
九连山自然保护区毛红椿天然林邻体干扰效应
2013-11-06 05:09杨清培付方林梁跃龙唐培荣刘足根郭晓燕
江西农业大学学报 2013年4期
杨清培,付方林,张 露*,梁跃龙,唐培荣,刘足根,郭晓燕
(1.江西农业大学 园林与艺术学院,江西 南昌 330045;2.江西九连山国家级自然保护区管理局,江西 龙南341701;3.江西省环境保护科学研究院,江西 南昌 330029)
毛红椿(Toona ciliata var.pubescens)为楝科香椿属植物,落叶大乔木,树高可达30 m,主要产于我国江西、福建、广东、四川、贵州和安徽等地[1]。其干形通直圆满,生长较迅速,木材花纹美观,素有“中国桃花心木”的美称[2]。因长期过度开发、天然更新缓慢等原因[3],毛红椿种质资源急剧下降,目前现存林分零星分布于自然保护区[4],是我国第一批二级保护的珍贵树种。近年广泛开展了资源状况、群落结构[3]、种间联结[5]及其空间遗传结构[6]、种子库与种子雨[7]等方面研究,但毛红椿自然更新困难的原因却知之甚少。
邻体干扰会强烈影响植物生长与种群更新,即相对某植物个体而言,其周边同种或他种个体(邻体)为争夺可利用资源而对其产生的负作用[8]。它对植物个体生长、种群动态和植物群落结构有着重要影响[9-11]。因此,植物邻体干扰引起了生态学工作者的极大关注,并成为植物种群生态学研究的热点之一[12-15]。那么邻体干扰是否及如何影响毛红椿自然更新的呢?
为此笔者提出了以下假设:在天然更新过程中,毛红椿的个体越小,它受到干扰的有效距离越宽,干扰物种就越多,受到邻体干扰的强度越大。为了验证以上假设,本文在江西九连山国家级自然保护区,选取典型毛红椿天然林为对象,开展邻体植物对不同径级毛红椿基株干扰的有效距离、干扰来源和干扰强度等方面的研究。研究结果对揭示毛红椿群落更新困难的原因,对毛红椿群落组建机制、人工造林及空间分布格局等具有重要参考意义。
1 材料与方法
1.1 研究区自然概况
研究区位于江西国家级九连山自然保护区内,地理位置为 N24°29'~24°38',E114°22'~14°31'。保护区年平均气温16.4℃,年平均日照1 069.5 h,年平均降水量2 155.6 mm,年平均蒸发量790.2 mm,年平均相对湿度为87%,2—9月为雨季,10—1月为旱季。保护区森林覆盖率达94.7%,核心区森林覆盖率98.2%。土壤类型为山地黄红壤,厚度一般1 m左右,土体中砾石较多,地表有厚约2~4 cm枯枝落叶层,其下为暗灰色腐殖质层[16]。本区地带性植被是常绿阔叶林,但也有植被类型主要有亚热带常绿阔叶林,但现分布有少量的落叶阔叶林。毛红椿群落主要分布冷水坑、虾公塘、大丘田等处[7]。
1.2 研究方法
1.2.1 样方调查 在全面踏查的基础上,选取毛红椿天然林作为研究样地,海拔410~420 m,坡向为南坡,样地大小分为40 m×40 m,对样地内所有乔木(胸径≥5 cm)进行每木调查,内容包括:植株的坐标、胸径、树高、枝下高和冠幅等。
1.2.2 数据处理 (1)物种重要值。它反映某种群在群落中的生态重要程度[17],其计算式为:

式①中,RA为相对多度,即某物种多度与群落内所有物种多度之和的比值(%),RF为相对频度,即某物种频度与群落内所有物种频度之和的比值(%),RD为相对显著度,即某物种显著度(断面积)与群落内所有物种显著度之和的比值(%),RH为相对高度,即某物种高度与群落内所有物种高度之和的比值(%)。
(2)植物空间分布。根据野外调查的每株植物的坐标数据,绘制植物分布图(图1)。并由②式计算出所有植物与毛红椿基株之间距离。

式②中,dij为邻体i与基株j之间的距离(m);xi,yi分别为邻体i在样地中的x、y坐标值,xj,yj分别为基株j在样地中的x、y坐标值。
(3)邻体干扰。采用张跃西[10]提出的邻体干扰强度模型来计算干扰强度的大小,该值越大,受干扰越强烈。其计算式为:

式③中,NIij为邻体i对基株j的干扰强度;Di为邻体的胸径(cm);Dj为基株j的胸径(cm);dij同②式。
(4)有效干扰距离。采用干扰距离逐步扩大法和分段拟合法[13],计算有效干扰距离。

图1 毛红椿空间分布与邻体干扰分解Fig.1 Spatial distribution of Toona ciliata var.pubescens and interference zone for objective tree

图2 邻体对毛红椿有效干扰距离(径级为20~30 cm)Fig.2 Effective inference distance form neighbor trees to Toona ciliata var.pubescens
假定所有对某基株产生干扰作用的树木都在以基株为中心的圆形区域S(S=π×r2)内,故以基株为圆心,将S划分为n个等距圆环(图1)。本文Δr=rk+1-rk=2 m(k=1、2、…、10)。离基株最近0~2 m以内定义为r1,2~4 m为r2,以此类推,18~20 m为r10,共得10个圆环。则圆环k内所有邻体对某基株的干扰强度计为NIk,并可通过式(3)计算而得。同时,邻体对毛红椿基株的干扰强度随距离增大而下降趋势,而且一定距离内下降较快,而后下降变缓。于是,采用分段拟合的方法[13],计算得到下降拐点,这点对应的距离即为有效干扰距离(图2)。

式④中,NIk为第k圆环内邻体(同种或他种)干扰强度,n为第k圆环内邻体数目。
(5)累积干扰强度。林木个体间的干扰具有可加性[13,18]。根据式④得到的有效干扰距离,将所有邻体(同种或他种)干扰强度相累加分别得到种内干扰、他种干扰和总的邻体干扰强度。
2 结果与分析
2.1 毛红椿群落与种群特征基本特征
毛红椿群落乔木层中共调查到植物26种,隶属12科18属。重要值前10位的依次为毛红椿(Toona ciliata var.pubescens)、闽楠(Phoebe bournei)、毛竹(Phyllostachys edulis)、华南桂(Cinnamomum austrosinense)、皂荚(Gleditsia sinensis)、红皮树(Styrax suberifolius)、乌楣栲(Castanopsis jucunda)、野黄桂 (Cinnamomum jensenianum)、青冈(Cyclobalanopsis glauca)、山桐子(Idesia polycarpa),这些物种重要值占80.95%。整个群落密度为1 156株/hm2,其中毛红椿密度最大,达319株/hm2(表1),胸径在20 cm<D≤30 cm的个体数量最多,占62.75%,而小植株(D≤10 cm)仅占3.92%,可见,此毛红椿群落处在发育盛期,毛红椿种群最优势,但已属衰退种群(图3),其他常绿树种已大量侵入,且生长良好。

图3 毛红椿种群密度及径级分布Fig.3 Diameter distribution of sample trees of Toona ciliata var.pubescens
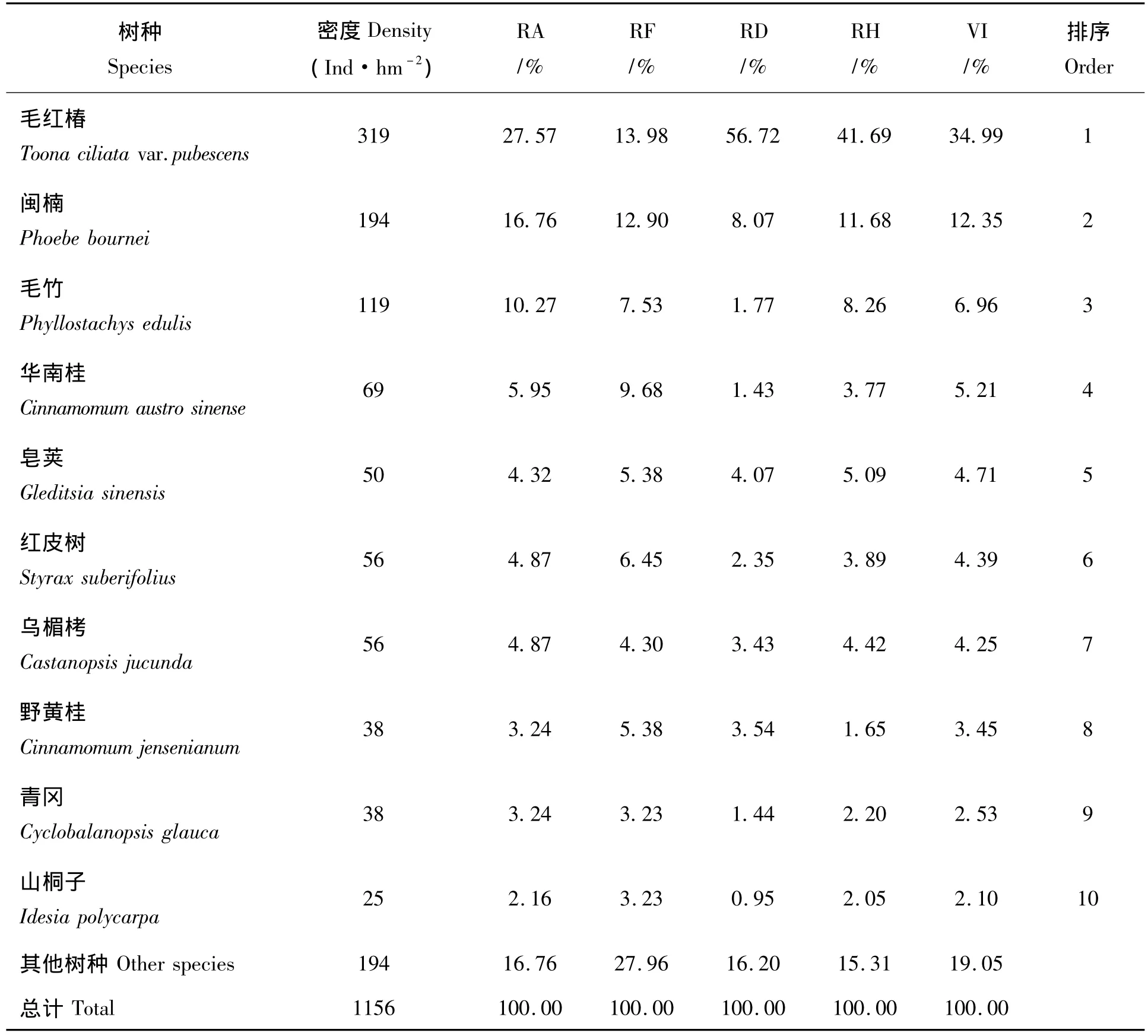
表1 毛红椿天然林物种组成及主要种群重要值Tab.1 Main species and its important value population in Toona ciliata var.pubescens forest
2.2 邻体干扰有效距离
由表2可知,基株胸径越小,其受邻体干扰的有效距离就越宽。5个径级(D≤10 cm、10<D≤20 cm、20<D≤30 cm、30 <D≤40 cm、D >40 cm),毛红椿受邻体干扰的有效距离分别为 10、8、6、6、6 m,只有当胸径超过20 cm后,毛红椿所受干扰的有效距离基本维持在6 m。可见,较小基株受邻体干扰的有效距离较宽。
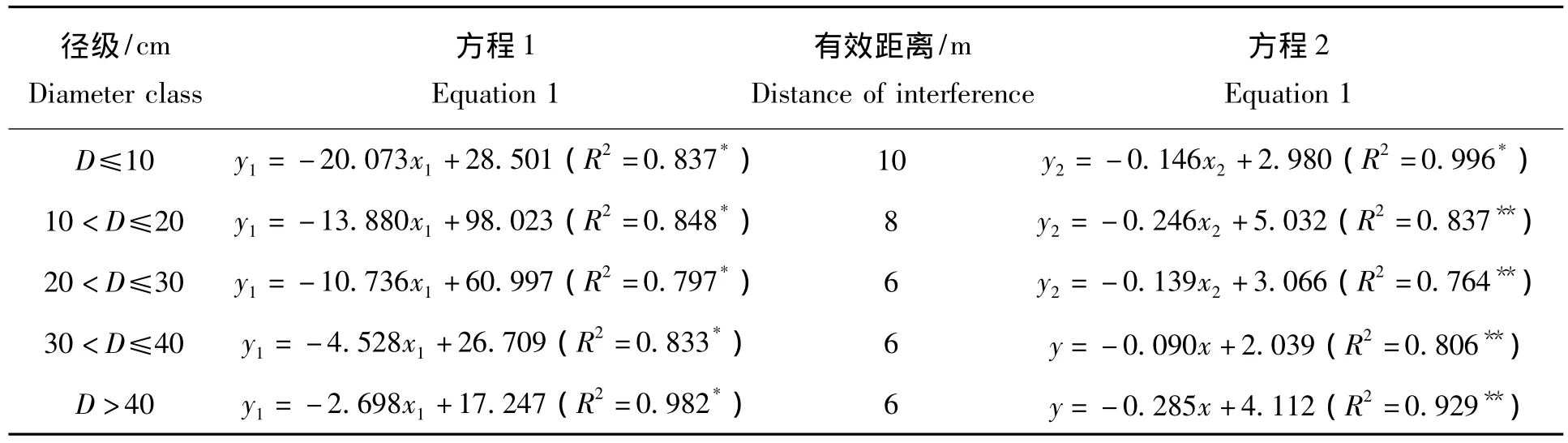
表2 邻体对不同径级毛红椿基株干扰的有效距离Tab.2 The effective interference distance of neighbors to Toona ciliata var.pubescens
2.3 毛红椿基株受邻体干扰强度与来源
利用有效干扰距离,计算不同径级毛红椿基株受种内和主要伴生树种的干扰强度(表3)。
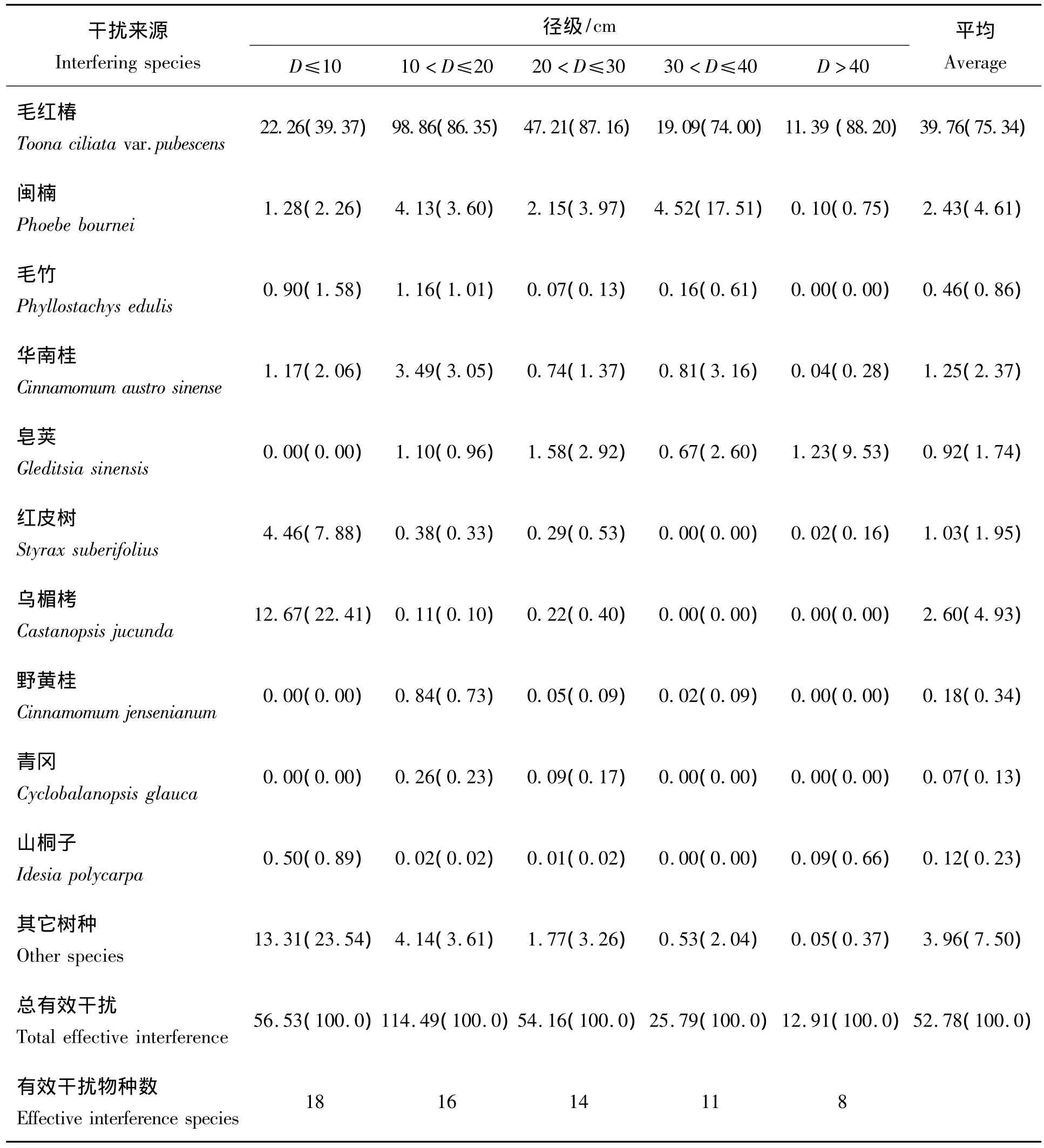
表3 毛红椿与主要伴生树种对不同径级基株的干扰强度Tab.3 Inference intensities to objective trees in different sizes from Toona ciliata var.pubescens and other accompany tree species
从表3可以看出,不同径级毛红椿基株受干扰的邻体植物种类不同。小于10 cm的基株受到的有效干扰为56.53,主要来自种毛红椿、乌楣栲、红皮树、闽楠、华南桂等;10~20 cm的基株受到的有效干扰为114.49,主要来自种毛红椿、闽楠、华南桂、毛竹等,20~30 cm的基株受到的有效干扰为54.16,主要来自种毛红椿、闽楠、皂荚、华南桂等;30~40 cm的基株受到的有效干扰为25.79,主要来自种毛红椿、闽楠、华南桂、皂荚等;大于40 cm基株受到的有效干扰为12.91,主要来自种毛红椿、皂荚等。可见除小个体(D≤10 cm)外(从样地调查看,只有较为稀疏的地方才有活小个体),邻体干扰强度与基株胸径之间关系可用指数函数(y=ae-bx)拟合(图3)。可见,毛红椿基株胸径越小,邻体有效干扰就越大;基株胸径越大,邻体有效干扰就越小。这可能毛红椿自然更新困难的原因之一。

图4 干扰强度与毛红椿胸径的关系(A:总邻体干扰;B:种内邻体干扰)Fig.4 Regression equations between Toona ciliata var.pubescens individual size and disturbance index of adjacent trees
另外,毛红椿基株受到的干扰主要来自种群内部的干扰,尤其是胸径超过10 cm后,种内干扰占全部干扰的比例超过70%(74.00% ~88.24%),这可能与这里毛红椿群落发育阶段(盛期),毛红椿最为优势有关(表1),但对小径级毛红椿(D≤10)来说,它所受干扰除来自种内外,还有60.63%干扰来自其它物种(如乌楣栲22.41%、红皮树7.88%、闽楠2.26%、华南桂2.06%等),这可能是毛红椿种群很难自然更新的原因。
3 结论与讨论
种群更新依赖于植物个体的生长状况,而植物个体的生长不仅取决于自身的遗传特性,而且取决于环境质量(如:光、热、水、土壤等)、植物对环境资源的利用能力、邻体植物对它的干扰状况等[13],但每个个体并非与群落中的所有个体争夺可利用资源,而是和它周围一定范围内的植株相互竞争,即存在一个有效竞争范围,这一范围植物空间分布格局和植物个体大小有关[12]。秦岭太白山国家森林公园和太白山国家级自然保护区境内太白冷杉(Abies sutchuenensis)天然林的太白冷杉所受有效干扰距离为5~9 m[13];黑龙江省凉水自然保护区红松天然林中红松受邻体干扰的有效距离为5~6 m[18]。本文研究发现毛红椿天然林中毛红椿基株受邻干扰的有效距离为6~10 m,基株个体越小,邻体干扰的有效距离就越宽,但胸径超过20 cm后,其有效干扰距离稳定在6 m,这与宋丁全[19]对光皮桦(Betula luminifera)的取样距离相似。与笔者提出的毛红椿个体越小,它受到干扰的有效距离就越宽的假设的一致。
群落发育可分为初期、盛期和末期3个阶段,不同植物群落的组成和种间关系差异较大。随群落发育,建群种从逐渐占优势到最后衰退而退出群落。本研究中毛红椿群落处在发育盛期,建群种毛红椿重要值最大,生态作用最强,但群落组成看,许多常绿植物纷纷侵入,并占有相当比例。毛红椿受到的竞争干扰除来自种内外,幼树、小树还面临其它物种,尤其是常绿植物的竞争干扰(表3)。这验证了毛红椿个体越小,就会有越多物种对它产生干扰的假设,这可能又是毛红椿自然更新困难的原因之一。
群落中树木径级的不同,其周围植物的分布格局也有很大的差别,小树部边邻体植物较大,且距离较近,而大树旁边邻体植物较小,这样小基株受到邻体干扰强度远远大于大基株[18]。毛红椿是落叶性的阳性植物,当基株径级较小时,如果周围的植株过近,可能因邻体的干扰(遮荫)作用而死亡,由于长期自然选择和适应,毛红椿只能更新于相对空旷或稀疏的林下,这可能是样地内毛红椿小树(D≤10 cm)较少(仅占3.92%)的原因,但随后(10~20 cm)毛红椿受邻体干扰会遽然增加,经过激烈竞争长成大树(30~40 cm)以后,受邻体干扰强度才下降。这基本回答了毛红椿个体越小,受到邻体干扰的强度就越大的假设。这可能也是毛红椿自然更新困难的原因之一。
总之,以九连山自然保护区毛红椿天然林为对象,采用干扰距离逐级扩大法,通过邻体对不同径级毛红椿基株干扰的有效距离、干扰来源和干扰强度等方面的研究与分析,发现群落中较小毛红椿基株受到邻体干扰的有效距离宽,干扰邻体种类多和干扰强度大,这可能是毛红椿天然更新困难的原因。
[1]中国科学院中国植物志编辑委员会.中国植物志(第43卷第3分册)[M].北京:科学出版社,1997:42.
[2]张露,郭联华,杜天真.遮荫和土壤水分对毛红椿幼苗光合特性的影响[J].南京林业大学学报:自然科学版,2006,30(5):63-66.
[3]刘军,陈益泰,罗阳富,等.毛红椿天然林群落结构特征研究[J].林业科学研究,2010,23(1):93-97.
[4]刘信中,肖忠优,马建华.江西九连山自然保护区科学考察与森林生态系统研究[M].北京:中国林业出版社,2002.
[5]付方林,张露,杨清培,等.毛红椿天然林优势种群的种问联结性研究[J].江西农业大学学报,2007,29(6):982-987.
[6]刘军,陈益泰,孙宗修,等.基于空间自相关分析研究毛红椿天然居群的空间遗传结构[J].林业科学,2008,44(6):45-52.
[7]黄红兰,张露,廖承开.毛红椿天然林种子雨、种子库与天然更新[J].应用生态学报,2012,23(4):972-978.
[8]Weiner J.Neighbourhood interference amongst Pinus rigida individuals[J].Journal of Ecology,1984,72,183 -195.
[9]Grace S L,Platt W J.Neighborhood effects on juveniles in an old - growth stand of longleaf pine,Pinus palustris[J].Oikos,1995,72(1):99 -105.
[10]张跃西,钟章成.木本植物邻体干扰研究进展[J].生态学杂志,1999,18(2):55-59.
[11]潘攀,李荣伟,覃志刚,等.杜仲人工林中的邻体干扰[J].四川林业科技,2002,23(2):15-18.
[12]段仁燕,王孝安.植物邻体干扰的研究范畴、热点及意义[J].西北植物学报,2004,24(6):1138-1144.
[13]段仁燕,王孝安.太白红杉种群邻体范围与邻体竞争强度的研究[J].西北植物学报,2004,24(12):2335-2340.
[14]Ramseier D,Weiner J.Competitive effect is a linear function of neighbour biomass in experimental populations of Kochia scoparia[J].Journal of Ecology,2006,94:305 - 309.
[15]陈德祥,班小泉,李意德,等.热带柚木(Tectona grandis L.f.)人工林中不同邻体干扰强度下林木CO2气体交换特征[J].生态学报,2008,28(9):4059 -4068.
[16]朱祥福,林宝珠,周华.九连山碟斗青冈群落结构及小气候特征分析[J].江西林业科技,2006,4(5):4-9.
[17]王伯荪,余世孝,彭少麟,等.植物群落学实验手册[M].广州:广东高等教育出版社,1996:6.
[18]国庆喜,杨光.红松天然种群邻体影响半径[J].应用生态学报,2006,17(12):2302-2306.
[19]宋丁全,姜志林,郑作孟.光皮桦群落邻体干扰指数的研究[J].生态学杂志,2002,21(3):15-17.
猜你喜欢
广西林业科学(2022年4期)2022-11-07
当代水产(2022年8期)2022-09-20
林业调查规划(2022年3期)2022-06-13
昆明医科大学学报(2022年2期)2022-03-29
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
食品安全导刊(2021年20期)2021-08-30
林业科技情报(2021年2期)2021-07-13
园艺与种苗(2021年3期)2021-05-26
河南科学(2020年3期)2020-06-02
