
水涝胁迫下海州常山形态和生理指标的变化
2013-12-05 02:49:16曾德静尹丽娟
浙江农林大学学报 2013年2期
曾德静,王 铖,刘 军,尹丽娟
(1.上海市园林科学研究所,上海 200232;2.四川省眉山市园林局,四川 眉山 620020;3.四川农业大学 林学院,四川 雅安 625014)
随着全球环境的不断恶化,生态系统遭到严重破坏,全球气候异常加剧,雨量分布极不均衡,局部地区水灾不断,土壤淹水现象极为常见。上海地区夏季的梅雨季节,降水量大,持续时间长,容易造成局部土壤积水,对该地区植物的耐涝能力提出了一定的要求。目前,关于植物耐涝方面的研究主要集中在作物、造林树种及果树方面,对园林观赏植物的研究还相对较少,且对植物耐涝能力研究主要是在淹水条件下植物的形态和生理变化方面进行的,对于不同水涝的程度即水位的高度,对植物的影响方面的报道极少。海州常山Clerodendrum trichotomum是马鞭草科Verbenaceae大青属Clerodendrum,落叶灌木或小乔木[1]。海州常山花开时红白相间,花朵繁密似锦,秋季果实成熟时,亮蓝紫色的球形果与花同时宿存枝顶,艳丽可爱,是点缀园林、庭院的优良观花观果植物。它抗性强,是一种很有潜力的盐碱地区园林绿化和城市废弃地恢复植物[2-3]。同时,该植物具有一定的药用价值,有降压、镇静、镇痛等作用[4],茎皮中抗肿瘤活性成分高达4.6%[5]。另外,据罗艳等[6]报道,海州常山的种子含油量达34.1%,是值得推广栽植的生物柴油植物。目前,关于海州常山抗性研究主要集中在光强[7]、盐胁迫[8-9]及二氧化硫[10]等方面,对于其耐水涝能力方面的研究报道很少,只有魏娟等[11]对海州常山在湿害、涝害和干旱等3种条件下的生理特性进行了研究,但并未对其耐受程度进行研究。本试验对海州常山耐水涝程度进行进一步研究,通过设置不同的淹水高度,观察各处理下植株的形态变化,并对其相关生理指标进行测定和分析,初步得出其耐水涝胁迫的能力,以期为海州常山的绿化应用、耐涝育种以及作为药物、生物柴油等栽植提供相关的依据。
1 材料与方法
1.1 材料
选取生长健壮、一致的海州常山1年生扦插盆栽苗作为试验材料,苗高为80~100 cm。于试验前1个月将盆栽苗移入上海市园林科学研究植物资源与环境试验区防雨棚内进行缓苗,并进行正常的日常管理。
1.2 试验方法
试验采用大盆套小盆的方法,设置不同的水位高度对海州常山进行水涝胁迫试验。盆栽苗盆的直径为20 cm,高25 cm,盆土厚度为22 cm。试验分水湿和淹水2种类型的处理,水湿处理设置水位达到1/4盆土厚度、1/2盆土厚度、2/3盆土厚度、盆土表面共4个水平,分别设为处理A,处理B,处理C和处理D。淹水处理为水位达盆土表面以上5 cm处,设为处理E。为防止水分蒸发,引起大盆内的水位下降,试验开始后,定期向处理植株的大盆内补水,保证水位维持在所设定的位置,对照为正常管理。自处理当天起,2 d观察1次,记录植株外观形态特征变化,并取植株上部第3~5叶序叶片带回实验室进行各项生理指标的测定,直至叶片枯黄或植株死亡。
1.3 测定指标及方法
叶片相对含水量、叶绿素质量分数、相对电导率、脯氨酸质量分数、可溶性糖质量分数、可溶性蛋白质量分数测定参照王学奎[12]的方法,超氧化物歧化酶(SOD)活性参照朱广廉[13]的方法测定,丙二醛(MDA)质量摩尔浓度测定参照刘祖祺等[14]的方法。
1.4 数据统计与分析
所有试验数据采用Microsoft Office Excel 2003和SPSS 18.0进行统计、分析。
2 结果与分析
2.1 水涝胁迫对海州常山形态的影响
处理A和处理B下,海州常山植株长势良好,与对照相比,形态无明显变化。处理C下,第6天,海州常山植株部分下部叶片起皱,开始出现萎蔫症状,中上部叶片形态无明显变化;第8天,植株下部叶片萎蔫现象明显,边缘卷曲,叶片泛黄,开始下垂;第10天,中部叶片均失水萎蔫,卷曲,下部部分叶片下垂,已经枯黄而死亡;第12天,植株失水严重,萎蔫范围进一步扩大,上部叶片出现轻度萎蔫,中部全部失水下垂,下部叶片完全干枯死亡,开始脱落。处理D和处理E之间植物形态变化基本一致,无明显的差异,在处理第2天,植株部分叶片开始出现萎蔫,仅表现在最下部的叶片上;第4天,植株下部叶片失水萎蔫,叶片起皱,边缘卷曲,明显下垂,中部叶片开始出现萎蔫现象,上部叶片无明显的变化;第6天,中部叶片出现大面积失水萎蔫下垂,上部叶片也失水变皱,下部叶片已经干枯死亡,开始脱落;第8天,上部叶片已卷曲下垂,中下部已干枯死亡,整个植株全部萎蔫,接近死亡。
2.2 水涝胁迫对海州常山生理指标的影响
2.2.1 对相对含水量的影响 海州常山叶片含水量随着湿涝胁迫时间延长而减小,且胁迫程度越高,叶片含水量越低(图1)。处理A和处理B的含水量在整个胁迫期间与对照的差异不显著(P>0.05),处理C在胁迫前6 d相对含水量变化不大,第8天开始出现明显失水现象,且之后每2 d的含水量的差异达到显著水平(P<0.05),处理D和处理E从第4天起含水量就显著(P<0.05)下降,且下降幅度较大,第8天分别比对照减少43.19%和46.24%。
2.2.2 对叶绿素质量分数的影响 由图2可知:水涝胁迫下,不同处理之间海州常山叶片叶绿素质量分数变化各不相同。处理A在整个胁迫期间叶绿素质量分数能保持稳定,叶绿素质量分数与对照差异不显著(P>0.05);随着胁迫时间延长,处理B,处理C,处理D和处理E叶绿素质量分数先下降后升高,处理B叶绿素质量分数缓慢下降,第6天与对照差异达到显著水平(P<0.05);处理C叶绿素质量分数在第8天急速下降,之后又急速升高,在第12天显著(P<0.05)高于对照的质量分数;处理D和处理E叶绿素质量分数在胁迫第2天明显低于对照,随胁迫时间延长,叶绿素质量分数持续升高,且上升幅度较大,胁迫结束时显著高于对照(P<0.05)。
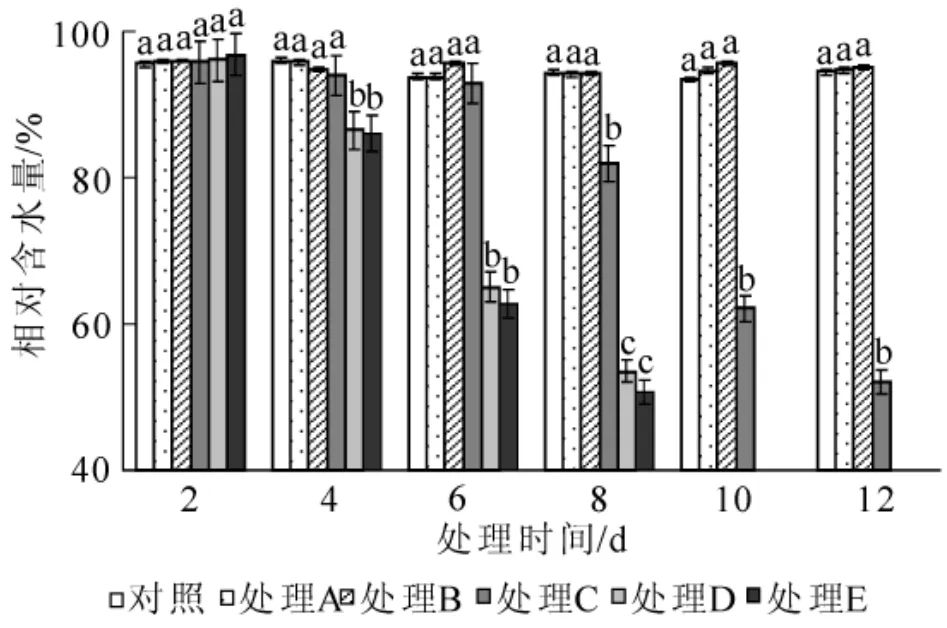
图1 水涝胁迫对相对含水量的影响Figure 1 Effect of water logging on relative water content
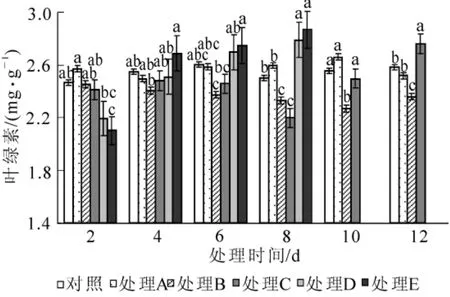
图2 水涝胁迫对叶绿素质量分数的影响Figure 2 Effect of water logging on chlorophy content
2.2.3 对相对电导率和丙二醛质量摩尔浓度的影响 水涝胁迫下,海州常山叶片相对电导率随胁迫时间延长逐渐增大(图3)。在胁迫前期,各处理的相对电导率水平较低,随着胁迫时间的延长,处理A和处理B变化不大,与对照无明显差异(P>0.05);处理C,处理D和处理E的相对电导率持续升高,处理C在第6天时与对照差异显著(P<0.05),处理D和处理E在第4天与对照差异就达到显著水平(P<0.05),升高幅度大小依次是处理E>处理D>处理C。由图4可以看出:水涝胁迫引起海州常山叶片的丙二醛质量摩尔浓度增加。处理A和处理B在整个胁迫过程中丙二醛质量摩尔浓度稳定,虽有起伏但差异不显著,与对照也无显著差异(P>0.05);处理 C,处理D和处理 E随着胁迫时间延长,丙二醛质量摩尔浓度显著(P<0.05)升高,试验结束时,丙二醛质量摩尔浓度分别达到对照的2.71倍、2.68倍和2.74倍。
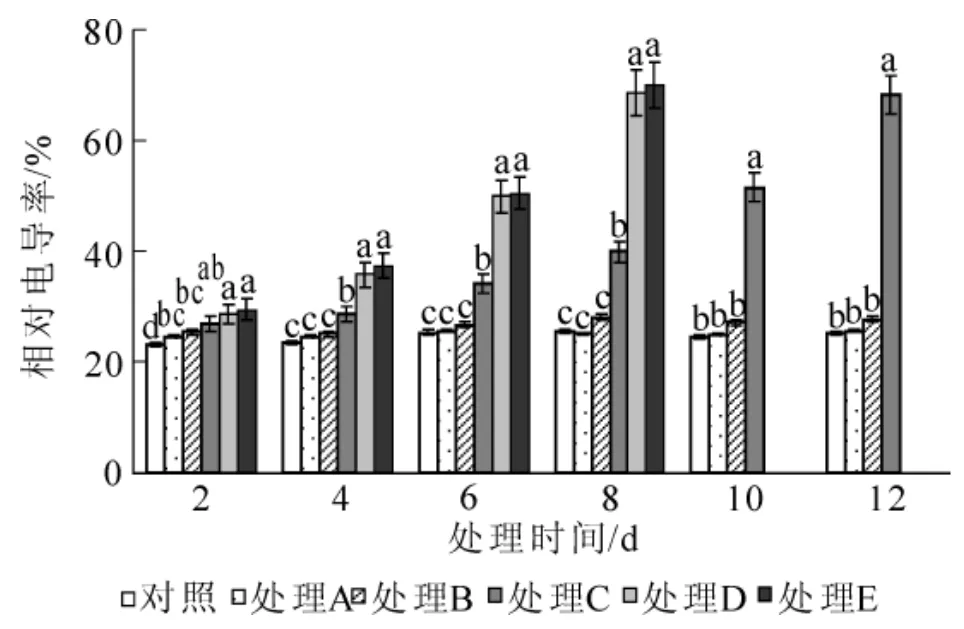
图3 水涝胁迫对相对电导率的影响Figure 3 Effect of water logging on relative electrical conductivity
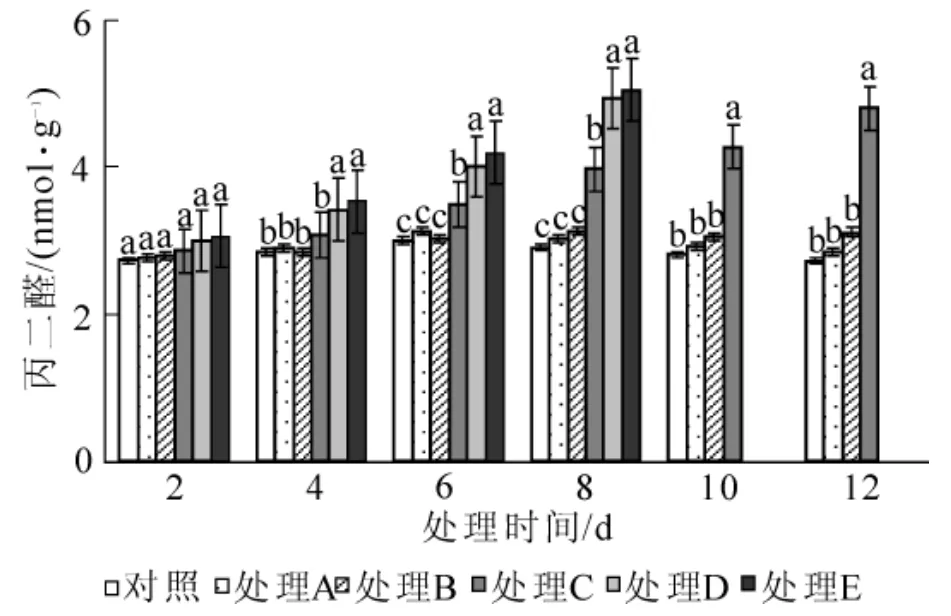
图4 水涝胁迫对丙二醛质量摩尔浓度的影响Figure 4 Effect of water logging on MDA content
2.2.4 对脯氨酸、可溶性糖、可溶性蛋白质量分数的影响 如图5所示:当水涝胁迫严重时,海州常山叶片脯氨酸质量分数迅速增加。各处理的脯氨酸质量分数变化规律与丙二醛变化一致,变化幅度更大。处理A和处理B在整个胁迫过程中与对照无显著差异(P>0.05);处理C,处理D和处理E脯氨酸质量分数随胁迫时间延长而急剧增加,最大值分别达到了对照的21.9倍、24.2倍和24.8倍。水涝胁迫能促使海州常山体内可溶性糖质量分数的增加(图6)。处理A在整个胁迫期间的可溶性糖质量分数变化不明显,与对照无显著差异(P>0.05);处理B在胁迫前8 d可溶性糖质量分数缓慢升高,第8天与对照差异显著(P<0.05),之后维持在较高水平;处理C,处理D和处理E可溶性糖质量分数在胁迫中持续增加,相比之下处理C增加速度较慢;处理D和处理E可溶性糖质量分数迅速增加,每次测定值间均存在显著差异(P<0.05),但两者之间差异不显著(P>0.05)。水胁迫下海州常山叶片可溶性蛋白质量分数的变化见图7。在水涝胁迫过程中,处理A和处理B的可溶性蛋白质量分数虽有所波动,但与对照组的差异不显著(P>0.05);处理C可溶性蛋白质量分数逐渐下降,第4天时与对照差异显著(P<0.05),第10天降到最低,之后在第12天有所回升;处理D和处理E可溶性蛋白质量分数出现先降后升趋势,谷值出现在第6天,在整个胁迫过程中可溶性蛋白质量分数均显著(P<0.05)低于对照。
2.2.5 对超氧化物歧化酶活性的影响 对照、处理A和处理B在各时间阶段下超氧化物歧化酶活性呈现一定的波动性,可能与外界环境因素如温度等的变化有关,但在各胁迫时间段,处理A和处理B的超氧化物歧化酶活性与对照差异不显著(P>0.05);处理C,处理D和处理E随着胁迫时间的延长,超氧化物歧化酶活性呈持续下降趋势,降幅较大直至植株死亡(图8)。
3 讨论
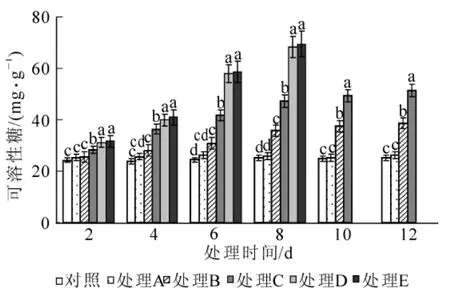
图6 水涝胁迫对可溶性糖质量分数的影响Figure 6 Effect of water logging on soluble sugar content
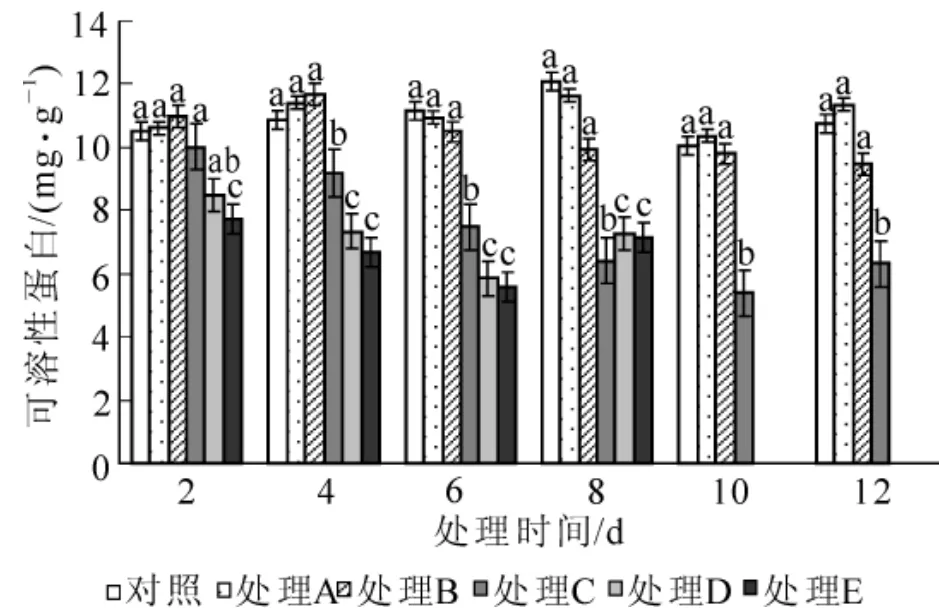
图7 水涝胁迫对可溶性蛋白质质量分数的影响Figure 7 Effect of water logging on soluble protein content
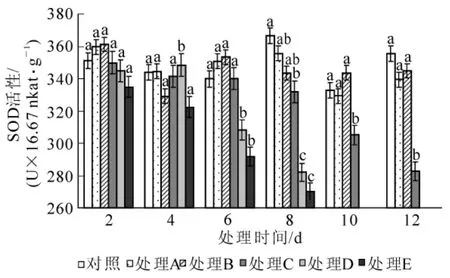
图8 水涝胁迫对超氧化物歧化酶活性的影响Figure 8 Effect of water logging on SOD activities
水涝胁迫使植物处于周期或长期的厌氧或缺氧状态,限制植物的需氧呼吸和维持生命活动所需的能量产生,从而对植物的水分代谢、光合作用、细胞膜系统、抗氧化系统、能量代谢等造成伤害。为了适应或减轻缺氧伤害,植物会启动形态、生理方面的适应机制。形态方面主要表现为叶片形态的改变、茎伸长生长、不定根和通气组织的形成等[15-16]。生理方面表现为代谢途径改变,通过厌氧代谢获得维持生命的能量;通过体内乙烯、赤霉素和脱落酸等激素水平的改变来调节生理活动或形态等方面的变化;抗氧化酶系统对活性氧自由基的清除等[17]。糖酵解被认为是植物对厌氧胁迫适应的基本功能,维持糖酵解途径需要源源不断的可溶性糖的供应[17],因此,水涝胁迫下植株可溶糖的增加,对植物生长的能量供应至关重要。脯氨酸除作为渗透调节物质外,还可以是能量库、氮素储存剂、羟基清除剂、保护酶系和细胞结构稳定的兼容性溶剂。水涝胁迫下,植物往往通过脯氨酸、可溶性糖的大量积累来适应环境,这在紫穗槐 Amorpha fruticosa[18],金钱松 Pseudolarix kaempferi[19]等中得到了验证。抗氧化酶能够清除植物体内的活性氧自由基,防止细胞膜脂过氧化,对植物形成有效保护。大量研究[20-21]显示,水涝胁迫下,植物的抗氧化酶系统能产生及时的响应,来增强植物的耐涝能力,从而减轻伤害。本试验结果显示,海州常山在水涝胁迫过程中其体内发生了一系列的生理变化。随着胁迫程度的增加,相对电导率、丙二醛质量分数的持续上升,说明在胁迫过程中海州常山细胞膜系统受到的伤害逐渐加重;超氧化物歧化酶活性持续下降,说明水涝胁迫下,海州常山的抗氧化酶系统不敏感,不能够做出迅速而适当的调整,而逐渐受到伤害;可溶性糖质量分数、脯氨酸质量分数不断升高,最高分别为对照的2.77倍和24.82倍,说明海州常山具有极强的渗透调节能力,能够通过脯氨酸和可溶性糖的大量积累来适应环境。
从海州常山植株在干旱胁迫[11]、水涝胁迫和土壤盐胁迫[8]下的生理指标变化来看,不同类型胁迫下海州常山的生理响应既有相同点也有不同点。3种胁迫下,植株的叶绿素质量分数均下降,光合作用受到影响;膜的透性和膜脂过氧化产物均增加,表现为相对电导率和丙二醛质量摩尔浓度的增加,导致细胞膜伤害加重;植株均具有较强的渗透调节能力,能够通过可溶性糖和脯氨酸的主动积累来适应胁迫环境。但抗氧化酶在胁迫中的表现不同,盐胁迫下植株能够迅速提高过氧化物歧化酶和过氧化物酶活性来增强植物的抗逆能力,干旱胁迫和水涝胁迫下过氧化物歧化酶的活性在试验初期能维持在对照组的水平,之后随胁迫时间延长过氧化物歧化酶活性逐渐下降,说明在水分胁迫下,海州常山植株的抗氧化酶系统不敏感,难以做出迅速而适当的调整,植株逐渐受到伤害。
本试验中,1/4和1/2盆土厚度的水位高度处理对海州常山影响不大,植株生长正常;而水位达到2/3盆土厚度处理,在胁迫12 d之后植株死亡。综合各生理指标可知,处理C在胁迫前期对水涝有一定的抗性,随着胁迫时间的延长,抗性降低,植株逐渐死亡,说明处理C对海州常山有伤害,但伤害较缓慢;当水位达到盆土表面或超过表面,对海州常山伤害较大,各生理指标变化十分剧烈,8 d后植株死亡。可见,海州常山耐水湿能力较强,1/2盆土厚度是其极限水位高度,耐水淹能力一般且持续时间不长。
[1]裴鉴,陈守良.中国植物志:第65卷[M].北京:科学出版社,1982:186-187.
[2]包志毅,陈波.工业废弃地生态恢复中的植被重建技术[J].水土保持报,2004,18(3):160-163.BAO Zhiyi,CHEN Bo.Study on revegetation during ecological restoration of industrial wasteland[J].J Soil Water Conserv,2004,18(3):160-163.
[3]MAHMUC R,INONE N,KASAJIMAS,et al.Assessment of potential indigenous plant species for the phytoremediation of arsenic-contaminated areas of Bangladesh[J].Int J Phytorem,2008,10(2):119-132.
[4]徐叔云.臭梧桐的药理学研究[J].生理学报,1962,25(4):272-276.XU Shuyun.The pharmacological research of Clerodendrum trichotomum[J].J Physiol,1965,25(4):272-276.
[5]NAGAO T,ABE F.Antiproliferative constituents in the plant 7.leaves of Clerodendron bungei and leaves and bark of C.trichotomum[J].Biol Pharm Bull,2001,24(11):1338-1341.
[6]罗艳,刘梅.开发木本油料植物作为生物柴油原料的研究[J].中国生物工程杂志,2007,27(7):68-74.LUO Yan,LIU Mei.Oil-bearing trees as a source for biodiesel[J].China Biotechnol,2007,27(7):68-74.
[7]胡凤琴,杨文杰,徐贵明,等.不同光强与水分条件对海州常山幼苗形态与生长的影响[J].南京林业大学学报:自然科学版,2009,33(5):27-30.HU Fengqin,YANG Wenjie,XU Guiming,et al.Influence of light intensity and soil water content on morphological and growth traits of Clerodendrum trichotomum seedling[J].J Nanjing For Univ Nat Sci Ed,2009,33(5):27-30.
[8]谢福春,张文婷,刘富强,等.土壤盐胁迫对海州常山生理生化特性的影响[J].江西农业大学学报,2008,30(5):839-844.XIE Fuchun,ZHANG Wenting,LIU Fuqiang,et al.Effect of soil NaCl stress on physiological and biochemical characteristics of Clerodendrum trichotonum[J].Acta Agric Univ Jiangxi,2008,30(5):839-844.
[9]谢福春,陈才业,张文婷,等.土壤盐胁迫对海州常山形态与气体交换特性的影响[J].浙江林学院学报,2009,26(2):176-181.XIE Fuchun,CHEN Caiye,ZHANG Wenting,et al.Soil NaCl stress and photosynthetic characteristics with Clerodendrum trichotomum[J].J Zhejiang For Coll,2009,26(2):176-181.
[10]郑立文,胡丁猛,刘伟峰,等.11个园林绿化树种对SO2胁迫的适应性反应[J].山东农业大学学报:自然科学版,2006,37(3):363-368.ZHENG Liwen,HU Dingmeng,LIU Weifeng,et al.Study of 11 ornamental tree species on sulfur dioxide stress resistance[J].J Shandong Agric Univ Nat Sci Ed,2006,37(3):363-368.
[11]魏娟,谢福春,王华田,等.水分胁迫对海州常山抗逆生理特性的影响[J].山东农业大学学报:自然科学版,2009,40(3):371-376.WEI Juan,XIE Fuchun,WANG Huatian,et al.Effect of water stress on growth and physiological characteristics of Clerodendrum trichotomum[J].J Shandong Agric Univ Nat Sci Ed,2009,40(3):371-376.
[12]王学奎.植物生理生化实验原理与技术[M].北京:高等教育出版社,2006.
[13]朱广廉.植物生理学实验[M].北京:北京大学出版社,1990.
[14]刘祖祺,张石城.植物抗性生理学[M].北京:中国农业出版社,1994.
[15]赵可夫.植物对水涝胁迫的适应[J].生物学通报,2003,38(12):11-14.ZHAO Kefu.Adaptation of plants to waterlogging stress[J].Bull Biol,2003,38(12):11-14.
[16]陈芳清,李永,郄光武,等.水蓼对水淹胁迫的耐受能力和形态学响应[J].武汉植物学研究,2008,26(2):142-146.CHEN Fangqing,LI Yong,QIE Guangwu,et al.The morphological responses and endurance of Polygonum hydropiper to flooding stress[J].J Wuhan Bot Res,2008,26(2):142-146.
[17]谭淑端,朱明勇,张克荣,等.植物对水淹胁迫的响应与适应[J].生态学杂志,2009,28(9):1871-1877.TAN Shuduan,ZHU Mingyong,ZHANG Kerong,et al.Response and adaptation of plants to submergence stress[J].Chin J Ecol 2009,28(9):1871-1877.
[18]王瑞,梁坤伦,周志宇,等.不同淹水梯度对紫穗槐营养生长和生理响应[J].草业学报,2012,21(1):149-155.WANG Rui,LIANG Kunlun,ZHOU Zhiyu,et al.Effect of different waterlogging stress condition on growth and some physiological characteristics of Amorpha fruticosa[J].Acta Pratacul Sin,2012,21(1):149-155.
[19]廖文燕,高捍东.金钱松幼苗在淹水胁迫过程中的生理响应[J].林业科技开发,2011,25(3):27-31.LIAO Wenyan,GAO Handong.Physiological responses of Pseudolarix kaempferi seedlings to waterlogging stress[J].China For Sci Technol,2011,25(3):27-31.
[20]李纪元.涝渍胁迫对枫杨幼苗保护酶活性及膜脂过氧化物的影响[J].安徽农业大学学报,2006,33(4):450-453.LI Jiyuan.Variations of activities for protecting enzymes and superoxide of lipid membrane in Chinese wingnut tree seedlings(Pterocarya stenoptera)after flooding treatment[J].J Anhui Agric Univ,2006,33(4):450-453.
[21]曾淑华,赵正雄,覃鹏,等.淹水对转超氧化物歧化酶或过氧化氢酶基因烟草某些生理生化指标的影响[J].植物生理学通讯,2005,41(5):603-606.ZENG Shuhua,ZHAO Zhengxiong,QIN Peng,et al.Effects of waterlogging on some physiological and biochemical indexes of transgenic tobacco(Nicotiana tobacum L.)lines with superoxide dismutaseor peroxidase gene[J].Plant Physiol Commun,2005,41(5):603-606.
猜你喜欢
江苏地方志(2023年6期)2024-01-18 07:07:48
江苏农村经济(2023年9期)2023-09-28 08:40:38
小学生优秀作文(低年级)(2022年3期)2022-03-29 07:09:14
中国现代中药(2022年12期)2022-02-18 08:24:16
连云港师范高等专科学校学报(2021年4期)2021-03-04 05:45:24
连云港师范高等专科学校学报(2021年4期)2021-03-04 05:45:22
世界科学(2020年1期)2020-02-11 07:39:34
中国生物医学工程学报(2019年5期)2019-07-16 07:56:56
传媒评论(2019年4期)2019-07-13 05:49:24
Coco薇(2017年5期)2017-06-05 13:03:24
