
基于树干液流技术的北京市刺槐冠层吸收臭氧特征研究
2013-12-20 03:06欧阳志云任玉芬张红星王效科郜世奇关云飞高付元
生态学报 2013年23期
王 华,欧阳志云,任玉芬,张红星,王效科,郜世奇,关云飞,高付元
(1.中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085;2.北京市农林科学院林业果树研究所,北京 100093;3.北京教学植物园,北京 100061)
近地层臭氧(O3)作为影响全球气候变化的重要因素和危害人类健康、动植物生长的二次污染物,受到广泛关注[1-3]。随着城市化进程的推进,近地层臭氧浓度迅速增加[4]。树木是城市生态系统的重要组成部分,可通过气孔途径和非气孔途径有效去除大气臭氧,在净化空气方面发挥着重要作用[5]。其中,通过气孔途径吸收是林木去除臭氧的主要途径,并与臭氧引起林木伤害密切相关[6]。因此,城市树木吸收臭氧方面的研究已成为目前植物生理生态学和城市生态学研究的热点。然而,现有的大部分研究主要集中在天然林吸收臭氧的影响机制[7-10],城市树木吸收臭氧的环境影响机制研究较少。
叶片尺度和生态系统尺度上森林去除臭氧的研究均有报道,但是冠层尺度上森林去除臭氧的研究很少。树干液流技术已经广泛应用于森林水分利用研究,由于水分蒸腾和吸收臭氧通过气孔行为耦合,故基于树干液流技术能够衡量森林冠层吸收臭氧量,这种方法考虑了边界层的影响,适用于异质性景观和山地景观[8,11]。因此,结合城市环境的异质化特征,本研究采用基于树干液流的方法衡量北京市典型绿化树种刺槐冠层吸收臭氧规律。
我国首都北京是世界上面积最大、历史最悠久的城市之一,是现代城市化的典型代表。由于大范围建设和机动车数量迅速增加,臭氧污染问题日趋严峻[12]。改善空气质量一直是该城市面临的重大难题。本文以北京市典型绿化树种刺槐为研究对象,通过测定树干液流密度,结合环境因子监测,探索冠层尺度上刺槐吸收臭氧特征及其与环境因子之间的关系。研究结果和方法可为明确环境如何影响树木吸收臭氧功能、空气质量提高、以及城市规划提供科学依据。
1 材料与方法
1.1 研究地点和研究对象
研究地点位于北京市南二环的北京教学植物园(116°25'37″— 116°25'50″E,39°52'20″— 39°52'28″N),占地面积116,500m2,周边商业区和住宅区密集,行人和机动车流量较大。该地区属于温带半湿润大陆性季风气候,年均气温11—12℃,年均降水量约为586 mm,超过70%发生在6—8月。
研究对象为北京市典型绿化树种刺槐,选择健康、生长环境一致的单株作为研究对象,2009年样树树形测定结果为:平均胸径为(38.43±2.43)cm,平均树高(12.87±0.20)m,平均边材面积为(129.12±12.11)cm2(表 1)。
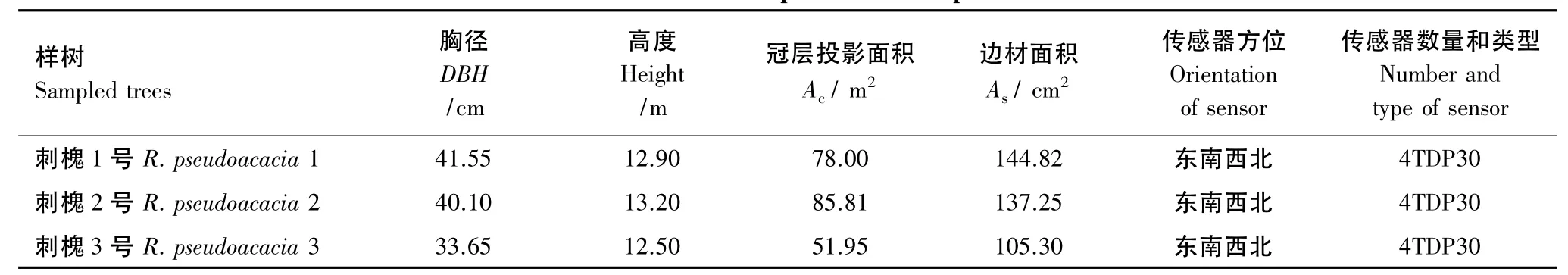
表1 测定树干液流样树的树形特征Table 1 Characteristics of the sampled trees for sap flow measurements
1.2 树形测定
树冠垂直投影面积采用测定东西和南北冠幅,结合椭圆面积计算公式确定。为避免伤害样树,于北京教学植物园和北京林业大学鹫峰教学林场选取各个径级的23株树木,采用生长锥钻取木芯,再用直尺测定边材厚度,建立边材面积与胸径之间的关系式,进而反推测定样树边材面积(图1)。
1.3 树干液流密度测定
2009年7月1日到2009年10月31日采用热消散传感器(Dynamax,Houston,TX,USA)连续测定单位边材面积上的瞬时树干液流密度(Js,g H2O cm-2s-1)。传感器的工作原理、安装、液流密度计算以及整树蒸腾计算方法参见文献[13-14]。将传感器馈线与数据采集器(CR1000 Dynamax,USA)连接,组成完整的树干液流测定系统,每10 min进行平均并储存数据,将笔记本电脑与数据采集器连接,定期采集数据。
在树干液流密度点的测定值尺度推广到整树尺度的过程中,忽视树干液流密度的轴向和径向变异,将引起结果的误差[15-16]。为明确树干液流密度的轴向变异,Herbst等建议小树采用1个或者2个传感器,大树采用4个传感器[17],因而,在每株刺槐样树东南西北面各安装一个传感器。由于刺槐样树边材厚度为2 cm左右,故树干液流密度径向变异不明显,因而,采用长度为30mm的传感器。
1.4 环境因素监测
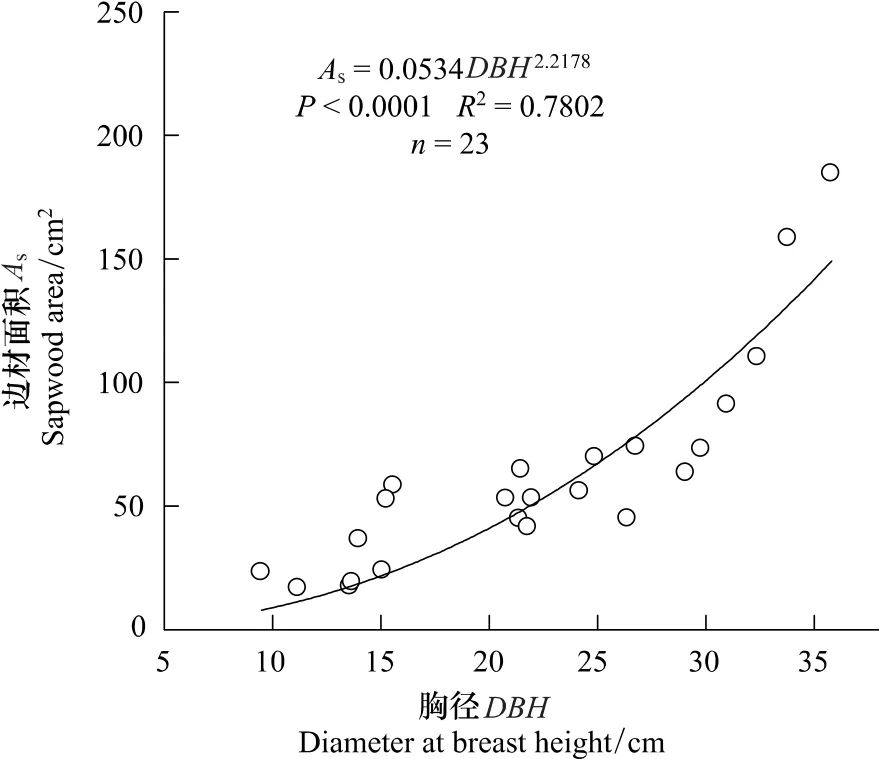
图1 刺槐边材面积与胸径之间的关系Fig.1 The relationship between sapwood area(A s)and diameter at breast height(DBH)for R.pseudoacacia
气象站位于北京教学植物园中较为开阔的一块空地上,避免树木、建筑物和其它障碍物的影响,同时距离研究对象近。空气温度湿度传感器(HMP45C,Vaisala Inc.,Helsinki,Finland),风速风向传感器(034B,Met One Instruments,Grants Pass,Oregon,USA)安装在高度为10 m的标准桅杆上。日照强度计(CMP-11,Kipp and Zonen,Delft,Netherlands)安装在高度为1.5 m的标准桅杆上测定太阳辐射。降雨由雨量计(TE525MM,Campbell Scientific Inc.,Logan,UT)在地面以上2 m以下高度测定。土壤温度传感器(109,Campbell Scientific Inc.,Logan,UT)埋设深度为10、30、50 cm和 80 cm。土壤含水量传感器(ECH2O,Decagon Devices Inc.,Pullman,WA,USA)埋设位于研究树木之间,深度为30 cm。水汽压亏缺是由大气温度和空气相对湿度计算得到[18]。上述气象和土壤参数监测与树干液流测定同步。
大气臭氧浓度采用 TEI Model 49i气体分析仪(Thermo Environmental Instruments Inc.,Franklin,MA,USA)监测。臭氧监测仪器距离研究对象近,每小时记录一个浓度值。
1.5 冠层导度计算
城市树木冠层开放,通风好,空气动力学粗糙,表明冠层表面与大气之间耦合度强[19]。因此,研究树种刺槐的整树冠层导度(Gc,mmol H2O·m-2·s-1)计算公式如下[20]:

式中,Ec是冠层蒸腾,将树干液流密度乘以边材面积再除以冠层投影面积得到[13];ρ是水的密度(998 kg/m3);GV是水蒸气的通用气体常数(0.462 m3kPa K-1kg-1);Ta是大气温度;D是水汽压亏缺(kPa)。参考Granier等人的建议[21],基于如下原则选择树干液流密度数据计算冠层导度:1)去除降雨或者降雨以后2h的数据,从而避免蒸散和树木蒸腾之间不一致;2)去除总辐射、水汽压亏缺、或者林段蒸腾非常低的数据(小于最大值的5%),因为在这些条件下计算冠层导度的不确定性较大。
Gc对水汽压亏缺(D)和总辐射(Rs)的响应规律通过边界线分析得到[22-23],采用不同D(步长0.2 kPa)或者Rs(步长50 W/m2)等级下3株样树的最大冠层导度。采用如下公式描述Gc响应D变化[19]:

式中,参数-m为回归方程斜率,量化了气孔导度响应D变化的敏感度;参数b为回归方程截距,为D=1 kPa的参比冠层导度。采用Sigma-Plot 10.0软件(Systat Software Inc.,San Jose,California)的最小二乘回归确定m和b。上述Gc和ln(D)之间的关系式为各样树之间的比较提供了简便的基准。
1.6 整树冠层吸收臭氧速率计算
叶肉中臭氧浓度近乎为0[24],因此,可以根据如下通量公式计算冠层吸收臭氧速率:

式中,FO3是单位冠层投影面积的冠层吸收臭氧速率,[O3]是大气臭氧浓度,Gc是冠层对水蒸气的导度,0.613是转换系数,代表大气中臭氧和水蒸气扩散系数比[7,25]。
2 结果
2.1 刺槐吸收臭氧的日变化规律
夏季晴天天气条件下,大气臭氧浓度随着总辐射增大逐渐上升,于15:00左右达到峰值,之后开始下降;刺槐冠层对臭氧的导度上午达到峰值,之后迅速下降;刺槐吸收臭氧速率呈单峰曲线,于15:00左右达到最大值,峰值范围较窄,尽管上午冠层对臭氧的导度最高,但由于臭氧浓度很低,故上午刺槐吸收臭氧速率并没有达到最大值;刺槐吸收臭氧累积量不断增加,中午前后增加最明显,夜间随着臭氧浓度降低和气孔关闭,其吸收臭氧累积量变化不大(图2)。
秋季晴天天气条件下,总辐射和水汽压亏缺明显减少,进而大气臭氧浓度也降低;刺槐冠层对臭氧的导度明显降低,且下午刺槐冠层对臭氧的导度响应水汽压亏缺下降而减少的幅度较小;刺槐吸收臭氧速率呈单峰曲线,峰值范围较宽;刺槐冠层吸收臭氧累积量明显减少,中午前后增加最明显,且从傍晚开始保持较为稳定的状态(图2)。
2.2 刺槐吸收臭氧的季节变化规律
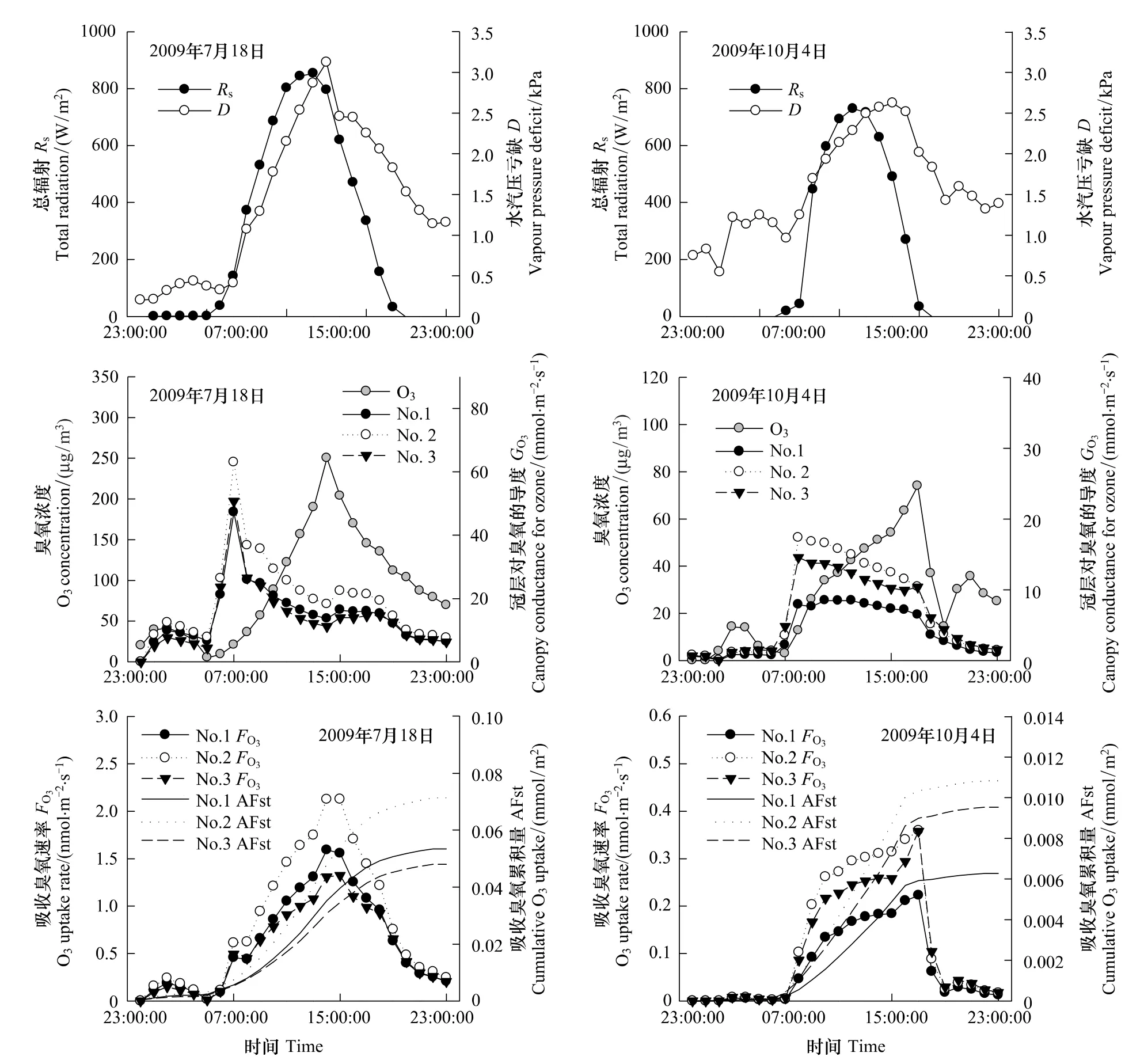
图2 夏(2009年7月18日)秋(2009年10月4日)季节晴天天气下总辐射(R s)、水汽压亏缺(D)、大气臭氧浓度(O3)、刺槐冠层对臭氧的导度(G O3)、吸收臭氧速率的日变化(F O3)以及吸收臭氧累积量(AFst)Fig.2 Diurnal course of total radiation(R s),vapour pressure deficit(D);ambient O3 concentration(middle)and canopy conductance for ozone(G O3);canopy O3 uptake rate(F O3)and cumulative O 3 uptake(AFst)in R.pseudoacacia on clear days in the summer(18th July 2009,left)and the autumn(4th October 2009,right)
大气臭氧浓度、刺槐冠层对臭氧的导度、吸收臭氧速率日均值以及吸收臭氧累积量的季节变化规律如图3所示。臭氧浓度日均值季节变化明显,夏季出现最大值146.4μg/m3,秋季出现最小值1.5μg/m3。夏秋季节刺槐冠层对臭氧的导度和吸收臭氧速率日均值波动较大,阴雨天气条件下冠层对臭氧的导度和吸收臭氧速率较低,而晴天天气条件下冠层对臭氧的导度和吸收臭氧速率较高;然而,从总体趋势上看,刺槐冠层对臭氧的导度和吸收臭氧速率从夏季到秋季逐渐降低,可见,刺槐吸收臭氧速率的日变化与季节变化规律交叠。刺槐吸收臭氧累积量夏季显著增加,秋季略有增加。
2.3 总辐射和水汽压亏缺对刺槐吸收臭氧的影响
刺槐冠层吸收臭氧速率日变化规律与大气臭氧浓度日变化规律相似(图2)。在一定的臭氧浓度下,刺槐冠层吸收臭氧速率的变化主要由冠层对臭氧的导度调控,进而受水汽压亏缺和总辐射影响(图4)。3株刺槐样树冠层对臭氧的导度接近,且对水汽压亏缺和总辐射的响应方式相似(图4)。
水汽压亏缺较高的条件下,臭氧浓度较高,但刺槐冠层对臭氧的导度降低,进而导致冠层吸收臭氧速率降低;水汽压亏缺较低的条件下,刺槐冠层对臭氧的导度很高,故在臭氧浓度较低的条件下,冠层吸收臭氧速率较高;水汽压亏缺很小的条件下,刺槐冠层吸收臭氧速率略降,可能是由于早晨光合作用较弱,气孔开度较小(图4)。
随着总辐射上升,臭氧浓度增加,当总辐射大于800 W/m2,臭氧浓度显著降低。当总辐射小于600 W/m2,刺槐冠层对臭氧的导度变化很小;当总辐射大于600 W/m2,刺槐冠层对臭氧的导度迅速下降,这个时候气孔可能达到了光饱和。因此,受臭氧浓度和冠层对臭氧的导度的双重影响,刺槐冠层吸收臭氧速率呈不对称单峰格局:当总辐射小于800 W/m2,随总辐射增强,刺槐冠层吸收臭氧速率略有增加;当总辐射在800 W/m2左右时,冠层吸收臭氧速率达到峰值,之后迅速下降(图4)。
3 讨论
3.1 刺槐整树冠层吸收臭氧特征
北京市典型绿化树种刺槐整树冠层吸收臭氧速率夏季高于秋季,与夏季大气温度、辐射、臭氧浓度等较高有关,这与高海拔地区成年挪威云杉(Picea abies)、瑞士五叶松(Pinus cembra)和欧洲落叶松(Larix decidua)吸收臭氧速率的季节变化规律一致[8]。与上述研究不一致,加那利群岛松林吸收臭氧速率季节变化不大,可能是如下原因导致:首先,当空气和土壤干旱时,在浓雾对空气湿度的影响和松林根系能获取土壤深层水分的联合作用下,松林吸收臭氧速率降低不显著[26,27]。其次,由于地处低纬度和冷洋流中,该群岛气候季节变化不大。最后,群岛的经济长期以农业为主,大城市很少,故污染源少。
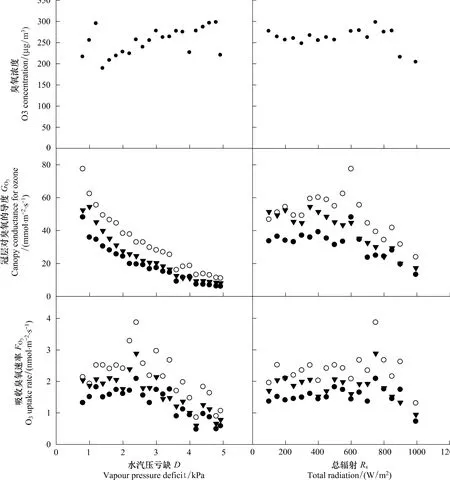
图4 2009年7月1日到2009年10月31日大气臭氧浓度(O3)、刺槐冠层对臭氧的导度(G O3)、吸收臭氧速率(F O3)对水汽压亏缺(D)和总辐射(R s)的响应规律Fig.4 Means of ambient air ozone concentration(O3),canopy conductance for ozone(G O3),and canopy O3 uptake rate(F O3)in relation to vapour pressure deficit(D)and total radiation(R s)from 1st July,2009 to 31th October,2009
北京市典型绿化树种刺槐单位冠层投影面积上年吸收臭氧量约为0.16 g/m2(例如,3株样树单位冠层投影面积上月均吸收臭氧量0.57 mmol·m-2·月-1×6个月×48 g/mol÷1000=0.16 g/m2),明显低于模型得到的结果[5,28]。采用城市森林影响模型,Nowak和 Dwyer[5]总结美国城市森林年均去除污染物量为 9.3 g/m2,变化范围为6.6—12.0 g/m2,臭氧占污染物比率为31.8%,因此其去除臭氧量约为2.96 g/m2(例如,9.3 g/m2×31.8%=2.96 g/m2)。采用同样的模型,北京城市森林去除污染物量较高(27.5 g/m2),臭氧占污染物比率为20%,因此其去除臭氧量约为5.50 g/m2[28]。野外观测数据得到的结果和模型方法得到结果区别较大,可能是如下原因导致。首先,基于树干液流的方法确定的是气孔吸收臭氧量,而模型方法确定的是干沉降值,包括气孔吸收量和树木非蒸腾表面吸附量。其次,模型方法得到的结果为森林去除污染物量的粗略估计,需要在如下几个方面调整参数以提高模型的准确度:模型产生结果的不确定性分析;测定树木个体去除污染物以改进模型的假设-树木处于相同层次;准确模拟大气污染物浓度及其与地点、城市森林的面积和树种组成之间的关系[28]。再次,老树较低的水力导度限制其光合速率[29],研究对象刺槐树龄超过100 a,因而推断其气孔导度明显降低,进而导致冠层吸收臭氧速率较低。最后,近年来北京市空气质量逐渐改善,大气臭氧浓度有所降低。两种方法得到结果的区别表明:评估森林受臭氧危害的风险应考虑各树种冠层臭氧通量。
3.2 刺槐整树冠层吸收臭氧的环境调控
城市典型绿化树种刺槐整树冠层吸收臭氧特征受臭氧浓度和冠层导度的共同影响。本研究地点臭氧浓度季节变化显著,夏季较高,秋季较低。在一定程度上,这导致了研究树种刺槐冠层吸收臭氧速率的季节变化规律与臭氧浓度的变化规律相似(图3)。在一定的臭氧浓度下,气孔导度是整树吸收臭氧速率的关键限制因子,进而受微气候、树龄、海拔等因素影响[7,25]。与上述研究一致,刺槐整树冠层对臭氧的导度主要受水汽压亏缺和总辐射影响(图4)。低辐射条件下刺槐保持一定的吸收臭氧速率,可能由于夜间、凌晨或者阴雨天气条件下气孔保持部分开放。很多研究表明某些植物夜间气孔开放,保持一定速率的蒸腾[30]。城市环境中较高的空气温度与较低的平均风速,可能诱导植物气孔增大开度。因此,城市环境下刺槐吸收臭氧速率可能高于自然环境下刺槐吸收臭氧速率。
3.3 冠层尺度上城市树木吸收臭氧研究方法
量化树木吸收臭氧的方法包括叶片水平上的气体交换法和冠层尺度上的涡度相关法、模型、基于树干液流的方法。气体交换法测定叶片吸收臭氧量,但是忽视了边界层的影响[31]。涡度相关技术测定冠层臭氧沉降量,但是无法区分气孔吸收臭氧与树木表面吸附臭氧[32]。模型则需要根据当地环境以及植被的特点对模型参数化[33]。基于树干液流的方法考虑了边界层的影响,适用于异质性景观和山地景观[7]。
由于城市环境突出的异质性特征,如何选择适合的方法研究城市树木吸收臭氧的一个难点。目前,城市林木吸收臭氧速率主要采用整合植被、气象和大气臭氧浓度信息的模型模拟的方法[28]。此类研究能反映区域尺度上城市树木去除臭氧速率,但是不能反映树木直接吸收臭氧量,也无法进行种间比较。为解决上述问题,根据树木吸收臭氧和蒸腾作用通过气孔行为耦合,提出基于树干液流技术和大气臭氧浓度监测确定城市树木整树吸收臭氧速率方法,为衡量城市林木吸收臭氧及其它痕量气体,以及评价城市树木受臭氧危害的风险等提供了准确可行的方法。
[1]Karnosky D F,Skelly JM,Percy K E,Chappelka A H.Perspectives regarding50 years of research on effects of tropospheric ozone air pollution on USforests.Environmental Pollution,2007,147(3):489-506.
[2]Sitch S,Cox P M,Collins WJ,Huntingford C.Indirect radiative forcing of climate change through ozone effects on the land-carbon sink.Nature,2007,448(7155):791-794.
[3]Selin N E,Wu S,Nam K M,Reilly J M,Paltsev S,Prinn R G,Webster M D.Global health and economic impacts of future ozone pollution.Environmental Research Letters,2009,4(4):044014.
[4]Vingarzan R.A review of surface ozone background levels and trends.Atmospheric Environment,2004,38(21):3431-3442.
[5]Nowak D J,Dwyer JF.Understanding the benefits and costs of urban forest ecosystem ∥ Kuser J E,ed.Urban and Community Forestry in the Northeast.New York:Springer Science and Business Media,2007:25-46.
[6]Landolt W,Keller T.Uptake and effects of air pollutants on woody plants.Experientia,1985,41(3):301-309.
[7]Wieser G,Häsler R,Götz B,Koch W,Havranek WM.Role of climate,crown position,tree age and altitude in calculated ozone flux into needles of Picea abies and Pinus cembra:a synthesis.Environmental Pollution,2000,109(3):415-422.
[8]Wieser G,Matyssek R,Köstner B,Oberhuber W.Quantifying ozone uptake at the canopy level of spruce,pine and larch trees at the alpine timberline:an approach based on sap flow measurement.Environmental Pollution,2003,126(1):5-8.
[9]Nunn A J,Wieser G,Metzger U,Löw M,Wipfler P,Häberle K,Matyssek R.Exemplifying whole-plant ozone uptake in adult forest trees of contrasting species and site conditions.Environmental Pollution,2007,146(3):629-639.
[10]Braun S,Schindler C,Leuzinger S.Use of sap flow measurements tovalidatestomatal functions for mature beech(Fagussylvatica)in view of ozone uptake calculations.Environmental Pollution,2010,158(9):2954-2963.
[11]Köstner B,Matyssek R,Heilmeier H,Clausnitzer F,Nunn A J,Wieser G.Sap flow measurements as a basis for assessing trace-gas exchange of trees.Flora-Morphology,Distribution,Functional Ecology of Plants,2008,203(1):14-33.
[12]Chan C K,Yao X H.Air pollution in mega cities in China.Atmospheric Environment,2008,42(1):1-42.
[13]Granier A.Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements.Tree Physiology,1987,3(4):309-320.
[14]Zhao P,Rao X Q,Ma L,Cai X A,Zeng X P.Application of Granier’s sap flow system in water use of Acacia mangium forest.Journal of Tropical and Subtropical Botany,2005,13(6):457-468.
[15]Delzon S,Sartore M,Granier A,Loustau D.Radial profiles of sap flow with increasing tree size in maritime pine.Tree Physiology,2004,24(11):1285-1293.
[16]Lu P,Müller W J,Chacko E K.Spatial variations in xylem sap flux density in the trunk of orchard-grown,mature mango trees under changing soil water conditions.Tree Physiology,2000,20(10):683-692.
[17]Herbst M,Roberts JM,Rosier P T W,Taylor M E,Gowing DJ.Edge effects and forest water use:a field study in a mixed deciduous woodland.Forest Ecology and Management,2007,250(3):176-186.
[18]Campbell G S,Norman JM.An Introduction to Environmental Biophysics.2nd ed.New York:Springer-Verlag,1998.
[19]Oren R,Sperry JS,Katul G G,Pataki D E,Ewers B E,Phillips N,Schäfer K V R.Survey and synthesis of intra-and interspecific variation in stomatal sensitivity to vapour pressure deficit.Plant,Cell and Environment,1999,22(12):1515-1526.
[20]Köstner B M M,Schulze E D,Kelliher F M,Hollinger D Y,Byers J N,Hunt J E,McSeveny T M,Meserth R,Weir P L.Transpiration and canopy conductance in a pristine broad-leaved forest of Nothofagus:an analysis of xylem sap flow and eddy correlation measurements.Oecologia,1992,91(3):350-359.
[21]Granier A,Loustau D,Bréda N.A generic model of forest canopy conductance dependent on climate,soil water availability and leaf area index.Annals of Forest Science,2000,57(8):755-765.
[22]Jarvis P G.The interpretation of the variations in leaf water potential and stomatal conductance found in canopies in the field.Philosophical Transactions of the Royal Society B:Biological Sciences,1976,273(927):593-610.
[23]Dye P J,Olbrich B W.Estimating transpiration from 6-year-old Eucalyptus grandis trees:development of a canopy conductance model and comparison with independent sap flux measurements.Plant,Cell and Environment,1993,16(1):45-53.
[24]Laisk A,Kull O,Moldau H.Ozone concentration in leaf intercellular air spaces is close to zero.Plant Physiology,1989,90(3):1163-1167.
[25]Wieser G,Havranek WM.Environmental control of ozone uptake in Larix decidua Mill.:a comparison between different altitudes.Tree Physiology,1995,15(4):253-258.
[26]Luis V C,Jiménez M S,Morales D,Kucera J,Wieser G.Canopy transpiration of a Canary Islands pine forest.Agricultural and Forest Meteorology,2005,135(1/4):117-123.
[27]Wieser G,Luis V C,Cuevas E.Quantification of ozone uptake at the stand level in a Pinus canariensis forest in Tenerife,Canary Islands:An approach based on sap flow measurements.Environmental Pollution,2006,140(3):383-386.
[28]Yang J,McBride J,Zhou J X,Sun Z Y.The urban forest in Beijing and its role in air pollution reduction.Urban Forestry and Urban Greening,2005,3(2):65-78.
[29]Hubbard R M,Bond B J,Ryan M G.Evidence that hydraulic conductance limits photosynthesis in old Pinus ponderosa trees.Tree Physiology,1999,19(3):165-172.
[30]Daley M J,Phillips N G.Interspecific variation in nighttime transpiration and stomatal conductance in a mixed New England deciduous forest.Tree Physiology,2006,26(4):411-419.
[31]Wieser G,Havranek W M.Ozone uptake in the sun and shade crown of spruce:quantifying the physiological effects of ozone exposure.Trees,1993,7(4):227-232.
[32]Zeller K F,Nikolov N T.Quantifying simultaneous fluxes of ozone,carbon dioxide and water vapor above a subalpine forest ecosystem.Environmental Pollution,2000,107(1):1-20.
[33]Wieser G,Emberson L D.Evaluation of the stomatal conductance formulation in the EMEP ozone deposition model for Picea abies.Atmospheric Environment,2004,38(15):2339-2348.
参考文献:
[14]赵平,饶兴权,马玲,蔡锡安,曾小平.Granier树干液流测定系统在马占相思的水分利用研究中的应用.热带亚热带植物学报,2005,13(6):457-468.
猜你喜欢
作物学报(2022年9期)2022-07-18
农业工程学报(2022年3期)2022-04-16
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
科学大众(中学)(2019年3期)2019-05-17
环境保护与循环经济(2017年9期)2017-03-16
电源技术(2015年5期)2015-08-22
电源技术(2015年7期)2015-08-22
中国环境监测(2015年3期)2015-04-26
