
产超广谱β-内酰胺酶菌株质粒图谱与耐药性分析
2013-11-21 03:06陆瑶张春云张守响姜文心于红
精准医学杂志 2013年6期
陆瑶,张春云,张守响,姜文心,于红
(青岛大学医学院,山东 青岛 266071)
产超广谱β-内酰胺酶(ESBLs)菌株的耐药性已经成为当前全球最重要的医院耐药问题之一[1-2]。ESBLs是由细菌质粒介导的能水解青霉素类、头孢菌素类及单酰环类抗生素的一种β-内酰胺酶,但不水解头霉素类和碳青霉烯类抗生素,而且可以被β-内酰胺酶抑制剂(如克拉维酸、舒巴坦)所抑制[3]。临床上,大肠埃希菌和肺炎克雷伯菌是最常见的产ESBLs细菌。研究结果表明,ESBLs的编码基因多定位于细菌所携带的质粒上,相对于染色体,质粒所携带的耐药基因更易于播散,因此,耐药质粒在产ESBLs菌株的多重耐药机制中发挥着重要作用。已有研究结果显示,质粒图谱在不同的地区与时间会存在一定的差异性,因此,研究细菌质粒的携带情况及其与耐药性的关系,对于控制疾病的发生、流行和治疗均具有指导意义[4]。本实验拟通过分析青岛地区临床分离的产ESBLs菌株的质粒图谱与耐药表型的相关性,为进一步深入研究ESBLs菌株质粒与耐药性的关系打下基础,同时亦为本地区临床抗生素的使用提供有意义的参考。
1 材料和方法
1.1 菌株
本实验收集的51株产ESBLs菌,来源于2010年6月—2011年5月青岛市市立医院检验科分离鉴定的临床标本(痰液、尿液、血液及脑脊液),已去除重复标本。所有菌株经美国临床实验室标准化委员会(NCCLS)1999年推荐纸片扩散法进行筛选实验,鉴定为产ESBLs菌,主要为大肠埃希菌及肺炎克雷伯菌。
1.2 微生物敏感性试验
采用西门子MicroScan Wal k Away 96 Pl us微生物鉴定-药敏测试系统,检测51株产ESBLs菌对24种常用抗生素(亚胺硫霉素、替卡西林、氨曲南、头孢哌酮-舒巴坦、复方磺胺甲 唑、氨苄西林-舒巴坦、美罗培南、妥布霉素、丁胺卡那霉素、庆大霉素、一代头孢唑林、二代头孢西丁、三代头孢噻肟、头孢他啶、头孢曲松、头孢泊肟、四代头孢吡肟、阿莫西林、氨苄青霉素、哌拉西林-三唑巴坦、氧哌嗪青霉素、环丙氟哌酸、加替沙星、罗米沙星)的敏感性,参照美国临床实验室标准化研究标准(2005年)进行操作及结果判断。
1.3 质粒DNA的抽提
采用碱裂解法提取质粒DNA[5]。
1.4 琼脂糖凝胶电泳检测质粒DNA条带
配制含有0.5 mg/L溴化乙锭的8 g/L的琼脂糖凝胶,取5μL质粒DNA,加上样缓冲液1μL,在80 V/cm的电压下电泳1.5 h,以λDNA/HindⅢ作为相对分子质量标记,将电泳图谱于紫外线灯下观察、记录结果。
1.5 统计学分析
采用SPSS 18.0软件进行统计学处理,采用χ2检验对细菌耐药性与质粒条带数进行相关性分析。
2 结 果
2.1 产ESBLs菌株对抗菌药物的耐药性
本文51株菌株对一代、二代、三代头孢药物耐药性最高。对氨曲南、头孢曲松、头孢他啶、氧哌嗪青霉素的耐药率也较高。而对美罗培南、亚胺硫霉素、丁胺卡那霉素、头孢哌酮-舒巴坦的耐药率较低;对亚胺硫霉素、美罗培南、丁胺卡那霉素、哌拉西林-三唑巴坦敏感。见表1。

表1 51株产ESBLs菌株对抗菌药物的耐药情况(χ/%)
2.2 产ESBLs菌株质粒电泳图谱
电泳结果显示,51株产ESBLs菌株质粒的检出率为62.75%,不同菌株中质粒数目1~3条(图1),每种质粒图谱中质粒条带分布见表2。其中,各菌株中均含有23 130 bp条带。

图1 产ESBLs菌株质粒电泳图谱
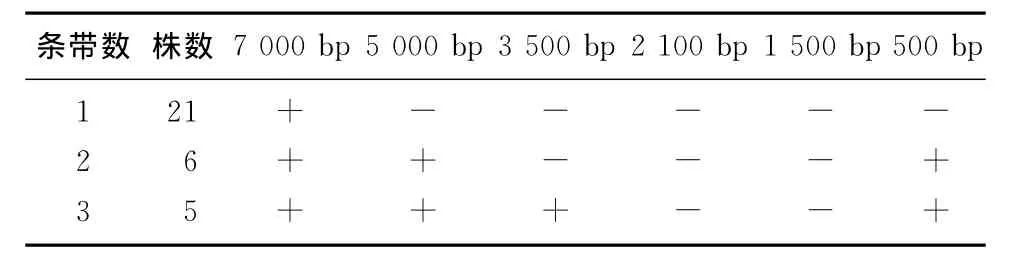
表2 产ESBLs菌株质粒条带分布
2.3 质粒条带数与细菌耐药率的关系
菌株对氨曲南、氨苄青霉素、头孢唑林、头孢吡肟、头孢噻肟、头孢他啶、头孢曲松的耐药率高于90%;对美罗培南、亚胺硫霉素、丁胺卡那霉素、头孢哌酮-舒巴坦的耐药率则低于20%。见表3。菌株质粒条带数与耐药性无显著相关性(P>0.05)。

表3 细菌质粒条带数与耐药率的关系(χ/%)
3 讨 论
ESBLs菌株是临床中最常见的耐药菌株之一,由于抗生素的滥用导致抗生素的选择压力逐渐增大,部分 ESBLs菌株甚至出现了多重耐药性[6-7]。目前对细菌耐药性的研究表明,质粒在异种菌株以及同种菌株耐药性传播中起关键作用,特别是质粒携带的产ESBLs耐药基因。质粒是最早发现的染色体外DNA,常携带有多种外源基因,其编码的性状与细菌的选择性优势和耐药性密切相关。研究表明,ESBLs耐药基因多定位于质粒上,不同的复制起始点的质粒具有相容性,导致细菌体内可有多种质粒共存。基于质粒可通过转导、接合等方式在同种或异种细菌间传播,其编码的耐药性状亦可随之扩散,这是产ESBLs菌株得以在不同地域间迅猛发展的分子机制[8]。由于各国(地区)使用抗生素的习惯不同,其ESBLs分布呈明显的地域性分布特点。
ESBLs属于A mbler分类的a类,按Bush分类属2be类,是一类能够水解青霉素、头孢菌素类及单环β-内酰胺类抗菌药物的β-内酰胺酶,普遍认为是一种质粒介导酶[9-10],同时可以对磺胺类、氨基糖苷类交叉耐药。研究表明,ESBLs的编码基因多位于细菌所携带的质粒上,而临床野生耐药菌株往往携带多种质粒。目前,对于ESBLs编码基因是定位于一种质粒,还是存在于同一细菌体内的不同质粒之上的报道较少[11]。
质粒图谱本质上是细菌内不同相对分子质量质粒的琼脂糖电泳迁移谱带,其测定简单、快速。通过分析细菌的质粒图谱,可以确定不同细菌耐药性状的来源以及菌株的同源性。同时,为临床分离菌株的分子流行病学研究,指导临床用药及耐药菌株的监测提供线索。
质粒图谱在不同的地区与时间会存在一定的改变。张志坚等[12]对郑州临床分离的46株产ESBLs大肠杆菌进行质粒图谱分析,结果表明,质粒的携带率为100%,菌株含有1~5条质粒条带。本研究中菌株质粒携带率为62.75%,检出率偏低,而且质粒谱的构成也有一定差别,可能与质粒的提取方法及临床菌株的来源不同有关。
本实验结果显示,含不同质粒条带的ESBLs菌株对头孢菌素类和青霉素类抗生素均具有普遍的耐药性,而对碳青霉烯类、氨基糖苷类抗生素敏感,与国内文献报道的结果相似[13],提示临床上一旦确诊为产ESBLs菌株感染,应立即停用头孢菌素类和β-内酰胺类抗生素,而首先选用碳青霉烯类或氨基糖苷类抗生素治疗。统计学分析结果表明,各菌株耐药性与质粒条带数无显著相关性,这与文献报道的结果一致[14]。同时研究结果显示,产ESBLs菌株可携带1~3条质粒,各菌株都在约7 000 bp处有一条质粒条带。基于质粒种类、数量与细菌耐药性无相关性,提示产酶基因可能定位于约7 000 bp的质粒条带上。此外,含相同质粒条带数的菌株其质粒图谱不同,原因可能为细菌种类不同或同种细菌但非同一感染菌株发生院内感染并在院内各病区播散[15]。
本实验仅对一定时间内收集的部分产ESBLs耐药菌株进行检测,在质粒水平上研究了其质粒与耐药谱的关系,其结果仅能解释部分菌株的耐药性及其同源性的问题和医院内流行菌株的问题,以指导本地区医院内的抗生素使用,而对于耐药基因的具体质粒定位,还需要进一步研究探讨。同时,对于本地区耐药菌株的流行趋势、耐药机制以及与其他地区的比较,均需要进行大量长期的流行病学研究。
[1]唐曙明,方红辉,杨自华.产ESBLs肺炎克雷伯菌的耐药型与基因分型[J].中国现代医学杂志,2009,19(13):1653-1657.
[2]贾建军,刘蓬蓬.院内感染常见细菌分布及其药敏试验分析[J].青岛大学医学院学报,2008,44(1):61-63.
[3]傅冷西,傅蔷,陈丽锦,等.大肠埃希菌和肺炎克雷伯菌耐药性和耐药质粒谱分析[J].中国病原生物学杂志,2010,5(1):11-13.
[4]马丽萍.产超广谱β-内酰胺酶肺炎克雷伯菌和大肠埃希菌药敏分析[J].检验医学与临床,2011,8(5):526-527.
[5]曹亚.实用分子生物学操作指南[M].北京:人民卫生出版社,2003:31-33.
[6]HA WKEY P M.The growing bur den of anti microbial resis-tance[J].J Anti micr ob Che mot her,2008,62:i1-i9.
[7]扈新花,陈维忠,许成芳,等.ICU产ESBLs大肠埃希菌耐药性及基因型分析[J].青岛大学医学院学报,2011,47(5):413-415.
[8]HARADA S,ISHII Y,YAMAGUCHI K.Extended-spectrum beta-lacta mases:implications for the clinical laboratory and therapy[J].Korean J Lab Med,2008,28(6):401-412.
[9]陆喜颜,邓文喻.产ESBLs的大肠埃希菌和肺炎克雷伯菌的检测和耐药性分析[J].国际医药卫生导报,2012,18(4):535-539.
[10]李云慧,翟如波,张昊,等.产超广谱β-内酰胺酶大肠埃希菌耐药性及基因型分布[J].国际检验医学杂志,2012,33(15):1822-1825.
[11]郑见康.产超广谱β-内酰胺酶细菌耐药性调查[J].中华医院感染学杂志,2007,17(9):1150-1151.
[12]张志坚,郭小兵,张钦宪.产ESBL大肠埃希菌质粒谱与耐药性分析[J].第四军医大学学报,2008,29(21):1970-1972.
[13]王贺,杨启文,徐英春,等.产超广谱β-内酰胺酶肠杆菌科对头孢美唑等抗菌药物的体外敏感性分析[J].中华医院感染学杂志,2008,18(6):850-854.
[14]ALAA H,CHARRAKH A L,SA MIRA Y,et al.Occurrence and detection of extended-spectr u m-lacta mases in Klebsiella isolates in Hilla,Iraq[J].African J Biotechnol,2011,10(4):657-665.
[15]张玲,黄留玉.革兰阴性杆菌产超广谱β-内酰胺酶研究进展[J].中华医院感染学杂志,2008,18(6):897-900.
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
国外医药(抗生素分册)(2016年3期)2016-07-12
系统工程与电子技术(2016年2期)2016-04-16
中华灾害救援医学(2015年7期)2016-01-07
中国光学(2015年1期)2015-06-06
现代检验医学杂志(2015年2期)2015-02-06
海岸工程(2014年4期)2014-02-27
现代检验医学杂志(2014年1期)2014-02-06
现代检验医学杂志(2014年4期)2014-02-02
草食家畜(2012年2期)2012-03-20
