
施肥梯度对高寒草甸群落结构、功能和土壤质量的影响
2013-12-16 08:18王长庭王根绪王启兰
生态学报 2013年10期
王长庭,王根绪,刘 伟,王启兰
(1.西南民族大学生命科学与技术学院,成都 610041;2.中国科学院水利部成都山地灾害与环境研究所,成都 610041;3.中国科学院西北高原生物研究所,西宁 810001)
长期的科学研究和实践证明,采用围栏封育、短期休牧等措施,是合理利用草地、防止草地退化的有效方法之一[1-3]。草地退化的原因十分复杂,但从生态系统学理论的观点来看,其本质上是由于草地生态系统中能量流动和物质循环失衡、入不敷出造成的。人类在利用草地时(如放牧及刈割等),土壤养分随着草产品及畜产品的输出被过量地带出草地,在没有得到有效补充的情况下,草地土壤肥力逐渐下降,甚至瘠薄,严重影响牧草的生长,草地生态系统严重退化。高寒草甸生态系统退化[4-6]不仅表现在物种数减少、多样性下降、能量的分配转向地下等;也表现在土壤肥力、土壤质量和微生物活性的降低等[7-9]。草地退化导致土壤有机碳减少[10-11]。随着放牧强度的增加,土壤有机碳亦呈明显下降趋势[12-13]。高寒草地的退化减少了植被生产力和土壤有机质输入量,并加快了土壤有机质分解速率,从而影响到生态系统的碳循环[8]。草地施肥是一种维持草地生态系统养分平衡的重要管理措施,草地施肥能够补充土壤营养物质,有助于草地生产力的恢复和提高,但也会减少群落物种数量,降低物种多样性[14-17]。草地生态系统的可持续性和草地生产力的维持在很大程度上依赖于草地植物群落的物种多样性。因此,无论是研究不同管理方式下土壤物质循环以维护土壤资源,还是为保护植物群落物种多样性以维系草地生态系统的可持续性发展,都需要人们对碳在各库之间的储量、草地生产力和稳定性的维持有清晰的认识,特别是对阐明土壤碳氮转化、评价土壤肥力、建立持续生产的施肥模式等均具有重要意义。
施氮肥可以增加土壤中的有效氮,刺激植物的生长,增加植物和土壤中有机碳的积累[18]。氮的输入(自然氮沉降和人为施氮)对于受到氮限制的陆地生态系统碳循环过程必然产生相应的影响,并影响其碳源汇状况[19-20]。大气氮沉降输入增加了高寒草甸生态系统氮素的可利用性,进而影响土壤以及整个生态系统的碳循环过程[21]。过多的氮输入也将使生态系统达到氮饱和状态,以致影响到许多生态系统过程,如:生态系统生产力、物种组成以及多样性等[22-26]。此外,当某个生态系统中氮积累到一定程度后,会出现氮饱和[27],也会减少碳储量[28]。然而,输入到生态系统中氮的最适宜量是多少、有多少能够被植物所吸收利用、氮在土壤中的分配情况如何,这些很大程度上决定着陆地生态系统中所能形成的碳汇大小。因此,本研究通过施氮梯度试验研究氮增加对高寒矮嵩草草甸群落结构、功能;土壤全量养分、速效养分;土壤有机碳和微生物生物量碳的影响,旨在揭示矮嵩草草甸群落特征;土壤养分和土壤微生物活性对氮素添加的响应,为高寒草甸的适应性管理提供科学依据。
1 材料与方法
本研究于2008—2009年8月在三江源腹地青海省果洛藏族自治州玛沁县进行。地理位置为34°30'—33°34'N,100°29'—99°54'E,平均海拔4 150 m,范围3800—4800 m。该地区气候具有典型的高原大陆性气候特点,无四季之分,仅有冷暖季之别,冷季漫长、干燥而寒冷,暖季短暂、湿润而凉爽。温度年差较小而日差较悬殊,太阳辐射强烈。日照充足,年日照平均值在2 500 h以上,年总辐射量在629.9—623.8 kJ/cm2。冷季持续时间长达7—8月;暖季湿润,长4—5月。年平均气温-1.7℃,1月平均气温-14.8℃,7月平均气温9.8℃。年平均降水量600 mm,主要降水量集中在5—9月,约占年降水量的80%,蒸发量1160.3 mm。土壤为高山草甸土和高山灌丛草甸土。高山嵩草草原化草甸、矮嵩草草甸为该地区主要的草地类型。建群种为小嵩草(Kobresia pygmaea)、矮嵩草(Kobresia humilis)。主要的伴生种有羊茅(Festuca ovina)、异针茅等禾草,杂类草有高山紫菀(Aster alpina)、湿生扁蕾(Gentianopsis paludosa)、高山唐松草(Thalictrum alpinum)、异叶米口袋(Gueldenstaedtis diversifolia)、黄帚槖吾(Ligularia virgaurea)等主要植物。
2006年在距果洛州大武镇约4 km的地方选择50 hm2的未退化矮嵩草草甸(冬春草场)用围栏保护。2008年5月下旬在围栏内选择面积为50 m×50 m的地势平坦样区,分成5个30 m×20 m的小样区,小样区间距为2 m,各个小样区四角用木桩标记。施肥时以单因素随机区组实验设计施肥。因为氮素是大多数陆地生态系统初级生产力的主要限制因子[29-30],并且根据以往在该地的长期试验资料[17,26,31-37]。施肥处理有5个水平,分别为 12、20、32、40、0 g/m2;所施肥料为尿素 CO(NH2)2(46.65%N)[17]。每个处理设置 3 个重复,共15个小区。从2008年5月下旬开始第1次施肥、每年5月下旬施肥1次。施肥在多云的阴天进行(因为在多云的阴天施肥,施肥当天或次日有降雨,这就保证了所施肥在短期内能溶于土壤中),用手将尿素颗粒均匀的洒在小样区。
2008、2009年8月下旬用样方法[38]在上述各个样地内对角线法设置10个1 m×1 m的观测样方进行植物群落调查,调查参数包括样方内所有植物种的分盖度、高度和频度,然后齐地面按禾本科、豆科、莎草科和杂类草4个功能群[38]剪草,在60℃烘箱烘干称重。
地下生物量用土钻取样[39],0—40 cm土层每10 cm取样,共4层,用内径5 cm土钻在每个样地取10钻(采用“V”字形法),清水冲洗得到根系样品,分别在60℃和105℃烘至恒重,称干重。
于2008、2009年8月在测定过地下生物量的样方(25 cm×25 cm),采用土壤剖面法分层采集土壤样品(0—10 cm、10—20 cm、20—40 cm),过筛(2 mm)保存于4℃冰箱中,用于微生物生物量、土壤酶活性的测定。另外,用土钻钻取10个样点(采用“V”字形法)混合为一个土壤样品,即0—40 cm土壤样品,6次重复,风干后测定土壤基本成分。土壤中的测试项目为土壤样品的全磷(钼锑抗比色法)、速效磷(碳酸氢钠浸提-钼锑抗比色法)、全N、硝态N和铵态N(凯氏法、比色法)以及有机质含量(丘林法)、全钾 (氢氧化钠碱熔-火焰光度法)、速效钾(乙酸铵浸提-火焰光度法)[40]。微生物生物量碳采用灭菌-提取法[41]。
生物量碳和土壤微生物熵的测定:

式中,鲜土样称10 g做熏蒸,后用50 mL抽提剂抽提熏蒸土样,再取1 mL抽提液稀释10倍测TC、IC含量)

统计分析
采用Microsoft Excel整理和计算数据,对不同氮素水平下土壤全量养分、速效养分含量进行单因素方差分析(Owo-way ANOVA);土壤有机碳、微生物量碳和植物群落盖度、生物量进行单因素方差分析(One-way ANOVA),用DMRT多重比较进行显著性检验,并用多元线性回归分析土壤全量养分、速效养分含量;土壤有机碳、微生物量碳含量等相关指标之间的关系,以上所有分析在SPSS 13.0、DPS 7.05中进行。
2 结果与分析
2.1 不同施肥梯度下矮嵩草草甸土壤资源的变化
在矮嵩草草甸中,随着施肥量的增加,土壤全氮、硝态氮、铵态氮、速效磷、全钾、速效钾和有机质含量均发生了显著的变化,而且全量养分和速效养分在施肥量为20 g/m2或32 g/m2时较高,施肥量增加到40 g/m2时土壤资源逐渐降低(表1)。与对照相比,施肥处理一定程度上提高了土壤养分,养分在土壤中的积累,使其有机质、全氮含量增加,土壤营养状况得到改善(表1)。
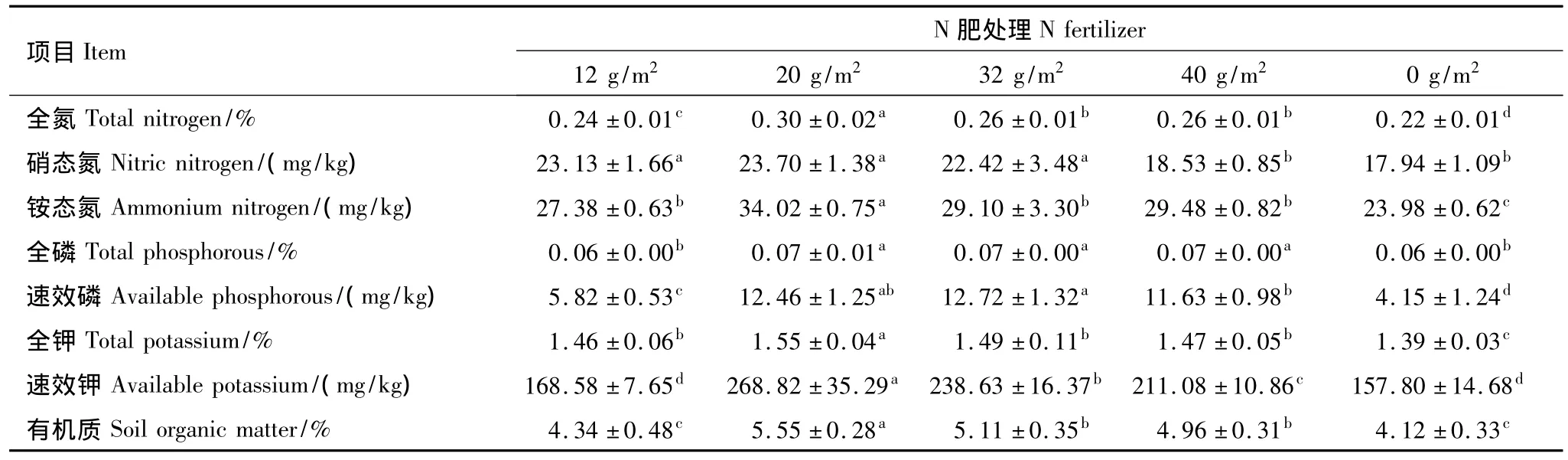
表1 高寒矮嵩草草甸群落的土壤资源Table 1 Soil resources(0—40 cm)of the K.humilis meadow community
2.2 各功能群盖度、生物量对施肥的响应
在施肥梯度上各功能群盖度和生物量的变化如表2。随着施肥量的增加,不同功能群的盖度响应是有差异的,禾本科植物的响应较大,而豆科和杂类草植物盖度明显降低,莎草科盖度变化不明显。施氮条件下对禾草类的影响最大,可明显使禾草类的盖度增加,但施肥量增加到一定程度,如施氮40 g/m2时,各功能群植物的盖度逐渐降低,具有一定的抑制作用。
从表2中可以看出:生物量随施肥梯度呈单峰曲线变化,不施肥时生物量最低,施肥20 g/m2或32 g/m2时生物量最高。随着施肥量的增加,植物功能群和群落生物量均有显著差异(表2)。禾本科、莎草科功能群和群落生物量显著降低,而豆科和杂类草功能群生物量显著增加(表2)。在0 g/m2到32 g/m2的施肥梯度间,植物功能群和群落生物量随施肥梯度提高而增加;在40 g/m2的肥力梯度上,植物功能群和群落生物量随施肥梯度提高而降低。

表2 不同施肥梯度矮嵩草草甸群落不同功能群植物生物量、盖度的变化Table 2 Changes in biomasses and cover of different plant groups in the K.humilis meadow community at different fertilization gradients
2.3 土壤有机碳、微生物量碳含量及微生物熵的变化
由表3可知,不同施肥梯度土壤有机碳含量具有一定的差异。随着施肥梯度的提高,土壤有机碳含量逐渐增加且呈单峰曲线变化。在0 g/m2到32 g/m2的施肥梯度间,土壤有机碳随施肥梯度提高而增加;在4 0 g/m2的肥力梯度上,随施肥梯度提高而降低。不同施肥梯度矮嵩草草甸土壤有机碳含量在0—10 cm土层明显较高,且随着施肥量的增加,分布在0—40 cm土层的土壤有机碳含量呈单峰曲线变化。施肥影响了土壤有机碳含量在土壤剖面上的分布(表3),即土壤有机碳含量随土层加深而减少,特别是在0—10 cm与10—20 cm、20—40 cm土层之间变化存在显著差异,施肥梯度显著地影响了土壤有机碳含量。

表3 高寒矮嵩草草甸群落土壤有机碳和微生物碳的分布特征(平均值)Table 3 Distribution of SOC and MBC in the K.humilis meadow community(Mean value)
不同施肥梯度土壤微生物生物量碳含量变化表现出与土壤有机碳含量变化类似的规律,随着施肥梯度的提高,土壤微生物生物量碳逐渐增加且呈单峰曲线变化。在0 g/m2到32 g/m2的施肥梯度间,土壤微生物生物量碳随施肥梯度提高而增加;在40 g/m2的肥力梯度上,随施肥梯度提高而降低。不同施肥梯度矮嵩草草甸土壤微生物生物量碳含量在0—10 cm土层明显较高(表3)。
不同土层的微生物熵因施肥梯度的不同而异,但其分布没有明显的规律性。只是随着施肥梯度,0—10 cm土层土壤微生物熵呈现下降趋势,但下降趋势相对于土壤有机碳和微生物量碳较平稳。
2.4 土壤有机碳、微生物量碳与土壤资源的相关性
土壤微生物生物量与土壤资源(图1,图2)尤其是土壤全量养分和速效养分有密切的关系。研究表明,高寒矮嵩草草甸随着施肥量的增加,植物群落功能群组成发生改变,群落组成的变化影响有机碳输入的数量和质量,进而影响有机碳在土壤中的积累,明显呈单峰曲线变化,而且随土壤全量养分和速效养分的变化而变化。施肥20 g/m2或32 g/m2时土壤有机碳和微生物量碳含量最高,反映出适宜施肥量的土壤微生物生态系统功能良好,反之土壤生态系统功能被抑制,土壤中微生物生物量降低,土壤微生物腐解能力减弱,土壤中营养元素循环速率和能量流动也减弱,导致高施肥量草地群落土壤质量低于适宜施肥量的草地。
3 讨论

图1 不同施肥梯度矮嵩草草甸土壤有机碳与土壤资源的关系Fig.1 Relationships between MBC and soil resources of the K.humilis meadow at different fertilization gradients
土壤理化性质包括土壤物理性质和化学性质,它是土壤质量的重要组成。土壤理化性质不仅影响土壤保持和供应水肥的能力,对调控土壤气热状况以及水分入渗性能和地表径流的产生也有重要作用[42]。草地生态系统功能的可持续利用最终必然要通过生态措施实现,通过物质循环形成自我维持系统。生态系统的物质循环是可持续发展的必要条件,施肥是提高草地生产力的主要途径[43-44]。从生态系统物质循环的角度出发,单纯改善土壤的物理性状,不能从根本上改善植物生长所需的营养元素,其效益难以持久。草地生态系统要想保持持续的良性循环,土壤养分的平衡是关键。适当地施肥是保证草地生态系统物质输入与输出间的平衡,实现系统持续生产的重要措施。一些研究表明,施氮肥提高生产力,就提高地上净生物量而言适合的施氮量是55.30—70.76 kg/hm2[45-46]。说明向自然生态系统中输入的氮素量的大小不仅影响土壤物质循环,而且对自然生态系统生产力、物种组成等群落结构和功能产生重大影响。另外一些研究表明,氮输入后土壤中可利用氮的数量增加,一方面会使得植物的碳同化作用增强,表现为地上植物生长加快,从而有更多的凋落物形成有机质进入土壤中;另一方面,由于有更多的氮素容易被植物获取,因而会使得通过根系分配到地下的碳的数量减少[47]。由此我们可以看出,氮输入会引起土壤中有机质两种不同的变化趋势。但通常情况下,高氮引起地上部分对土壤有机碳输入的增加会补偿高氮引起的地下碳分配的减少,最终表现为土壤中的有机质的增加[48]。本研究结果表明,施氮(低水平和高水平)均一定程度上提高了全量养分、速效养分含量,土壤营养状况得到改善,因为不同水平氮的输入:1)可能通过影响植物凋落物的产生以及植物碳向地下根的分配来影响土壤中有机质的数量及质量;2)增加的有机质将为土壤微生物提供更多的可利用的底物和能量,从而有利于微生物的活动;3)外源氮输入时,植物和微生物之间对氮的竞争可能减弱,微生物可利用氮素增加,使得微生物数量发生变化。4)外源氮输入导致的草地植物组成的改变通过影响进入土壤的植物组织的碳的质量和数量从而间接地影响微生物群落组成、微生物活性等。但施氮对土壤养分的影响并不是简单的线性正相关关系,可能存在一个响应阈值,超过这个阈值范围,施肥的作用就不明显了。
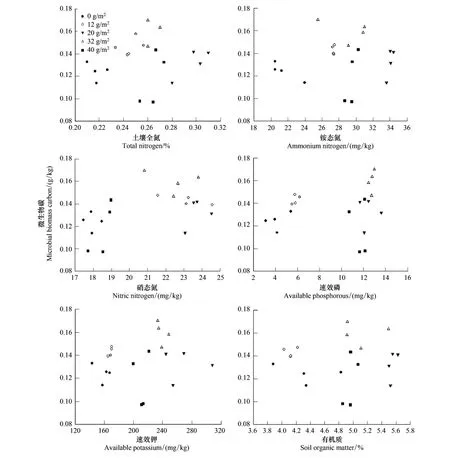
图2 不同施肥梯度矮嵩草草甸土壤微生物碳与土壤资源的关系Fig.2 Relationships between microbial biomass carbon and soil resources of the K.humilis meadow at different fertilization gradients
氮素添加通过增加土壤中NO-3-N等可利用资源的含量来改善土壤养分,显著增加了植物群落的地上生产力[26]。施用氮肥是补给土壤氮素和维持地上生产力的主要措施,是土壤氮素的主要来源[49]。Aerts等[50]发现草地群落中地上生物量与土壤的全氮含量呈正相关。氮素添加明显减少了成熟草原的物种丰富度[51]。随着施入不同资源组合数量的增加,物种多样性降低[37]。本研究表明,植物功能群和群落生物量随施肥梯度呈单峰曲线变化,而不是简单的线性正相关。在0 g/m2到32 g/m2的施肥梯度间,植物功能群和群落生物量随施肥梯度提高而增加;在40 g/m2的肥力梯度上,植物功能群和群落生物量随施肥梯度提高而降低。各植物功能群中,禾本科植物对氮素添加的响应极为显著,这可能是因为禾草具有较高的氮素利用率等自身属性,其地上部分迅速生长,高度和盖度增加,遮蔽了其他功能群中的矮小物种,从而在与其他各功能群物种光的竞争中取胜,减小了杂类草和豆科植物功能群的生物量和盖度。群落水平的自疏假说[34,52]认为施肥后群落中植物个体增大、总密度随施肥的增加而降低,而在特定面积的取样使得物种丰富度减小。总之,氮素添加通过改变植物群落的土壤养分含量等特性,改变了植物群落的物种、功能群组成,增加了禾本科植物个体大小,植物群落生产力增加,个体大小的增加导致密度降低,使群落中稀有种丧失。
微生物生物量作为土壤有机质的活性部分,是土壤养分的储存库和植物生长可利用养分的一个重要来源,并可作为土壤肥力水平的活指标[53]。草地生态系统功能的维持本质上是其能量流动和物质循环的平衡。人类在利用草地时(如放牧及刈割干草等),土壤养分随着草产品及畜产品的输出被过量地带出草地,在没有得到有效补充的情况下,草地土壤肥力逐渐下降,甚至瘠薄,严重影响草地生态系统的结构和功能。而草地施肥能补充土壤中所缺少的植物生长所必需的营养元素,提高了植物功能群、群落生物量,为土壤微生物提供丰富的能源物质。徐明岗等[54]对南方红壤丘陵区牧草的肥料效应与施肥研究确定了氮肥效应曲线为抛物线型,表明在其他养分供应一定时,氮肥并非施的越多越好,而是在一定量时为最佳,而且氮素需要每年持续施用才能高产。我们同样也发现,施肥20 g/m2或32 g/m2时土壤有机碳和微生物量碳含量最高,反映出适宜施肥量的土壤微生物生态系统功能良好,反之土壤生态系统功能被抑制,土壤中微生物生物量降低,土壤微生物腐解能力减弱,土壤中营养元素循环速率和能量流动也减弱,导致高施肥量草地群落土壤质量低于适宜施肥量的草地。
因此,相对于高寒草甸生态系统而言,其临界氮阈值是多少?合理的氮输入对我国草地生态系统多样性的保护和其生态功能的维持意义深远。根据以往的研究发现,青海省人工禾草草地施氮试验中其施肥量为150 kg/hm2[33]。青藏高原小嵩草草甸尿素的最佳施用时间应在6月下旬至7月上旬,较为经济合理的施肥量应在300 kg/hm2左右[55]。而德科加等[3]在干草原类草场获得最佳产草量及经济效益的施肥量是450 kg/hm2。本研究初步判定高寒草甸最佳施肥(氮肥)水平是30 g/m2。值得注意的是在研究氮输入对高寒草甸生态系统的影响时,应强调长期试验研究的重要性,以便降低研究结果的不确定性;临界氮阈值与草地生态系统的初始氮状况和植被、土壤特征关系。
4 结论
通过高寒草甸不同施肥梯度对高寒草甸群落结构、功能和土壤质量的影响,综合以上结果,可得到如下结论:
(1)外源氮(低水平和高水平)均一定程度上提高了全量养分、速效养分含量,土壤营养状况得到改善。但施氮对土壤养分的影响并不是简单的线性正相关关系,可能存在一个响应阈值,超过这个阈值范围,施肥的作用就不明显了。
(2)植物功能群盖度和生物量随施肥梯度呈单峰曲线变化。
(3)施肥水平在20—30 g/m2时土壤有机碳和微生物量碳含量最高,反映出适宜施肥量的土壤微生物生态系统功能良好,反之土壤生态系统功能被抑制,土壤中微生物生物量降低。
(4)从维持高寒草甸生态系统功能方面考虑,30 g/m2施肥量可作为高寒草甸最佳施氮水平。高施肥量(≥40 g/m2)视为影响高寒草甸生态系统结构与功能、土壤养分及土壤微生物活性的阈值。施肥梯度下土壤有机碳和微生物量碳含量可作为衡量土壤肥力和土壤质量变化的重要指标。
[1] Li B Y,Wang J F,Zhao S J,Zhang H W,Zhao M X.The influence of fertilizers on the soil fertility,population structure and yield of Herbages Grown in Degraded Grassland.Grassland of China,2004,26(1):14-17.
[2] Zhou X D,Shen X L,Gao H W,Chen W D,Li T P,Lian D W.Effect of foliage dressing on forage yield and nutrition value of alpine grassland.Acta Prataculturae Sinica,2000,9(4):27-31.
[3] De K J,Zhou Q P,Xu C T.The influencing on herbage yield of grasslands by different manuring amount type of nitrogenous ferbilizer.Chinese Qinghai Journal of Animal and Veterinary Sciences,2001,31(3):12-13.
[4] Sun R,Liu C M,Zhu Q J.Relationship between the fractional vegetation cover change and rainfall in the Yellow River Basin.Acta Geographica Sinica,2001,56(6):667-672.
[5] Piao S L,Fang J Y.Dynamic vegetation cover change over the last 18 years in China.Quaternary Science,2001,21(4):294-302.
[6] Wang C T,Wang Q L,Jing Z C,Feng B F,Du Y G,Long R J,Cao G M.Vegetation roots and soil physical and chemical characteristic changes in Kobresia pygmaca meadow under different grazing gradients.Acta Prataculturae Sinica,2008,17(5):9-15.
[7] Li X Z,Chen Z Z.Influences of stocking rates on C,N,P contents in plant-soil system.Acta Agresila Sinica,1998,6(2):90-98.
[8] Wang C T,Long R J,Wang Q L,Cao G M,Shi J J,Du Y G.Response of plant diversity and productivity to soil resources changing under grazing disturbance on an alpine meadow.Acta Ecologica Sinica,2008,29(8):4144-4152.
[9] Wang Q L,Wang X,Cao G M,Wang C T,Long R J.Soil quality assessment of alpine meadow in Haibei State of Qinghai Province.Chinese Journal of Applied Ecology,2011,22(6):1416-1422.
[10] Wu R,Tiessen H.Effect of land use on soil degradation in alpine grassland soil,China.Soil Science Society of America Journal,2002,66(5):1648-1655.
[11] Zou C,Wang K,Wang T,Xu W.Overgrazing and soil carbon dynamics in eastern Inner Mongolia of China.Ecological Research,2007,22(1):135-142.
[12] Wang Q L,Cao G M,Wang C T.The impact of grazing on the activities of soil enzymes and soil environmental factors in alpine Kobresia pygmaea meadow.Plant Nutrition and Fertilizer Science,2007,13(5):856-864.
[13] Dong Q M,Zhao X Q,Ma Y S,Shi J J,Wang Y L,Sheng L.Effect of Grazing Intensity on soil organic matter and organic carbon in alpine-cold artificial grassland.Chinese Qinghai Journal of Animal and Veterinary Sciences,2007,37(1):6-8.
[14] Gough L,Osenberg C W,Gross K L,Collins S L.Fertilization effects on species density and primary productivity in herbaceous plant communities.Oikos,2000,89(3):428-439.
[15] Fridley J D.Resource availability dominates and alters the relationship between species diversity and ecosystem productivity in experimental plant communities.Oecologia,2002,132(2):271-277.
[16] Rajaniemi T K.Why does fertilization reduce plant species diversity?Testing three competition-based hypotheses.Journal of Ecology,2002,90(2):316-324.
[17] Wang C T,Long R J,Wang Q L,Liu W,Jing Z C,Zhang L.Fertilization and litter effects on the functional group biomass,species diversity of plants,microbial biomass and enzyme activity of two alpine meadow communities.Plant and Soil,2010,331(1/2):377-389.
[18] Printice I C,Farquhar G,Fasham M,et al.The carbon cycle and atmospheric carbon dioxide//Houghton J,ed.IPCC third Assessment Report.Cambridge:Cambridge University Press,2001:184-237.
[19] Bauer G A,Bazzaz F A,Minocha R,Long S,Magill A,Aber J,Berntson G M.Effects of chronic N additions on tissue chemistry,photosynthetic capacity,and carbon sequestration potential of a red pine(Pinus resirtosa Ait.)stand in the NE United States.Forest Ecology and Management,2004,196(1):173-186.
[20] Peng Q,Dong Y S,Qi Y C.Influence of external nitrogen input on key processes of carbon cycle in terrestrial ecosystem.Advances in Earth Science,2008,23(8):875-883.
[21] Zhu T H,Cheng S L,Fang H J,Yu G R,Deng J J,Li Y N.Early responses of soil CO2emission to simulating atmospheric nitrogen deposition in an alpine meadow on the Qinghai Tibetan Plateau.Acta Ecologica Sinica,2011,31(10):2687-2696.
[22] Aber J,McDowell W,Nadelhoffer K,Magill A,Berntson G,Kamakea M,McNulty S,Currie W,Rustad L,Fernandez I.Nitrogen saturation in temperate forest ecosystems-Hypotheses revisited.Bioscience,1998,48(11):921-934.
[23] Vitousek P M,Aber J,Howarth R,Likens G E,Matson P A,Scindler D W,Schlesinger W H,Tilman D G.Human alteration of the global nitrogen cycle:sources and consequences.Ecological Applications,1997,7(3):737-750.
[24] Li L J,Zeng D H,Yu Z Y,Ai G Y,Yang D,Mao R.Effects of nitrogen addition on grassland species diversity and productivity in Keerqin Sandy.Chinese Journal of Applied Ecology,2009,20(8):1838-1844.
[25] Li L J,Yu Z Y,Zeng D H,Ai G Y,Li J S.Effects of fertilizations on species composition and diversity of grassland in Keerqin Sandy Lands.Acta Prataculturae Sinica,2010,19(2):109-115.
[26] Zhang J Q,Li Q,Ren Z W,Ren Z W,Yang X,Wang G.Effects of nitrogen addition on species richness and relationship between species richness and aboveground productivity of alpine meadow of the Qinghai-Tibetan Plateau.Chinese Journal of Plant Ecology,2010,34(10):1125-1131.
[27] Xu Z Z,Zhou G S.Relationship between carbon and nitrogen and environmental regulation in plants under global change-From molecule to ecosystem.Journal of Plant Ecology,2007,31(4):738-747.
[28] Högberg P,Nordgren A,Buchmann N,Taylor A F S,Ekblad A,Högberg M N,Nyberg G,Ottosson-Löfvenius M,Read D J.Large-scale forest girdling shows that current photosynthesis drives soil respiration.Nature,2001,411(6839):789-792.
[29] Vitousek P M,Howarth R W.Nitrogen limitation on land and in the sea:how can it occur?Biogeochemistry,1991,13(2):87-115.
[30] LeBauer D S,Treseder K K.Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed.Ecology,2008,89(2):371-379.
[31] Qiu B,Luo Y J,Du G Z.The effect of fertilizer gradients on vegetation characteristics in alpine meadow.Acta Prataculturae Sinica,2004,13(6):65-68.
[32] Chen Y M,Li Z Z,Du G Z.Effects of fertilization on plant diversity and economic herbage groups in alpine meadow.Acta Botanica Boreali-Occidentalia Sinica,2004,24(3):424-429.
[33] Ji Y J.Effects of nitrogen on grass yield in Qinghai alpine meadow.Grassland of China,2005,27(6):67-69.
[34] Luo Y J.Plant Community Structure During Degradation and Restoration:a Theory Frame and Examples from an Alpine Meadow of Eastern Qinghai-Tibet Plateau[D].Lanzhou:Lanzhou University,2006.
[35] Niu K C,Luo Y J,Choler P,Du G Z.The role of biomass allocation strategy in diversity loss due to fertilization.Basic and Applied Ecology,2008,9(5):485-493.
[36] Zhang N L,Wan S Q,Li L H,Bi J,Zhao M M,Ma K P.Impacts of urea N addition on soil microbial community in a semiarid temperate steppe in northern China.Plant Soil,2008,311(1/2):19-28.
[37] Ren Z W,Li Q,Chu C J,Zhao L Q,Zhang J Q,de Ai X,Yang Y B,Wang G.Effects of resource additions on species richness and ANPP in an alpine meadow community.Journal of Plant Ecology,2010,3(1):25-31.
[38] Wang C T,Long R J,Ding L M.The effects of differences in functional group diversity and composition on plant community productivity in four types of alpine meadow communities.Biodiversity Science,2004,12(4):403-409.
[39] Ren J Z.Research methods of Pratacultural Science.Beijing:China Agricultural Press,1998:1-29.
[40] Institute of Nanjing Soil Science,Chinese Academy of Sciences.Soil analysis of Physics and Chemistry.Shanghai:Science and Technology Press,1983.
[41] Vance E D,Brooks P C,Jenkinson D S.An extraction method for measure soil microbial biomass C.Soil Biology and Biochemistry,1987,19(6):703-707.
[42] Zhang B Y,Chen T L,Wang B.Effects of long-term uses of chemical fertilizers on soil quality.Chinese Agricultural Science Bulletin,2010,26(11):182-187.
[43] Li Z J,Zhu Y C,Hu Y G.Effects of different variety and level of supplying fertilizer on the production of forage rye.Ⅰ.Effects on the hay yield of forage rye.Acta Prataculturae Sinica,2003,12(4):104-110.
[44] Rong Y P,Han J G,Wang P.Effects of different range restorations oil soil and vegetation of Russian wildrye(Psathyrostachys juncea)pasture.Acta Prataculturae Sinica,2002,11(1):17-23.
[45] Bao X K,Zhao B L.Study of effect soil nitrogen,phosphorus on primary productivity in the Kobresia humilis meadow//International Academic Symposium on Alpine Meadow System.Beijing:Science Press,1989:169-175.
[46] Zhang S L,Zhou X M.Effects of fertilization on aboveground biomass of the Kobresia humilis meadow.Acta Biologica Plateau Sinica,1986,(5):7-12.
[47] Warembourg F R,Esterlich H D.Plant phenology and soil fertility effects on below-ground carbon allocation for an annual(Bromus erectus)grass species.Soil Biology and Biochemistry,2001,33(10):1291-1303.
[48] van der Krifi T A J,Kuikman P J,Möiler F,Berendse F.Plant species and nutritional-mediated control over rhizodeposition and root decomposition.Plant Soil,2001,228(2):191-200.
[49] Wang X N,Wang Z H,Li S X.The effect of nitrogen fertilizer rate on summer maize yield and soil water-nitrogen dynamics.Acta Ecologica Sinica,2007,27(1):197-204.
[50] Aerts R,Caluwe DH,Beltman B.Is the relation between nutrient supply and biodiversity co-determined by the type of nutrient limitation?.Oikos,2003,101(3):489-498.
[51] Bai YF,Wu J G,Clark C M,Naeem S,Pan Q,Huang J H,Zhang L X,Xing G H.Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning:evidence from Inner Mongolia Grasslands.Global Change Biology,2010,16:358-372.
[52] Stevens M H H,Carson WP.Plant density determines species richness along an experiment fertility gradient.Ecology,1999,80(2):455-465.
[53] Zhang J E,Liao Z W.Discussion on soil ecological fertility and its cultivation.Soil and Environment Science,2000,9(3):253-256.
[54] Xu M G,Zhang J Q,Wen S L.The fertilizer effects and fertilization of herbages in red soil hilly regions of Southern China.Pratacultural Science,1997,14(6):21-23.
[55] Ma Y S,Lang B N,Li Q Y,Shi J J,Dong Q J.Effect of fertilizing nitrogen rate and time on Kobresia pygmaea meadow grassland.Pratacultural Science,2003,20(3):47-49.
参考文献:
[1] 李本银,汪金舫,赵世杰,张宏伟,赵明旭.施肥对退化草地土壤肥力、牧草群落结构及生物量的影响.中国草地,2004,26(1):14-17.
[2] 周学东,沈景林,高宏伟,陈卫东,李天平,连大伟.叶面施肥对高寒草地产草量及牧草营养品质的影响.草业学报,2000,9(4):27-31.
[3] 德科加,周青平,徐成体.不同施氮量对天然草场牧草产量影响.青海畜牧兽医杂志,2001,31(3):12-13.
[4] 孙睿,刘昌明,朱启疆.黄河流域植被覆盖度动态变化与降水的关系.地理学报,2001,56(6):667-672.
[5] 朴世龙,方精云.最近18年来中国植被覆盖的动态变化.第四纪研究,2001,21(4):294-302.
[6] 王长庭,王启兰,景增春,冯秉福,杜岩功,龙瑞军,曹广民.不同放牧梯度下高寒小嵩草草甸植被根系和土壤理化特征的变化.草业学报,2008,17(5):9-15.
[7] 李香真,陈佐忠.不同放牧率对草原植物与土壤C、N、P含量的影响.草地学报,1998,6(2):90-98.
[8] 王长庭,龙瑞军,王启兰,曹广民,施建军,杜岩功.放牧扰动下高寒草甸植物多样性、生产力对土壤养分条件变化的响应.生态学报,2008,29(8):4144-4152.
[9] 王启兰,王溪,曹广民,王长庭,龙瑞军.青海省海北州典型高寒草甸土壤质量评价.应用生态学报,2011,22(6):1416-1422.
[12] 王启兰,曹广民,王长庭.放牧对小嵩草草甸土壤酶活性及土壤环境因素的影响.植物营养与肥料学报,2007,13(5):856-864.
[13] 董全民,赵新全,马玉寿,施建军,王彦龙,盛丽.放牧强度对高寒人工草地土壤有机质和有机碳的影响.青海畜牧兽医杂志,2007,37(1):6-8.
[20] 彭琴,董云社,齐玉春.氮输入对陆地生态系统碳循环关键过程的影响.地球科学进展,2008,23(8):875-883.
[21] 朱天鸿,程淑兰,方华军,于贵瑞,郑娇娇,李英年.青藏高原高寒草甸土壤CO2排放对模拟氮沉降的早期响应.生态学报,2011,31(10):2687-2696.
[24] 李禄军,曾德慧,于占源,艾桂艳,杨丹,毛瑢.氮素添加对科尔沁沙质草地物种多样性和生产力的影响.应用生态学报,2009,20(8):1838-1844.
[25] 李禄军,于占源,曾德慧,艾桂艳,李晶石.施肥对科尔沁沙质草地群落物种组成和多样性的影响.草业学报,2010,19(2):109-115.
[26] 张杰琦,李奇,任正炜,任正炜,杨雪,王刚.氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响.植物生态学报,2010,34(10):1125-1131.
[27] 许振柱,周广胜.全球变化下植物的碳氮关系及其环境调节研究进展——从分子到生态系统.植物生态学报,2007,31(4):738-747.
[31] 邱波,罗燕江,杜国祯.不同施肥梯度对高寒草甸植被特征的影响.草业学报,2004,13(6):65-68.
[32] 陈亚明,李自珍,杜国祯.施肥对高寒草甸植物多样性和经济类群的影响.西北植物学报,2004,24(3):424-429.
[33] 纪亚珺.青海高寒天然草地施氮肥试验.中国草地,2005,27(6):67-69.
[34] 罗燕江.退化和恢复中的植物群落:以高寒草甸为例的理论和试验研究[D].兰州:兰州大学,2006.
[38] 王长庭,龙瑞军,丁路明.高寒草甸不同草地类型功能群多样性及组成对植物群落生产力的影响.生物多样性,2004,12(4):403-409.
[39] 任继周.草业科学研究方法.北京:中国农业出版社,1998:1-29.
[40] 中国科学院南京土壤研究所.土壤理化分析.上海:上海科学技术出版社,1983.
[42] 张北赢,陈天林,王兵.长期施用化肥对土壤质量的影响.中国农学通报,2010,26(11):182-187.
[43] 李志坚,祝延成,胡跃高.不同施肥水平与组合对饲用黑麦生产性能的影响研究.Ⅰ.对饲用黑麦产草量的影响.草业学报,2003,12(4):104-110.
[44] 戎郁萍,韩建国,王培.不同草地恢复方式对新麦草草地土壤和植被的影响.草业学报,2002,11(1):17-23.
[45] 鲍新奎,赵保莲.土壤氮、磷养分对矮嵩草草甸初级生产力影响的初步研究//中国科学院西北高原生物研究所.高寒草甸生态系统国际学术讨论会论文集.北京:科学出版社,1989:169-175.
[46] 张松林,周兴民.施肥对矮嵩草草甸草场地上生物量的影响.高原生物学集刊,1986,(5):7-12.
[49] 王西娜,王朝辉,李生秀.施氮量对夏季玉米产量及土壤水氮动态的影响.生态学报,2007,27(1):197-204.
[53] 章家恩,廖宗文.试论土壤的生态肥力及其培育.土壤与环境,2000,9(3):253-256.
[54] 徐明岗,张久权,文石林.南方红壤丘陵区牧草的肥料效应与施肥.草业科学,1997,14(6):21-23.
[55] 马玉寿,郎百宁,李青云,施建军,董全民.施氮量与施氮时间对小嵩草草甸草地的影响.草业科学,2003,20(3):47-49.
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
现代农村科技(2022年9期)2022-08-16
青海草业(2022年2期)2022-07-23
活力(2019年21期)2019-04-01
河北林业科技(2016年5期)2016-11-08
浙江农业学报(2016年7期)2016-06-15
中国农业文摘-农业工程(2016年5期)2016-04-12
现代农业(2016年4期)2016-02-28
环境科技(2015年2期)2015-11-08
湖南农业科学(2014年20期)2014-02-27
