
优雅蝈螽雄性附腺发育过程的超微结构
2013-10-28 05:12:24张小霞常岩林石福明周娜
河北大学学报(自然科学版) 2013年1期
张小霞,常岩林,石福明,周娜
(河北大学 生命科学学院,河北 保定 071002)
优雅蝈螽雄性附腺发育过程的超微结构
张小霞,常岩林,石福明,周娜
(河北大学 生命科学学院,河北 保定 071002)
利用透射电镜(TEM)技术研究了优雅蝈螽GampsocleisgratiosaBrunner von Wattenwyl雄性附腺不同发育时期的超微结构.结果表明:优雅蝈螽雄性附腺乳白长腺管和乳白短腺管在末龄若虫期已分化形成腺管,透明腺管在羽化之后形成腺管.优雅蝈螽雄性附腺在发育过程中,上皮细胞中的细胞器表现出由分散到集中和由少到多的趋势;腺管管腔中的分泌物由少到多,成熟期分泌物充满整个管腔.
直翅目;优雅蝈螽;雄性附腺;发育
昆虫雄性附腺形态不同,功能多样,其分泌物主要成分是蛋白质,是精液的主要成分[1-2].在雌雄交配过程中,雄性附腺分泌物对雌虫行为和生理的调节具有重要作用[3-4].目前昆虫雄性附腺的研究主要集中在形态结构、起源、发育、分泌物的生理生化特性和分泌物的功能等方面[1,5-7].
昆虫雄性附腺在显微结构、超微结构等方面具有种间差异性,即使是同种昆虫也可能有明显的区域特异性和时序性[6].优雅蝈螽雄性附腺发达,代表了螽斯类昆虫雄性附腺的结构特征,搞清其雄性附腺的结构和功能对深入了解螽斯类的生殖能力具有重要意义.冯晓丽等[8]对优雅蝈螽雄性附腺显微结构及其分泌蛋白特性等方面的研究做了报道.本文在了解优雅蝈螽雄性附腺显微结构及分泌蛋白特性的基础上仅对其附腺的发育过程进行研究,为阐明螽斯雄性附腺的功能奠定基础.
1 材料与方法
1.1试虫、试剂与仪器
研究材料为采自河北顺平的优雅蝈螽GampsocleisgratiosaBrunner von Wattenwyl成虫,室内虫笼饲养,每笼1雌1雄.室内产卵,越冬后的卵在室内孵化,饲养孵化的个体至末龄若虫及羽化后的成虫.
主要试剂:直翅目昆虫生理盐水(NaCl 6.8 g,MgCl2·6H2O 0.2 g,CaCl20.2 g,KCl 0.2 g,NaHCO30.15 g,葡萄糖7.7 g,H2O 1 000 mL)、DMP-30(2,4,6,-三(二甲氨基甲基)苯酚)、MNA(甲基内次甲基四氢苯二甲酸酐)、环氧树脂Epon812、DDSA(十二烷基琥珀酸酐)、四氧化锇、戊二醛、柠檬酸铅、醋酸铀.
主要仪器:Nikon YS2-H光学显微镜;Reichert-Jung超薄切片机;JEM-100SX透射电镜.
1.2雄性附腺不同发育时期超微结构的观察
1.2.1 取材
分别解剖优雅蝈螽雄性末龄若虫、羽化后1~9 d的成虫、羽化后10~15 d的成虫和性成熟的成虫,在直翅目昆虫生理盐水中分离雄性附腺的乳白长腺管、乳白短腺管和透明腺管.
1.2.2 固定
将分离的乳白长腺管、乳白短腺管和透明腺管立即投入体积分数为2.5%的戊二醛中,4 ℃固定1 h.在0.1 mol/L磷酸缓冲液(pH 7.4)中反复漂洗后,加入质量浓度为10 g/L的四氧化锇4 ℃固定2 h.
1.2.3 包埋
梯度脱水后,将材料转移到环氧树脂中包埋.
1.2.4 切片与观察
超薄切片机切片,厚度60 nm.将切片捞在200目(孔径约150 μm)或300目(孔径约100 μm)的铜网上,醋酸铀及柠檬酸铅双染色,透射电子显微镜观察并拍照.
2 结果
优雅蝈螽雄性附腺腺管的发育分化始于末龄若虫期.根据腺管上皮细胞的结构以及管腔中分泌物的多少,将其雄性附腺的发育过程划分为4个时期描述,即末龄若虫期、羽化后1~9 d的初生期、羽化后10~15 d的少生期、性成熟后的成熟期.
2.1乳白长腺管发育的超微结构
2.1.1 末龄若虫期的超微结构
末龄若虫期,乳白长腺管分化形成.管壁结构从外到内大致分为围膜、肌肉层、基膜、上皮细胞层4部分,上皮细胞附着在基膜上,基膜外环绕横纹肌纤维及富含微气管的围膜.末龄若虫期管壁上皮细胞排列不规则,未形成单层.上皮细胞中只有一些游离的核糖体(图1A),细胞顶部微绒毛稀疏且长短不一,排列不规则(图1B).腺管管腔小,管腔中未见分泌物(图1C).
2.1.2 初生期的超微结构
羽化后1~9 d的个体,乳白长腺管管壁上皮细胞与末龄若虫期相似,上皮细胞排列不规则.上皮细胞中含有一些游离的核糖体(图1D),靠近管腔的区域分布有少量分泌泡和粗面内质网(图1E).上皮细胞顶部微绒毛稀疏,管腔扩大但未见分泌物(图1F).
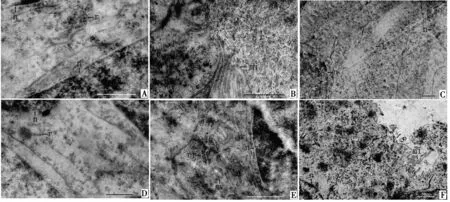
A.端部上皮细胞;B.微绒毛;C.腺管管腔;D.基部上皮细胞;E.端部上皮细胞;F.腺管管腔.er.粗面内质网;L.腺管管腔;mi.微绒毛;n.细胞核;r.核糖体;sv.分泌泡.标尺 = 1 μm.
2.1.3 少生期的超微结构
羽化后10~15 d的个体,乳白长腺管上皮细胞发育为单层,细胞中含有大量粗面内质网(图2A),靠近管腔的细胞顶部区域有线粒体分布(图2A).腺管管腔较初生期明显扩大,管腔中出现少量电子密度高的颗粒状分泌物(图2B).
2.1.4 成熟期的超微结构
性成熟后乳白长腺管管壁上皮细胞为单层,细胞层薄,排列规则,腺管管腔大.细胞基部及顶部邻近微绒毛的区域富含粗面内质网,细胞顶部区域含有大量分泌泡(图2C),大量核糖体游离或附着在内质网上(图2D).细胞顶部微绒毛长且不规则,周围分散有电子透明的空泡(图2D).腺管管腔中含有大量大小不等的分泌物,分泌物呈细颗粒状和电子致密的颗粒状物质(图2E).偶尔存在上皮细胞凋亡现象(图2F).上皮细胞的分泌方式以局部分泌为主(图2D).
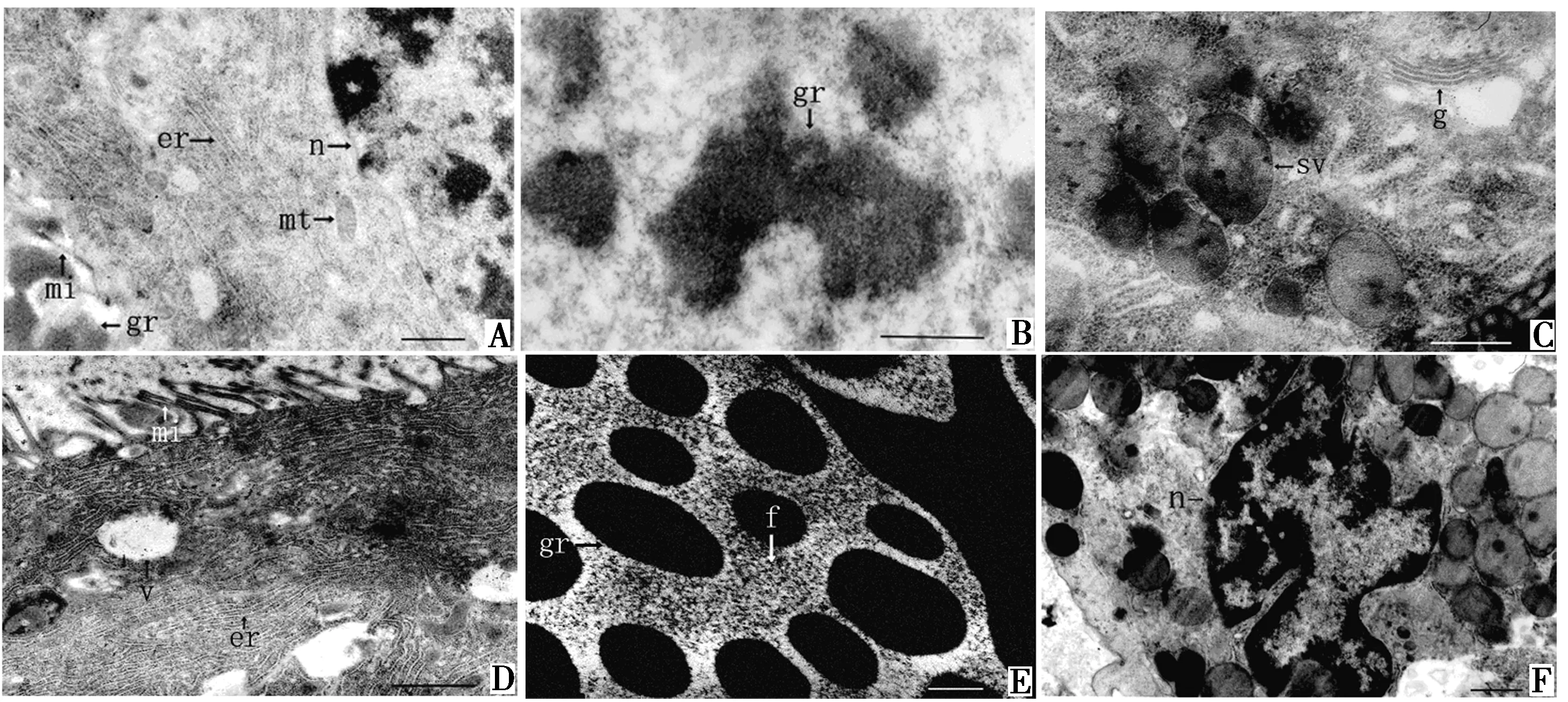
A.端部上皮细胞;B.分泌物;C.分泌泡;D.微绒毛 ;E.分泌物;F.细胞凋亡.er.粗面内质网;f.细颗粒物质;g.高尔基体;gr.颗粒物质;mi.微绒毛;mt.线粒体;n.细胞核;sv.分泌泡;v.电子透明空泡.标尺=1 μm.
2.2乳白短腺管发育的超微结构
2.2.1 末龄若虫期的超微结构
乳白短腺管的发育分化始于末龄若虫期,上皮细胞排列不规则,未形成单层(图3A).上皮细胞中有一些游离的核糖体.细胞顶部质膜形成微绒毛,稀疏且短(图3B),管腔中没有分泌物(图3A).
2.2.2 初生期的超微结构
刚羽化的个体,乳白短腺管上皮细胞排列不规则,细胞中含有一些游离的核糖体,细胞顶部临近管腔存在一些分泌泡,微绒毛较若虫期长,但排列仍不规则(图3C,D).腺管管腔中未见分泌物(图3D).
A.腺管整体结构;B.微绒毛;C.腺管管腔周围;D.腺管管腔.L.腺管管腔;mi.微绒毛;n.细胞核;sv.分泌泡.标尺= 1 μm.
2.2.3 少生期的超微结构
羽化10 d后的个体,乳白短腺管的上皮细胞形成单层,排列规则,细胞之间连接紧密,细胞顶部临近管腔的区域有一些粗面内质网(图4A).腺管中央有大小不等的分泌物(图4B).
2.2.4 成熟期的超微结构
性成熟后乳白短腺管的管壁上皮细胞层厚,腺管管腔相对较小.上皮细胞中富含粗面内质网(图4C),线粒体数量较多(图4D),细胞端部与腺管管腔之间有较多高尔基体及分泌泡分布(图4D, F).腺管管腔中富含2种分泌物:一种为细纤维状物质,形成网状空泡,中间镶嵌着电子致密的颗粒状物质(图4E);另一种为纤维状的颗粒物,形成多边形,包裹着大小不等的、电子致密的圆球状或多边形物质(图4F).上皮细胞的分泌方式以局部分泌为主(图4F).

A.粗面内质网;B.分泌物;C.粗面内质网;D.细胞器;E.分泌物;F.腺管管腔周围.cj.细胞连接;er.粗面内质网;fm.纤维状物质;g.高尔基体;gr.颗粒物质;mi.微绒毛;mt.线粒体;n.细胞核;pv.多边形分泌物;sv.分泌泡.标尺= 1 μm.
2.3透明腺管发育的超微结构
2.3.1 初生期的超微结构
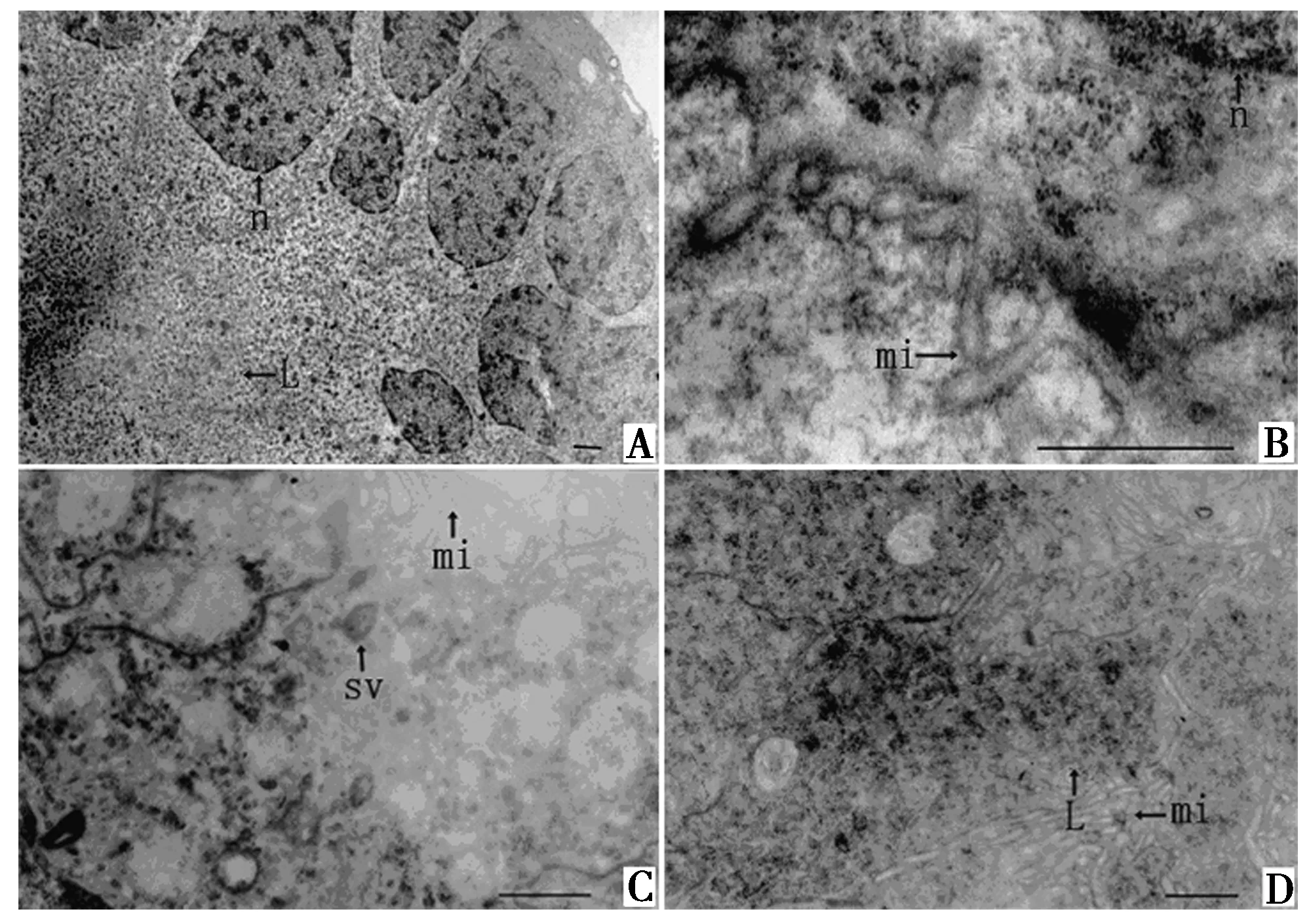
羽化之后,在乳白长腺管和乳白短腺管之间出现透明腺管.羽化1~9 d透明腺管管壁上皮细胞排列不规则,未形成单层.腺管管腔较小.管壁上皮细胞结构与前2类腺管相似,管壁结构从内到外依次为:上皮细胞层、基膜、肌肉层、围膜.上皮细胞靠近管腔的顶部含有一些分泌泡(图5A),细胞之间连接紧密(图5B).管腔中未见分泌物.
2.3.2 少生期的超微结构
羽化10 d后的透明腺管管壁上皮细胞为单层,相对前一时期管腔较大.上皮细胞靠近基膜的细胞质中含有大量电子透明小泡(图5C),上皮细胞靠近管腔的顶部含有较多分泌泡,管腔中含有少量电子致密的颗粒状分泌物(图5D).
2.3.3 成熟期的超微结构
性成熟后透明腺管的管壁上皮细胞扁平,单层排列.上皮细胞基部含有大量粗面内质网,内质网之间存在片层体结构(图5E).与前一时期相似,上皮细胞富含电子透明的小泡.腺管管腔中,接近管腔中心的分泌物为绒球状的物质和松散的细颗粒物质;靠近上皮细胞的分泌物为电子致密的颗粒物和纤维状物质(图5F).上皮细胞的分泌方式主要为顶质分泌(图5F).

A.分泌泡;B.细胞连接;C.电子透明小泡;D.管腔中的分泌物;E.粗面内质网;F.细胞顶膜部微绒毛和分泌物.b.片层体;bl.基膜;cj.细胞连接;er.粗面内质网;fm.纤维状物质;gr.颗粒物质;mi.微绒毛;mt.线粒体;n.细胞核;sv.分泌泡;v.电子透明小泡.标尺= 1 μm.
3 讨论
Kaulenas等认为昆虫雄性附腺腺体一般成形于末龄若虫期或蛹期,羽化数天后会迅速增长[6].黄粉虫Tenebriomolitor雄性附腺在蛹期已形成腺管,分泌上皮细胞中含有少量的内质网及分散的线粒体,羽化后附腺分泌细胞中内质网和高尔基体数量迅速增加[9].家蟋蟀Achetadomesticus在羽化之后,雄性附腺的上皮细胞中逐渐聚集大量粗面内质网,2~3周腺体发育成熟[10].
优雅蝈螽成虫雄性附腺腺管数量众多,有数百根,至少可以分为3类,即乳白长腺管、乳白短腺管和透明腺管[8].该虫与黄粉虫类似,雄性附腺的乳白长腺管和乳白短腺管在末龄若虫期已经发育为管状,但其管壁上皮细胞未形成单层,细胞之间连接不紧密,靠近管腔的细胞排列相对规则,上皮细胞内仅有一些核糖体,管腔小,管腔中也没有分泌物.透明腺管与乳白长腺管、乳白短腺管不同,其形成较晚,到羽化后才出现.
优雅蝈螽雄性成虫羽化后的少生期3类腺管的基本结构趋向一致,与成熟期的腺管结构相似,上皮细胞逐渐形成单层,腺管管腔开始出现少量分泌物.
优雅蝈螽雄性成虫性成熟期的3类腺管上皮细胞均为单层,排列紧密,细胞内有大量核糖体、粗面内质网、线粒体、高尔基体等细胞器,管腔中聚集了大量分泌物.上皮细胞富含粗面内质网、线粒体、高尔基体,具有分泌细胞的功能,与管腔中出现大量分泌物相一致.成熟期乳白长腺管管壁上皮细胞存在细胞凋亡的现象(图2F).细胞凋亡是一个受到一系列相关基因严格调控的细胞死亡过程,细胞分裂产生新的细胞,替换那些衰老的、失去功能的细胞[11-12].笔者推测,性成熟期乳白长腺管管壁上皮细胞出现细胞凋亡的现象可能是上皮细胞的正常更替,即使附腺发育成熟后仍然存在细胞分裂产生新的细胞.该虫雄性附腺在发育过程中,上皮细胞中的细胞器在种类和数量上表现出由分散到集中和由少到多的趋势,管腔中分泌物从无到有,从少到多,最后管腔中充满分泌物.
尽管优雅蝈螽雄性成虫附腺腺管数量众多,种类较多,但其发育过程基本遵循一般昆虫雄性附腺的发育规律.其雄性附腺的发育分化始于末龄若虫期,羽化后附腺上皮细胞快速分裂分化,细胞内的细胞器迅速增加,管腔中累积大量分泌物.
[1]GILLOTT C.Male accessory gland secretions: modulators of female reproductive physiology and behavior[J].Ann Rev Entomol, 2003, 48: 163-184.
[2]WOLFNER M F.The gifts that keep on giving: physiological functions and evolutionary dynamics of male seminal proteins inDrosophila[J].Heredity, 2002, 88: 85-93.
[3]CHEN P S.The functional morphology and biochemistry of insect male accessory glands and their secretions[J].Ann Rev Entomol, 1984, 29: 233-255.
[4]SWANSON W J.Sex peptide and the sperm effect inDrosophilamelanogaster[J].Pnas, 2003, 100(17): 9643-9644.
[5]COUCHE G A, GILLOTT C.Structure of the accessory reproductive glands of the male migratory grasshopper,Melanoplussanguinipes[J].Morphol, 1990, 203: 219-245.
[6]KAULENAS M S.Insect accessory reproductive structures: function, structure and development[M].New York : Sprinkler-Verlag, 1992:1-159.
[7]ROGINA B.The effect of sex peptide and calorie intake on fecundity in femaleDrosophilamelanogaster[J].Scientific World Journal, 2009, 9: 1178-1189.
[8]冯晓丽, 常岩林, 王莉,等.优雅蝈螽雄性附腺结构与分泌蛋白特性[J].昆虫知识, 2009, 46(5): 761-764.
FENG Xiaoli, CHANG Yanlin, WANG Li, et al.Structure and traits of secreted proteins of male accessory glands ofGampsocleisgratiosa[J].Chinese Bulletin of Entomology, 2009, 46(5): 761-764.
[9]HAPP G M.Structure and development of male accessory glands in insects[C]//KING R C, AKAI H.Insect Ultrastructure.New York: Plenum Press, 1984, 2: 365-396.
[10]KAULENAS M S, POTSWALD H E, BURNS A L ,et al.Development of structural and functional specializations for export protein synthesis by the accessory gland of the male cricket,AchetadomesticusL.(Orthoptera: Gryllidae)[J].Int J Insect Morphol & Embryol, 1979, 8(1): 33-49.
[11]BIALIK S, KIMCHI A.The death-associated protein kinases: structure, function and beyond[J].Annu Rev Biochem, 2006, 75: 189-210.
[12]EISENBERG-LERNER A, BIALIK S, SIMON H-U,et al.Life and death partners: apoptosis, autophagy and the cross-talk between them[J].Cell Death Diff, 2009, 16(7):966-975.
UltrastructureofmaleaccessoryglanddevelopmentfromGampsocleisgratiosa
ZHANGXiaoxia,CHANGYanlin,SHIFuming,ZHOUNa
(College of Life Sciences, Hebei University, Baoding 071002, China)
The ultrastructure of diffrent developmental periods fromGampsocleisgratiosaBrunner von Wattenwyl were examined by TEM (transmission electron microscope).The results revealed that long opalescent gland and short opalescent gland were fully differentiated in last-instar nymph but hyaline gland after eclosion.In epithelial cells, organelles under went the change from dispersed to concentrative and few to more during the male accessory glands development.Secretions under went the change from few to more and were filled in whole gland lumen in mature period.
Orthoptera;Gampsocleisgratiosa; male accessory glands; development
10.3969/j.issn.1000-1565.2013.01.012
2012-02-10
国家自然科学基金资助项目(31093430;31071955)
张小霞(1985-),女,内蒙古鄂尔多斯人,河北大学硕士研究生.E-mail:shuqi1229@126.com
常岩林(1965-),女,山西寿阳人,河北大学副教授,主要从事昆虫生理研究.E-mail:changylin@126.com
O96
A
1000-1565(2013)01-0057-06
(责任编辑赵藏赏)
猜你喜欢
浙江医学(2023年2期)2023-03-01 09:19:16
粮油食品科技(2020年6期)2020-12-08 03:26:50
化工管理(2020年14期)2020-06-15 06:58:54
老友(2019年8期)2019-09-09 16:02:10
川北医学院学报(2018年4期)2018-09-04 12:33:52
保健与生活(2018年15期)2018-01-27 15:14:20
保健与生活(2018年1期)2018-01-27 01:25:15
——鞭毛、纤毛和微绒毛(2)
生物学通报(2017年8期)2017-04-09 00:54:29
北方文学(2016年2期)2016-03-22 08:39:18
长春理工大学学报(自然科学版)(2015年5期)2015-10-12 02:18:26
