
干热河谷主要造林树种生长季树干液流的时滞特征
2013-11-05 06:40王小菲孙永玉张春华
江西农业大学学报 2013年3期
王小菲,孙永玉 ,李 昆,张春华,李 彬,侯 锐
(1.中国林业科学研究院 资源昆虫研究所,云南 昆明650224;2.西南林业大学 环境工程与科学院,云南 昆明650224)
干热河谷是我国西南地区对气候干燥炎热的河谷之称谓,区域内光、热资源丰富,气候炎热少雨,植被稀少,水土流失严重,生态十分脆弱[1-2],干热河谷区限制生态系统发生与发展最关键的要素是水分[3],认识干热河谷主要造林树种的耗水规律对该区植被恢复和管理有重要的科学意义。
树干液流其时滞现象指的是以液流反映的树木蒸腾往往滞后于树冠的实际蒸腾[4,7]。许多研究发现基于树干液流测定值来计算冠层气孔导度、冠层蒸腾或者解释其与环境因子的关系时,如果不考虑时滞,误差会很大[8,10]。国内外已有许多学者对一些地区常见树种及造林树种其时滞特征得出了许多研究结果,如王华[11]等对北京常见树种银杏(Ginkgo biloba)、七叶树(Aesculus chinensis)、玉兰(Magnolia denudata)、刺槐(Robinia pseudoacacia)、油松(Pinus tabulaeformis)、雪松(Cedrus deodara)生长季树干液流的时滞特征研究得出其树干液流均滞后于总辐射,而提前于水汽压亏缺。目前,由于对热河谷地区树干液流的时滞现象尚未见报道。因此,本文基于热扩散式探针法,通过对干热河谷主要造林树种山合欢(Albizzia kalkora)、印楝(Azadirachta Indica)、大叶相思(Acacia auriculaeformis)、新银合欢(Leucaena leucocephala)、赤桉(Eucalyptus camaldulensis)、柠檬桉(Acacia confusa)的树干液流监测,采用气象因子与液流间的时间序列的错位对应分析方法,分析该地区树木时滞效应的规律及其影响因素,揭示树干液流对气象因子的响应规律,旨在为计算蒸腾和冠层气孔导度提供依据,同时也为干热河谷区植被建设及恢复提供基础资料。
1 研究区概况
研究地点位于国家林业局云南元谋荒漠化生态系统定位观测站山地人工林试验场,该站位于云南省元谋县,地理位置101°51'41″N,25°40'18″E,海拔1 180 m,属典型的金沙江干热河谷区。该区气候异常干、热,且干湿季分明。年平均气温为21.9 ℃,最冷月均温为14.9 ℃(12 月),最热月均温为27.1 ℃(5 月),最高气温达42 ℃;年平均降雨量为613.8 mm,6 -10 月为湿季(雨季),11 月至翌年5 月为干季(旱季),湿季降雨量占年降雨量的90%以上;年均相对湿度为54%,年均日照时数为2 630.4 h,全年无霜期360 ~365 d,年平均干燥度4.0。
2 材料与方法
2.1 试验材料
选择具有代表性的,生长良好,无被挤压的12 年生山合欢、印楝、大叶相思、新银合欢、赤桉、柠檬桉为研究对象,边材面积测量方法参见文献[3]。测树因子如表1。
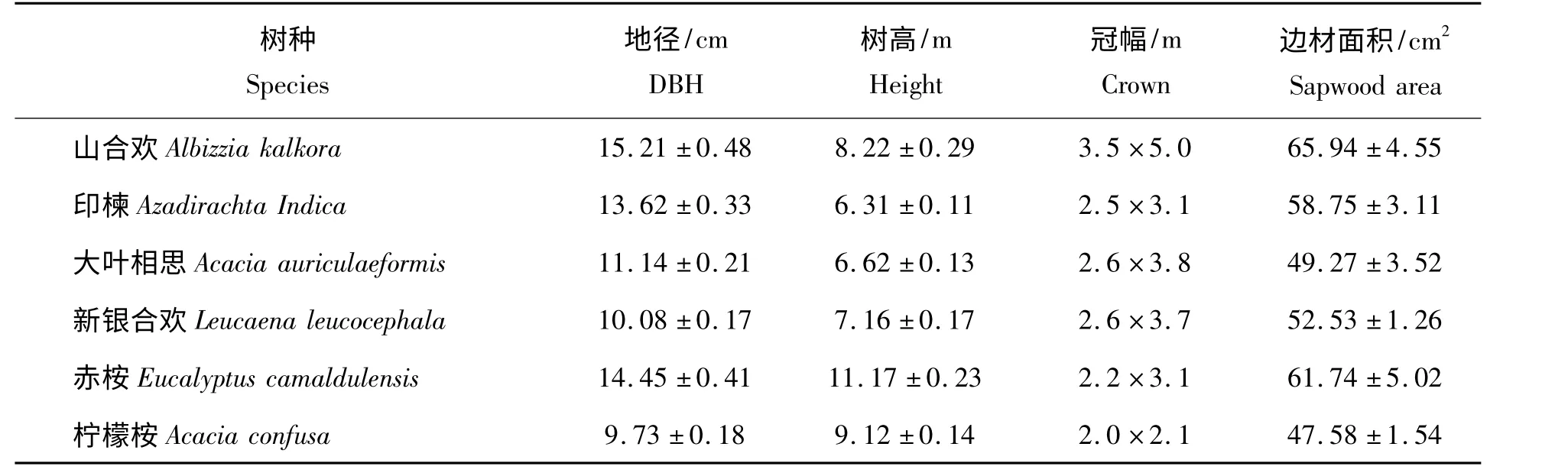
表1 监测样木树形因子Tab.1 Characteristics of the sampling trees for sap flow measurements
2.2 试验方法
2.2.1 树干液流的测定 采用Granier 热扩散式探针法对各样木于2012 年生长季(5—8 月)树干液流进行连续监测。根据所选样木边材部分的厚度确定探针规格,地径小于10 cm 的使用TDP -20 型探针,大于10 cm 的使用TDP-30 型探针,探针直径为1.32 mm。由于大叶相思、印楝、山合欢等第一侧枝比较低,为监测整株液流动态,故将探针均安装在样木树干地径处,并于样木南、北向各装一个探头,所有传感器均与数据采集仪(Grant 2 020 Series)相连,安装好探针后用铝箔在探头的外部围住,以防止太阳辐射对探针的影响;数据采集间隔时间设为10 s,每10 min 进行平均值计算并储存数据。
采用Granier[12]的公式来计算液流速率,即:Vs =0.011 9 × [(ΔTm- ΔT)/ΔT]1.231,式中,Vs 为瞬时液流速率(cm/s),ΔTm 为无液流时加热探针与参考探针之间的最大温差,ΔT 为瞬时温差。而液流通量(F)是指单位面积边材单位时间内通过液流的体积,其计算式为:F=Σni=1Vi×As×Δt,式中,n 为每日取样次数,Vi 为第i 次取样时的液流速率(cm/s),As 为边材面积(cm2),Δt 为取样间隔时间(s)。
2.2.2 气象因子的测定 样地内设有自动气象站,实时监测空气温度(℃)、湿度(%)、风速(m/s)、风向、太阳辐射(w/m2)等气象参数,所有传感器均与数据采集仪相连,并与液流测定同步。水汽压亏缺计算方法[12]:
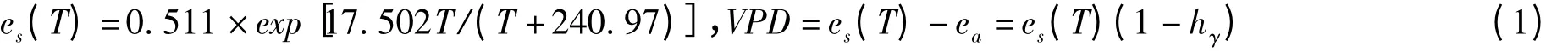
式中,es(T)为T 大气温度下的饱和水汽压(kPa);T 为大气温度(℃);ea为周围气体水汽压(kPa);hγ为相对湿度(%);VPD 为水汽压差(kPa)。
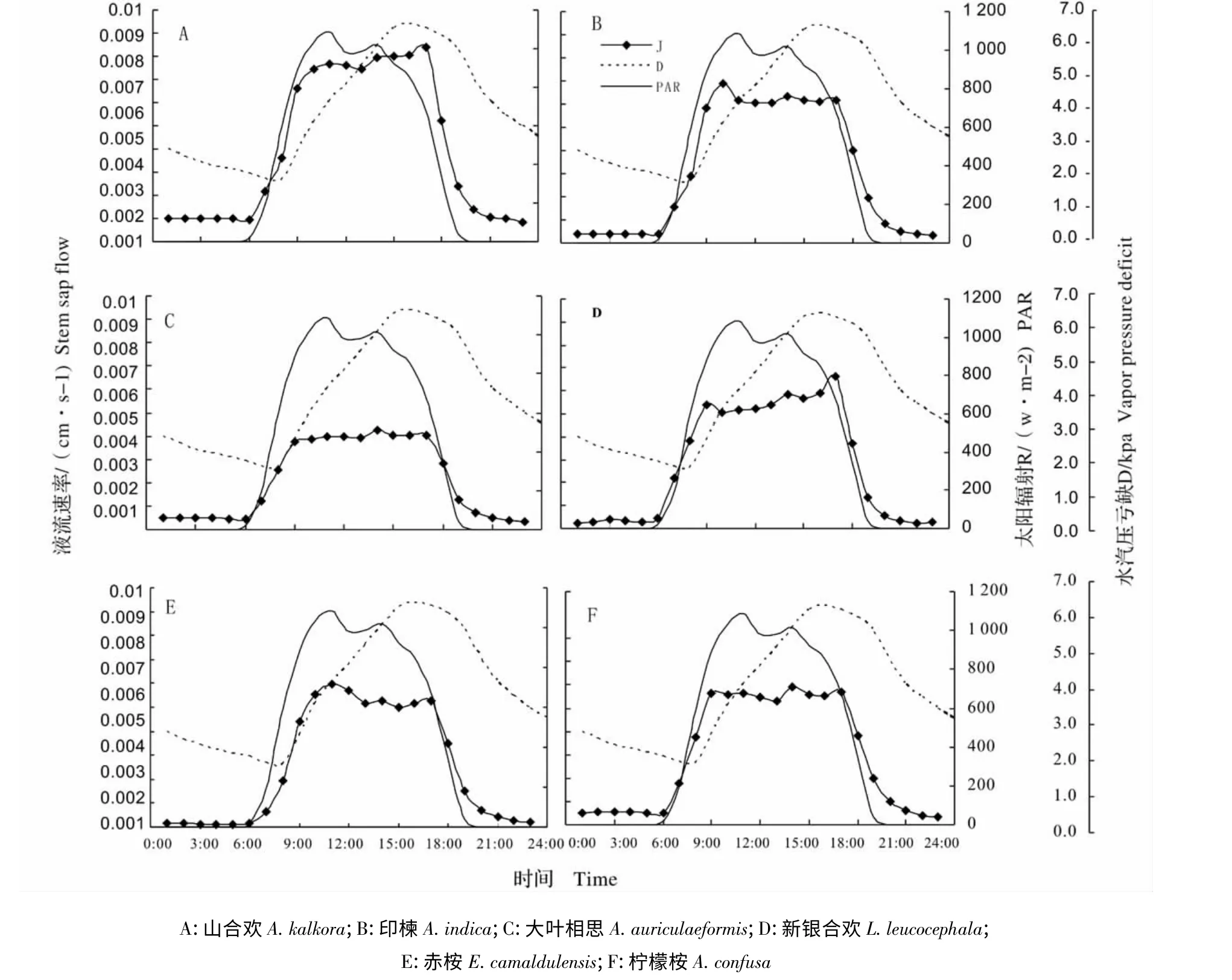
图1 8 月干热河谷主要造林树种晴天树干液流速率与太阳辐射、水汽压亏缺的日动态Fig.1 Diurnal changes of stem sap flow (J),PAR and vapor pressure deficit (D),for main afforestation tree species in August
2.2.3 时滞的计算方法 使用错位对比法,即对每种样树的平均液流速率分别与总辐射和水汽压亏缺逐次按10 min 进行错位移动,分析错位移动后数据的相关关系,当相关系数达到最大值时,所对应的错位时间即为液流对总辐射或者水汽压亏缺的实际时滞,具体请参见文献[9 -10]。
2.2.4 夜间水分补充量计算方法 夜间水分补充量计算公式:W=Σ (Js·As·t),式中,W 为夜间水分补充量(g);Js 为夜间光合有效辐射为0 时的液流速率(cm/s);As 为边材面积(cm2);根据数据采集频度,Js 是10 min 液流速率的平均值,因此时间t 为600 s。
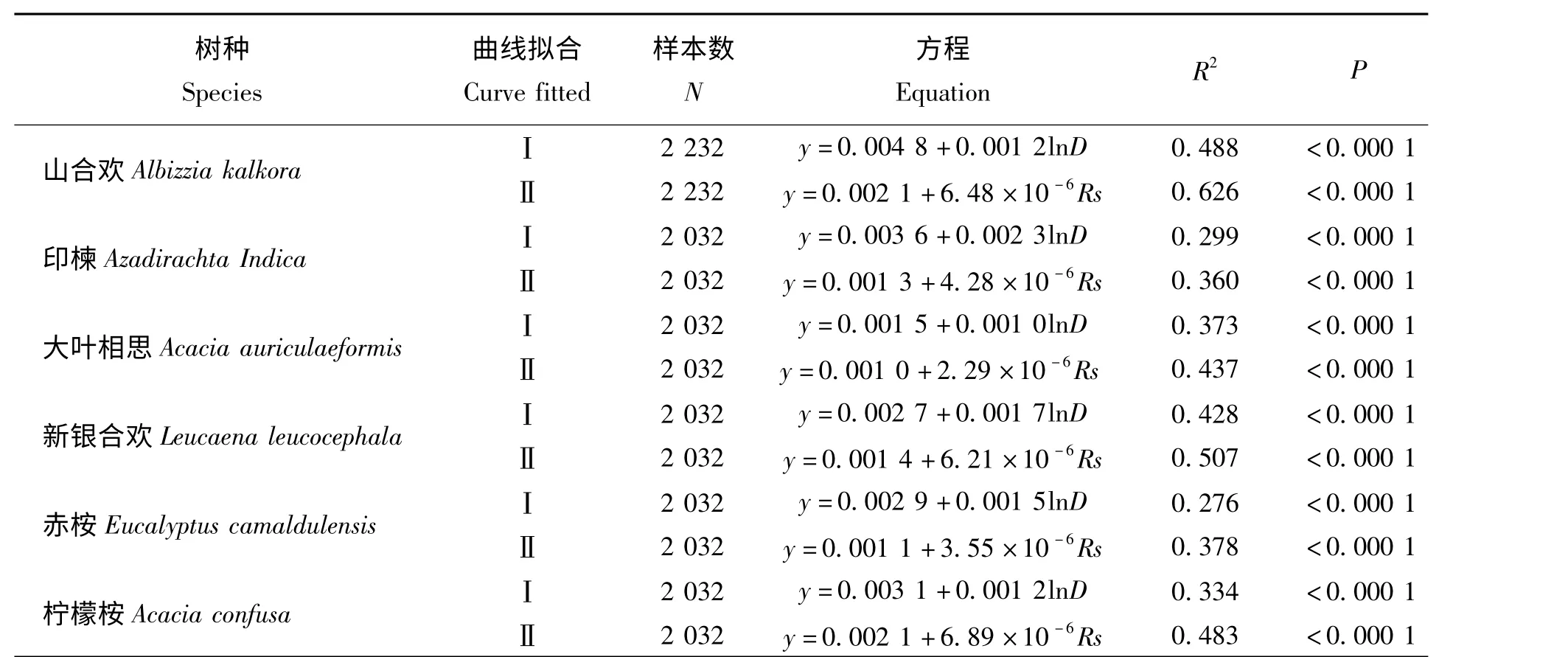
表2 干热河谷主要造林树种白天小时尺度上树干液流密度与水汽压亏缺(Ⅰ)、太阳辐射(Ⅱ)间的曲线拟合Tab.2 Curve fitted between sap flow (y)and vapor pressure deficit (D)(Ⅰ),as well as total radiation (Rs)(Ⅱ)for main afforestation tree species on the daily hourly scale
2.3 数据处理
数据使用Excel2003、SPSS19.0 等统计分析软件处理。
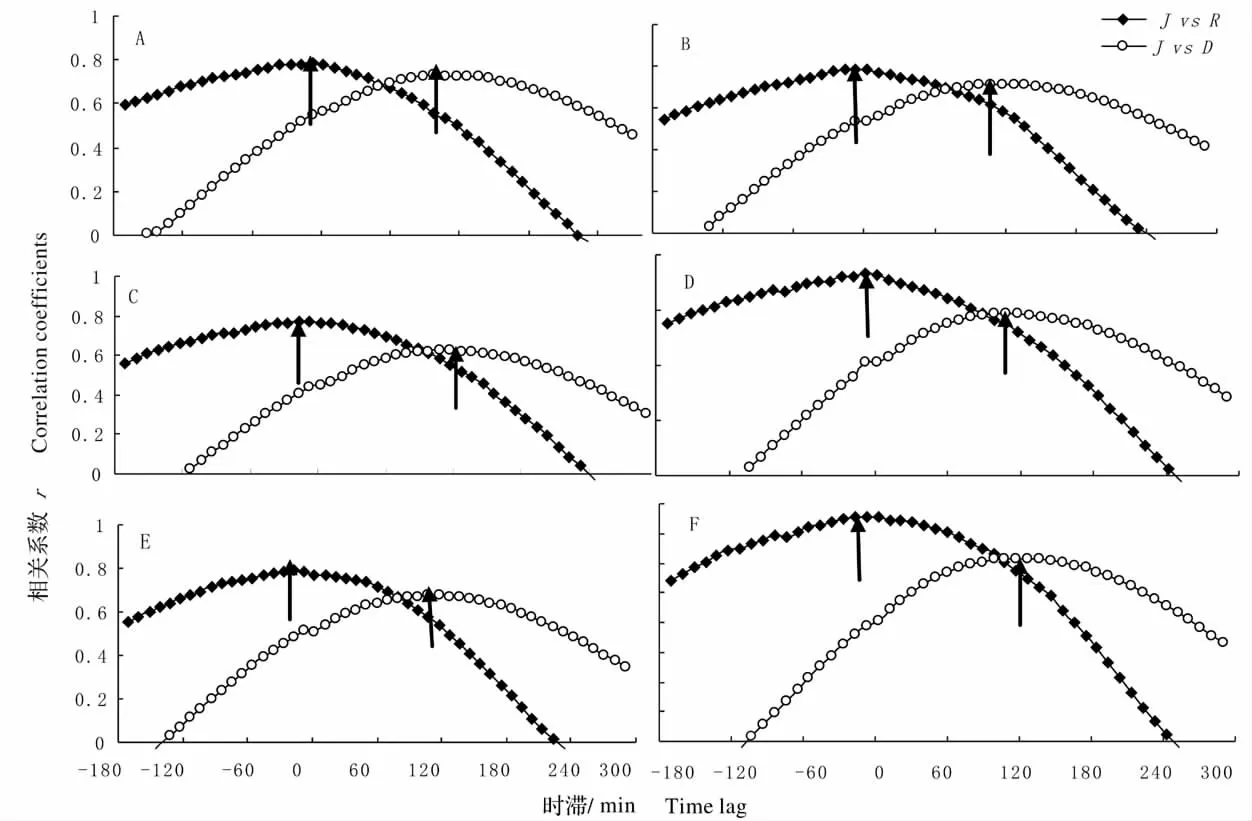
图2 5 ~8 月干热河谷主要造林树种树干液流速率均值(J)与太阳辐射(R)和水汽压亏缺(D)之间的相关系数Fig.2 Correlation coefficients between mean stem sap flow and total radiation(R),as well as and vapor pressure deficit (D)for main afforestation tree species from May to August in 2012
3 结果与分析
3.1 干热河谷造林树种树干液流与蒸腾驱动因子的关系
为直观表达时滞,选择8 月中晴天树木树干液流与水汽压亏缺、总辐射日变化进行作图(图1),图中可以看到,树干液流速率均表现出“昼高夜低”的昼夜变化规律,均呈现为“宽峰”曲线;树干液流速率曲线与太阳辐射基本保持一致;而与水汽压亏缺均存在明显的时滞。其中山合欢、印楝和新迎合欢液流速率上午与太阳辐射有很好的一致性,下午则滞后于太阳辐射。
表2 为干热河谷常见树种白天小时尺度上树干液流密度与水汽压亏缺、太阳辐射间的曲线拟合及相关性,表中可知,山合欢和新银合欢树干液流速率与太阳辐射相关性较高,即更依赖于太阳辐射的变化。
3.2 干热河谷树干液流与蒸腾驱动因子的时滞效应
使用错位对比法,对各种样树的平均液流速率分别与总辐射和水汽压亏缺逐次按10 min(计算时滞区间为180~300 min)进行错位移动并做相关分析,相关系数最大的时间差即为液流的时滞值(图2、图3),由图2,3 可知,树干液流均滞后于太阳辐射,并提前于水汽压亏缺,且树干液流与太阳辐射见最大相关系数(0.360 ~0.626)均高于树干液流与水汽压亏缺间的最大相关系数(0.267 ~0.488),同时,各造林树种其树干液流与太阳辐射的时滞范围(10 ~20 min)明显低于树干液流与水汽压亏缺的时滞范围(90 ~120 min),表明在日尺度上,树干液流很大程度依赖于太阳辐射的变化。
同时,从图3 中还可发现干热河谷6 个研究树种的种间时滞效应的差异较小,各树种树干液流与太阳辐射的时滞差在10 min 内,与水汽压亏缺的时滞差在30 min 内,表现出较好的一致性。
3.3 树干液流与蒸腾驱动因子间的时滞影响因素
表3 可知,干热河谷主要造林树种生长季树干液流与PAR 和VPD 之间的时滞长短与树木的地径、高度、冠幅、边材面积以及夜间水分补充量等的相关关系均不显著,可知该区所测树木液流与PAR 和VPD 之间的时滞受树形因子和夜间水分补充量的影响较小甚至没有。
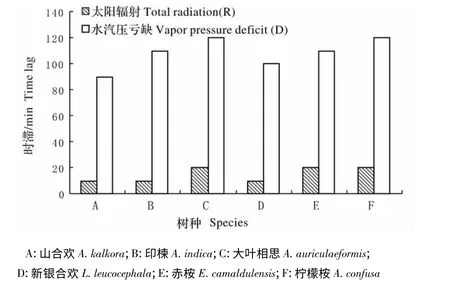
图3 树干液流与太阳辐射、水汽压亏缺之间的时滞Fig.3 Time lag between J vs R,J and D for main afforestation tree species
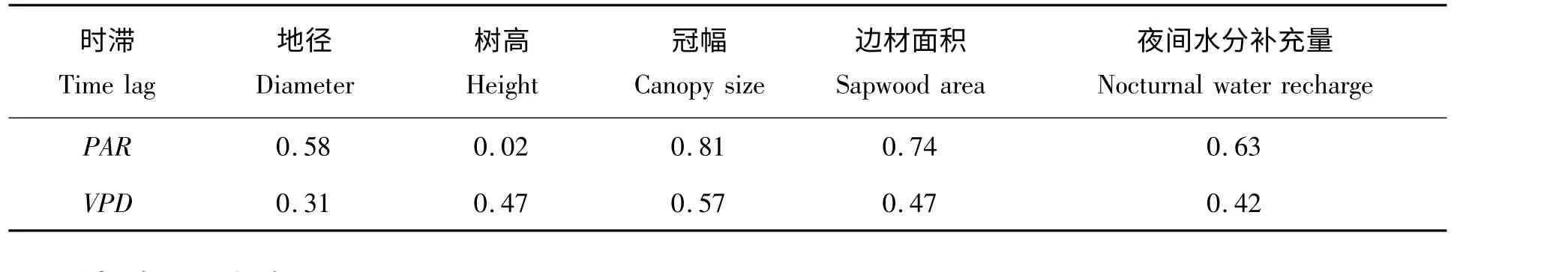
表3 干热河谷主要造林树种树干液流与水汽压亏缺、太阳辐射间的时滞与各树形因子、夜间水分补充量的相关关系(r)Tab.3 Correlations between time lag and tree features and nocturnal water recharge for several common urban tree species from April to September
4 结论与讨论
干热河谷主要造林树种在生长季树干液流速率与太阳辐射、水汽压亏缺存在明显的时滞,即呈现树干液流均滞后于太阳辐射,提前于水汽压亏缺。这与王华[12]、赵平[13]、Phillips N[7]& Kume T[8]等的研究结果一致。
各造林树种树干液流响应太阳辐射、水汽压亏缺变化的格局存在差异,树干液流与太阳辐射见最大相关系数(0.360 ~0.626)均高于树干液流与水汽压亏缺间的最大相关系数(0.267 ~0.488),树干液流与太阳辐射的时滞时间在10 ~20 min,明显低于与水汽压亏缺的时滞时间(90 ~120 min),可知树干液流与太阳辐射之间表现出很好的一致性,即在日尺度上,树干液流很大程度依赖于太阳辐射的变化。
本文所选6 个研究树种的种间时滞效应的差异较小,表现出较好的一致性,都依赖于太阳辐射的变化,这可能是所选树种都是阔叶乔木的缘故,王华等[9]对北京城区常见树种的研究发现,叶片较小的针叶树种油松(Pinus tabulaeformis)和雪松(Cedrus deodara)的树干液流变化比银杏(Ginkgo biloba)、七叶树(Aesculus chinensis)、玉兰(Magnolia denudata)等阔叶树木更依赖于水汽压亏缺的变化。
本研究表明,干热河谷主要造林树种生长季树干液流与PAR 和VPD 之间的时滞受树高、地径、冠幅、边材面积以及夜间水分补充量的影响较小甚至没有,这与王华等[11]对马占相思湿季的研究结果相一致,一般认为可能是环境因素、树干水分补充和水力导度等的作用掩盖了树形因子对时滞的影响[11]。
[1]李昆,曾觉民.金沙江干热河谷主要造林树种蒸腾作用研究[J].林业科学研究,1999,12(3):244 -250.[2]李昆,孙永玉.干热河谷植被恢复技术[M].昆明:云南科技出版社,2011:2 -8.
[3]钟祥浩.干热河谷区生态系统退化及恢复与重建途径[J].长江流域资源与环境,2000,9(3):376 -383.
[4]赵平,饶兴权,马玲,等.Granier 树干液流测定系统在马占相思的水分利用研究中的应用[J].热带亚热带植物学报,2005,13(6):457 -468.
[5]Schulze E D,Cermk J,Matyssek R,et al. Canopy transpiration and water fluxes in the xylem of the trunk of Larix and Piceatrees-A comPARison of xylem flow,porometer and cuvette measure ments[J].Oecologia,1985,66:475 -483
[6]Phillips N,Nagchaudhuri A,Oren R,et al.Time constant for water transport in loblolly pine trees estimated from time series of evaporative demand and stem sap-flow[J].Trees,1997,11:412 -419.
[7]Phillips N,Oren R,Zimmermann R,et al.Temporal patterns of water flux in trees and lianas in a Panaman -ianmoist forest[J].Trees,1999,14:116 -123.
[8]Kume T,Komatsu H,Kuraji K,et al.Less than 20 min time lags between transpiration and stem sap flow in emergent trees in a Bornean tropical rainforest[J].Agricultural and Forest Meteorology,2008,148:1181 -1189.
[9]Granier A,Loustau D,Breda N. A generic model of forest canopy conductance dependent on climate,soil water availability and leaf area index[J].Annals of Forest Sciences,2000,57:755 -765.
[11]王华,欧阳志云,郑华,等. 北京城区常见树种生长季树干液流的时滞特征[J]. 应用生态学报,2009(9):2111 -2117.
[12]王华,赵平,蔡锡,等.马占相思树干液流与光合有效辐射和水汽压亏缺间的时滞效应[J].应用生态学报,2008,19(2):225 -230.
[12]Granier A.Evaluation of transportation in a Douglas -fir stand by means of sap flow measurements[J]. Tree Physiology,1987(3):309 -320.
[13]赵平,饶兴权,马玲,等.马占相思(Acacia mangium)树干液流密度和整树蒸腾的个体差异[J]. 生态学报,2006,26(12):4050 -4058.
[14]吴芳,陈云明,于占辉.黄土高原半干旱去刺槐生长盛期树干液流动态[J].植物生态学报,2010,34(4):469 -476.
猜你喜欢
气候与环境研究(2022年4期)2022-08-15
干旱区地理(汉文版)(2022年3期)2022-05-30
小哥白尼(神奇星球)(2021年3期)2021-07-22
小哥白尼(神奇星球)(2021年4期)2021-07-22
沙漠与绿洲气象(2021年1期)2021-04-02
Zoological Research(2019年3期)2019-06-28
乡村地理(2018年4期)2018-03-23
中国质量与标准导报(2014年5期)2014-02-28
储能科学与技术(2014年5期)2014-02-27
储能科学与技术(2014年1期)2014-02-27
