(DI),即重合指数(coincidence index):DI= 2a/(2a+b+c);
③Ochiai系数(OI):OI=a/[(a+b)(a+c)]1/2;
此外,还用点相关系数计算关联度,用贡献度系数来计算盖度贡献度,群落盖度对钉螺生境具有决定性意义,对钉螺群落稳定种间关系形成具有重要作用。
④点相关系数:PCC=(ad-bc)/[(a+b)(a+c)(b+d)(c+d)]1/2;
⑤贡献度系数(CI):CI=Ns/C,Ns为单位样方面积钉螺的数量,C为矫正盖度大小;
种间关联有正负2种,若a>(a+b)(a+c)/(a+b+ c+d)为正,反之为负。
正负连接度由下式计算可得:R=a-(a+b)(a+c)/(a+b+c+d),正值为正连接,表示二者相遇的概率;负值为负连接表示二者相离的概率。
种间连接强弱有X2决定: X2=(ab-bc-0.5N)2.N/(a+b)(b+c)(b+d)(c+d)
若X2>3.841(0.05<P<0.01),则表示种间连接显著;若X2>6.635(P<0.01),表示种间连接性极显著;若X2<3.841(P<0.05),表示种间不存在任何关联。
计算单一样方的物种丰富度、覆盖度、草本高度,用SPSS统计分析土壤因子、草本群落因子与钉螺数量或密度之间的相关性矩阵,分析影响钉螺分布密度的因子及其作用因子。
正负连接物种分布区域查询与钉螺生境重叠度评价,判定共生植物的专一性。
2 研究结果
调查土地类型为:林地,耕地、农田、沟渠、河滩草地等。钉螺分布密度(只/0.11 m2)大小排序为:沟渠(8.89)>水田(8.56)>河滩草地(7.49)>荒草坡(3.71)>旱地(耕地1.50)>林地(0.18)。本文统计样框数为157个,有螺框数为49个,有螺框出现率:31.2%。并将有螺框数出现的草种进行关联分析,共出现草种85种,有正连结关系的68种,负连结关系的17种。
2.1 几个参数计算结果比较分析
几种计算方法均反映了钉螺与这些草种的联结关系,其中贡献度是将钉螺密度与该草种所占盖度的乘积,是钉螺密度的量化指标,正负值是判断该草种对钉螺促进或抑制,为零时,表示两者完全无关,将这几种计算进行相关性分析结果见表1。
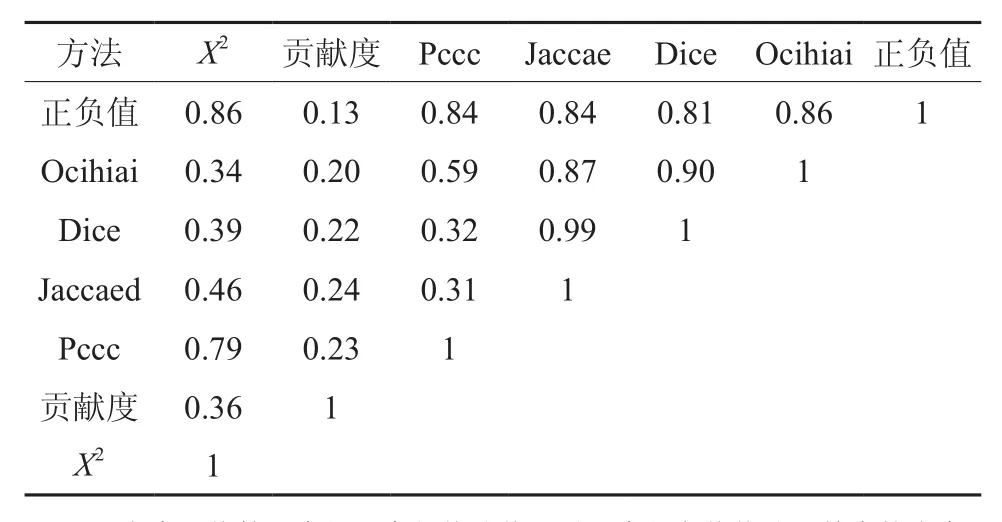
表1 7种种间连结度计算结果的相关性(R2)分析(f=84)Table 1 Correlation (R2) analysis of inter-specific coupling degree among seven methods (f = 84)
表1可知,贡献值虽为钉螺密度的直观指标,但其它6种方法计算的R2值均较小,说明钉螺密度的大小与该物种盖度的大小关系较小。从R2值大小看,可分为两组指标,PCC、正负值和X2可为一组,3个相关系数较高;正负值计算结果与其它五个公式计算值相关系数(R2)圴大于0.8,说明这个具有通用性;Dice和Jaccae两者相关系数达0.99,说明两个指标作为物种重合重现的一致性,但Dice值明显大于Jaccae值;Ocihiai与Dice和Jaccae的相关系数R2分别为0.87和0.90,均达到显著标准,说明3个指标在评价物种关联时的共用性,但计算值以Ocihiai最大。
2.2 钉螺密度与群落因子相关分析
通过Pearson相关分析,含水率、土地利用类型、土壤温度与钉螺密度分别呈极显著、轻微显著和极显著相关关系,表明非生物因子直接决定了钉螺密度。土地利用类型与草本群落盖度、高度、含水率、土壤温度等存在极显著相关性,表明土地利用类型自身存在上述群落及其环境因子的显著差异(见表2)。

表2 钉螺密度与生物群落、土壤因子的Pearson相关分析†Table 2 Correlation analysis among oncomelania density and biological community, soil factor by Pearson method
2.3 钉螺与草种间连接分析
与调查群落物种涉及85种,通过种间关系的X2分析,与钉螺种间连接关系呈现不显著的有71种,显著5种,极显著的9种,表3列出达到显著以上的物种计算结果。
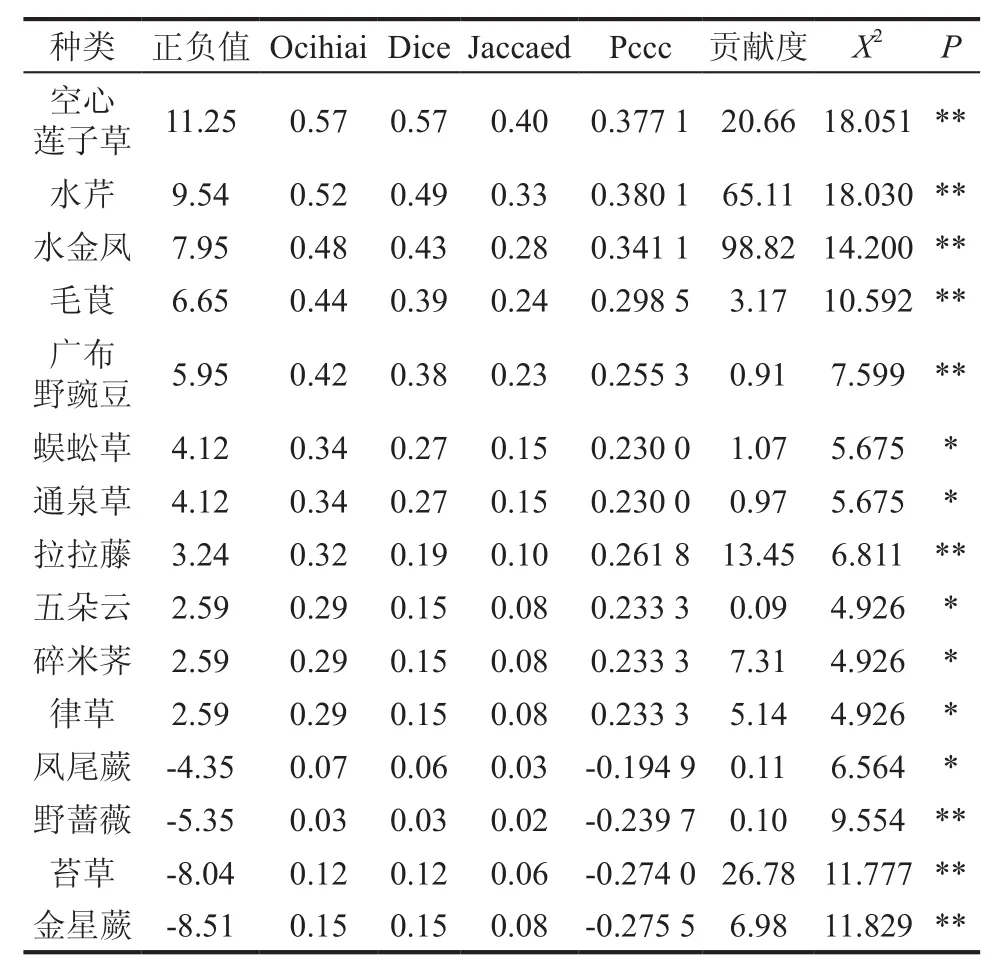
表3 钉螺与草本群落间的关系系数显著性计算†Table 3 Significant calculation of relationship coefficients between oncomelania and herbosa
极显著正连接的种类:空心莲子草、水芹、水金凤、毛茛、广布野豌豆、拉拉藤、等6种;显著正连接的种类:蜈蚣草、通泉草、五朵云、碎米荠、葎草等4种;极显著负连接的种类:野蔷薇、苔草、金星蕨等3种;显著负连接的种类有凤尾蕨1种(见表3)。正负连结度与Pccc一致,均存在正负,能很反映物种间相互关系,而Pccc值与X2变化特征完全一致。贡献度虽是钉螺密度表征,但值的大小并不能完全对应草本种间关系。
2.4 钉螺与草种间关联度的显著性分布
X2值反映了显著性程度或梯度。钉螺种间关联度能与X2之间建立一种关系,也反映了连接度存在一个阈值,超过这一阈值二者之间的显著性就迅速提高。通过散点拟合,发现各种关联度(包括CI)均与X2形成二次抛物线关系,X2显著性起点3.841至最大处,均反映了关联度或贡献度的显著性。种间还存在一种结构,用连接度或关联度来度量,反映了种间空间相遇概率的指标,客观反映种间关系。在这种种间关系达到一定的阈值时,二者就呈现显著性关系,这一阈值就是X2所规定的3.841以上,基本上就是抛物线扣除缓慢递变的底部而进入快速增长的中上部分,Y=3.841这条直线上方部分。

图1 钉螺与草种间关联度与其显著性(X2)回归拟合Fig.1 Regression and fi tting of correlation degree and signif i cant (X2) between oncomelania and vegetations
从上表可以看出,Pccc法与X2评价法有极高的相似性,说明两个指标评价结果的一致性。
3 结论与讨论
钉螺与共存草本种类连接性反映了动植物混合群落之间共栖与竞争用。山丘疫区钉螺分布生境狭窄沿水系连续性或泉眼间断性分布[14],且具水文异质性较强,生境稳定性差,人为干扰强度大等特征。钉螺具有水陆两栖特性,离不开湿地生境,在长期适生演化中与草本群落建立依存关系,钉螺与草本种群间连接性强弱反映钉螺和这些草本对生境要求的共性,同时也反映钉螺对这些湿生草本群落的依存度。本文用X2检测种间连接性的显著性,找到了与钉螺存在相互促进而共生的物种10种,如水芹Oenanthe javanica、水金凤Impatiens uliginosa、毛茛Ranunculus japonicus、拉拉藤Galium aparine var. tenerum、空心莲子草Alligator alternanthera、广布野豌豆Vicia cracca(极显著正连接),以及五朵云Euphorbia helioscopia、蜈蚣草Nephrolepi scordifolia、碎米荠Cardamine leucantha、葎草Humulus scandens(显著正连接);4种种类为相互抑制关系,如野蔷薇Rosa multif l ora、苔草Carex tristachya、金星蕨Parathelypteris glanduligera(极显著),以及凤尾蕨Pteris multif i da(显著)。

表4 不同评价指数与X2统计模型Table 4 Relationship of different evaluation indexes of oncomelania community and X2 fitted models
山丘型钉螺栖息地可划分为三类,一类是极度适生生境(Excellent habitat),能长期繁衍,一类是一般性适生生境(f i tness habitat),为迁移性或退化性栖息地,能短期内繁衍,一类是不适生性生境(Unf i t habitat),为水文驱动迁移而不适生生境,不能满足繁衍需求。从群落选择上看,极度适生生境能选择出正连接的物种种类,而一般性生境次之,不适生生境则为干扰因子。水芹、水金凤、毛茛、拉拉藤、空心莲子草、广布野豌豆,以及五朵云、蜈蚣草、碎米荠、葎草等均为亚热带山丘区沟道和水岸水陆交错带上永久或临时性湿地上的常见种类,通过大范围调查与统计检验,获得了显著性的钉螺伴生种类,能指示钉螺的存在;与指示植物相反,一些种类与钉螺形成相互抑制的关系,即钉螺与这些物种之间存在极显著的不相遇关系,在空间上完全隔离,如野蔷薇、苔草、金星蕨,以及凤尾蕨等种类,此类植物出现则也表示钉螺不存在,这些负连接种类也构成了指示植物的内容。
从钉螺适生群落环境整体上考虑,群落、土壤因子与钉螺密度之间相关性分析也表明了适宜的土壤水分、土地利用类型,以及土壤温度对密度的促进作用(如表2所示)。按林地、荒坡、沟渠、旱地、河滩和水田等6类进行分类统计,水田、荒地和河滩的活螺出现率与钉螺密度相对较高,沟渠和旱地次之,林地最低[8,10]。土地利用方式宏观上决定了水分、热量、肥力状况,进而对物种种类构成了基础性的选择压力。人为土地利用景观背景下,钉螺主要栖息于坡坎下排水沟、引水沟或沟渠;水田、荒地、旱地坡坎与河滩都满足草本生存适宜的水文条件,沟渠与坡坎类似,深度较大,林地则直接从内部和外部均质化了非生物景观,无法满足钉螺栖息的水热适宜富集性的条件。这种生物群落种间连接离不开群体环境的协作,或者建立在群体环境基础上。正指示植物与钉螺连接性也建立在对群体生物与非生物生境的适应性基础上,即非生物土壤因子是指示植物与钉螺共有生境,构成了同资源种团。
OI、JI、DI实际上是等效的,他们都不受均未出现的样方数(d值)的影响,与连接系数、Pccc不同[15]。OI、JI、DI关联度忽略了d值的存在,值越大,则正连接越紧密,值越小,负连接越显著,能反映X2检验的连接的不显著的连接性;X2无正负之分,分析简单,有明确的指标(3.841与6.667作为P0.05与P0.01的界限),能比较准确客观地表现种间连接性[12]。通过 OI、JI、DI、Pccc,以及CI,与X2建立线性关系,则发现均符合抛物线函数关系,且形成了一致的种间关联度变化趋势线,反映了种间关系形成的过程,即经历了缓慢的积累和适应过程,达到3.841 X2值后,便分化出正连接或负连接关系,要么形成紧密结合的共享生态位特性,要么形成相互分离而不相干的生态位分离。OI、JI、DI、Pccc是经过检验的种间关联度分析方法,而CI则是依照钉螺自身的畏强光特性需要群落盖度遮阴这一特点而建立的指数,也与其他关联指数一样,与X2得到抛物线函数关系,也可用盖度变化与钉螺分布密度的联系来阐明两因子之间连接的紧密性,即非生物因子也可作为种类或类似随机性程度不等的因子进行关联度分析。这一点与钉螺适生草本群落盖度在60%~100%[16-19],经历低盖度积累到高盖度而形成紧密相关的影响钉螺分布概率的参数。
钉螺肉眼难以识别,但与钉螺相伴生的指示植物,那些正连接的种类,能协助找寻钉螺,如看到这些指示植物,就会认为,这些生境容易栖息钉螺,或者为钉螺栖息的潜在生境。钉螺指示植物也能协助判定或评价血防林抑螺效果。传统药物灭螺或物理灭螺均未从栖息生境上彻底改变钉螺栖息的生态位;仅仅是临时性灭螺,土壤深层钉螺无法杀灭,且草丛盖度60%以上,喷施药物多被草丛截留。结合钉螺与群落、土壤因子,以及种间连接、贡献度问题,灭螺需要从钉螺生境彻底改造入手,简单的表层改变,如物理化学灭螺已经不能满足需要了,不具有生态性、高效性、稳定性、可持续性。林业血防工程则通过改造环境,通过生态系统过程持续的发挥抑螺防病功能。血防工程评价标准长期以钉螺密度和感染率作为指标,评价结果具短期性和临时性,需要研发新的评价指标来,使评价更符合实际和科学性。
钉螺与指示植物关系的建立为新标准的建立提供了方向和理论基础。指示植物作为血防林效果评价指标具有严密的科学性,值得在生产中细化研究,以获得更为细致的评价指标体系,来取代钉螺密度评价指标。血防林通过水分消耗、光照滤过和地表枯落替代来改造钉螺适生生境,使得原有的指示植物因水分改变或光照改变而无法定居,要么湿地植物消失,要么喜光植物消失。最终指示植物被非指示植物替代,钉螺则更为敏感和脆弱,使钉螺生境破碎化、或完全补破坏,因此指示植物的存在与否,是钉螺随着指示植物而不适生,形成易于辨认的钉螺血防林功能评价指标。以往种间关系研究多在植物种间进行,而本文是以钉螺为核心,在钉螺与草本之间进行,这是生境指示性研究的一种新的探索,值得深化完善。
[1] 蒋俊明,辜建军,高贵东,等.山丘型抑螺防病林的功能分析[J].湿地科学与管理,2010,6(2):4-8.
[2] 郭玉红,郎南军,杨文灿,等.云南高原山地林分郁闭度与钉螺密度的关系[J].湿地科学与管理,2011,7(1):14-15.
[3] 钟花相,董兴齐.基于遥感监测山丘型钉螺分布的研究进展[J].疾病预防控制通报,2008,23(6):77-79.
[4] 秦建新,谭子芳,张 春.洞庭湖区江滩钉螺分布的环境因子及空间特征——基于RS/GIS的研究[J].自然灾害学报,2008,(14):19-27.
[5] 郭家钢,林丹丹,胡广汉,等..应用遥感技术快速确定鄱阳湖区钉螺孳生地的研究[J].中华流行病学杂志,2002,(2):99-101.
[6] 张治英,孙志东,徐德忠,等.应用地理信息系统对江宁县钉螺孳生地环境特征的分析[J].疾病控制杂志,2004,(1):16-18.
[7] 周晓农,孙乐平,洪青标,等.地理信息系统应用于血吸虫病的监测Ⅲ.长江洲滩钉螺孳生地的监测[J].中国血吸虫病防治杂志,1999,(4):199-201.
[8] 张旭东,漆良华,黄玲玲,等.山丘区土壤环境因子对钉螺(Oncomelania snail)分布的影响[J].生态学报,2007,27(6):2460-2467.
[9] 黄玲玲.山丘区钉螺分布特征及其与环境因子的关系[D].北京:中国林业科学研究院,2006.
[10] 蒋俊明,何亚平,费世民,等.山丘型地区钉螺孽生数量与植被和土壤环境因子的关系[J].湿地科学与管理, 2006,2(4):33-39.
[11] 许发森,钱晓洪,文 松,等.安宁河流域植物、土壤特征与钉螺分布的关系[J].四川动物, 1999, 18(2):62-63.
[12] 张倩媚,陈北光,周国逸.鼎湖山主要林型优势树种种间连接性的计算方法研究[J].华南农业大学学报, 2006,27(1):79-83.
[13] 张任好.檫树群落主要树种种间连接性研究[J].林业科技开发,2006,20(4):24-26.
[14] 江泽慧.兴林灭螺文集[M].北京:中国林业出版社,1995.
[15] 王伯荪,彭少麟.南亚热带绿叶林种间连接测定技术研究Ⅰ:种间连接测定的探讨与修正[J].植物生态与地植物学丛刊,1985, 9(4):274-285.
[16] 吴 刚,苏瑞平,张旭东. 长江下游滩地植被与钉螺孽生关系的研究[J].生态学报,1999,19(1):118-121.
[17] 张旭东,杨晓春,彭镇华.钉螺分布与滩地环境因子的关系[J].生态学报,1999,19(2):265-269.
[18] 申瀚文,鄢前飞,曾思齐,等.木荷次生林种内和种间竞争研究[J].中南林业科技大学学报,2012,32(4):81-86.
[19] 谭一波,詹潮安,杨海东,等.广东南澳岛华润楠群落主要树种种间联结性[J].中南林业科技大学学报,2012,32(11):92-99.
Research on indicator plants of oncomelania habitats in Sichuan basin
JIANG Jun-ming1,2, ZHOU Jin-xing1, FEI Shi-min2, LIU Guo-hua2
(1. Chinese Academy of Forestry, Beijing 100091, China; 2.Sichuan Academy of Forestry, Chengdu 610081, Sichuan, China)
Based on the survey data of oncomelania density and herb layer characteristics at 157 quadrats (snail endemic area) located in Renshou country of Sichuan province, the indicator plants of snail habitat and interspecies association degree with the oncomelania habitats were investigated by using the methods such as variance analysis, X2points statistical measures, Ocihiai, Dice, Jaccaed, Pccc,etc. The results show that in the 85 herbaceous plant communities, 6 species (Oenanthe javanica, Impatiens uliginosa, Ranunculus japonicus., Galium aparine var. tenerum, Alligator alternanthera, Vicia cracca) , had a very signif i cant positive association with snail,4 species (Rosa multif l ora, Carex tristachya, Parathelypteris glanduligera and Pteris multif i da had a negative association with snail;3 species (Rosa multif l ora, sedge, marsh fern) had a highly signif i cant negative correlation with oncomelania, Pteris majestica had a negative correlation; but 71 kinds of species had no signif i cant association to oncomelania. Pearson relation analyses indicate that the content of soil water and land use types were two key factors of determining the snail density and distribution. After analysis, it was found that Ocihiai, Dice, Jaccaed, Pccc for association index among species reached the level of positive and negative association while these indexes reached a certain threshold, and these processes accorded with a parabolic relation between four indexes and X2. Snail habitats were classif i ed into three kinds: extreme adaptation, common adaptation and no adaptation, the last was the consequence that the extreme habitat degraded, reestablished after snail-killed forestation. The reconstructed effects of the forest for schistosome prevention could be described by using the no adaptation to snail, and its symbol was that the positive species disappeared and the negative association species occurred. Thus, the snail index species could be used to distinguish different snail habitats, also to evaluated the effects of mode forest for forestation and schistosome prevention.
oncomelania; oncomelania habitats; indicator plant; hilly infected area; evaluation indexes; for snail control and schistosomiasis prevention forest
S718.53;Q948
A
1673-923X(2013)07-0014-06
2013-02-15
国家“十二五”攻关项目“高原山地林业血防生态安全体系构建技术研究与示范”(2011BAD38B07-01);国家自然基金项目“滩地林业血防生态工程抑螺防病机理研究”(41071334)
蒋俊明(1966-),男,四川仁寿人,副研究员,主要从事森林生态方面的研究,Junming621@sina.com
E-mail:zjx9277@126.com
[本文编校:吴 毅]
猜你喜欢
热带病与寄生虫学(2022年4期)2022-09-20
热带病与寄生虫学(2022年4期)2022-09-20
热带病与寄生虫学(2022年4期)2022-09-20
河南师范大学学报(自然科学版)(2022年5期)2022-08-08
中国农业科学(2022年10期)2022-06-28
华人时刊(2022年7期)2022-06-05
华人时刊(2022年4期)2022-04-14
广东蚕业(2021年12期)2022-01-18
湖北大学学报(自然科学版)(2021年1期)2021-01-06
蔬菜(2018年5期)2018-05-17
