1株蛇足石杉根际铁载体细菌的分离、筛选与鉴定
2013-10-25 08:40:34肖艳红刘祝祥
微生物学杂志 2013年2期
肖艳红,李 菁,刘祝祥,杜 次,李 贵
(吉首大学植物资源保护与利用湖南省高校重点实验室,湖南 吉首 416000)
蛇足石杉(Huperzia serrata(Thumbs.)Trev.)为我国传统中草药,属石杉科(Huperiaceae)石杉属(Huperzia Bernh.)的蕨类植物。民间常用于治疗痈疖肿毒、跌打损伤等症。从蛇足石杉中分离得到的石杉碱甲(Huperzine A),是一种选择性强、高效、可逆的乙酰胆碱酯酶抑制剂,对于预防和治疗老年性痴呆具有显著疗效且毒副作用小[1]。然而,石杉类植物作为古老的蕨类植物类群,存在着生境特化、植株矮小、居群偏小、自然更新速度慢和生长周期较长等诸多不利因素。通过其他途径获得石杉碱无法实现工业化生产规模。目前,石杉碱甲的临床用药都直接或间接地源于蛇足石杉的自然资源,这导致了蛇足石杉野生资源破坏严重而日趋枯竭。铁载体(siderophores)是一类具有很强的特异螯合Fe3+(螯合系数可达1020~1030)的小分子(1 ~2 ku)化合物[2],它的产生是地球上微生物适应地球地壳高氧低铁环境的结果[3]。细菌利用合成的铁载体攫取外界游离Fe3+来维持正常的生理活动。同时,细菌在环境中的生存地位也可以通过其对环境中铁源的争夺能力而反映出来[4]。因此对自然条件下产铁载体菌的研究能够解决许多因细菌引起的植物病害问题。若能从土壤中分离、筛选出产铁载体的拮抗菌,再将其应用到田间,在作物根际对铁进行生物富集,从而提高土壤中铁的生物有效性,这将大大地促进作物的生长[4-5]。由于产生铁载体的细菌与不能产生铁载体的或产生铁载体少的有病害的微生物竞争铁元素,从而抑制有害微生物的生长繁殖,改善整个作物的生态环境,为保护作物免受病原菌的侵害起到一定的作用[4-6]。迄今,有关蛇足石杉铁载体细菌方面的研究尚未见报道。本文以蛇足石杉根际土壤为研究对象,对蛇足石杉根际土壤微生物进行分离、筛选,获得1株产铁载体细菌,并对该菌进行了初步鉴定,为蛇足石杉根际促生菌的开发利用提供优良的菌种来源,同时为解决蛇足石杉快速繁殖及生长发育问题提供一定的参考。
1 材料与方法
1.1 材料
1.1.1 蛇足石杉根际土样采集与处理 供试材料蛇足石杉根际土样采自湖南省古丈县高望界林场(海拔 570 m,N28°38'27.7″,E110°02'02.1″),采用五点法取样。采样时选取健康、长势较为旺盛的植株,去除表层土壤和杂物。采集其根际土样时将蛇足石杉整个植株连根挖出,去除根系上附着的大土粒,用无菌的毛刷将根上粘着的细土扫落到无菌袋后封口,贴上标签,然后将土样用冰盒迅速带回实验室进行根际土壤细菌的分离。
1.1.2 培养基及铁载体检测液 ①细菌分离纯化培养基(牛肉膏蛋白胨培养基)[7]:牛肉膏 3.0 g,蛋白胨 5.0 g,琼脂 18 g,NaCl 5.0 g,蒸馏水1000 mL,pH 7.0 ~7.2;②铁载体检测 CAS 染液:CAS(铬天青)1 mmol/L,FeCl30.1 mmol/L,十六烷基三甲基溴化铵(HDTMA)[8]4 mol/L;0.1 mol/L磷酸盐缓冲液(pH 6.8):每100 mL含Na2HPO4· 12H2O 2.427 g,NaH2PO4·2H2O 0.5905 g;KH2PO40.075 g,NH4Cl 0.250 g,Na-ClO 0.125 g,使用时稀释 10 倍[9];③铁载体检测CAS检测平板配制:每100 mL含20%蔗糖溶液1 mL,10% 酸水解酪素 3 mL,1 mmol/L CaCl2100 μL,1 mmol/L MgSO42 mL,琼脂 1.8 g[9]。
1.1.3 仪器、设备及试剂 生化培养箱BMJ100(上海博讯实业有限公司),高压蒸汽灭菌锅MJ-54A(美国STIK),单人水平垂直两用净化工作台CJ-1F(苏州华宏净化技术有限公司),高速台式离心机 Centrifuge 5810R(德国 Eppendorf),PCR仪ABI 2720型(美国ABI),凝胶成像系统Tanon-2500(上海天能科技有限公司),所用分子生物学试剂均购自上海生物工程有限公司。
1.2 方法
1.2.1 蛇足石杉根际细菌分离与纯化 称取1 g根际土样加入到装有99 mL无菌生理盐水的250 mL三角瓶中,120 r/min振荡30 min,得到菌悬液,梯度稀释至10-7g/mL。采用涂布平板法,取0.2 mL稀释后不同浓度梯度的菌悬液分别涂布到分离培养基上,每个稀释梯度设3次重复,将涂布好的平板至37℃培养箱培养,24 h后开始观察,同时记录分离平板上菌株的菌落特征,根据菌落形态、颜色和大小等特征挑取不同的单菌落,在分离纯化培养基上用四分体划线法经连续4代划线纯化,得到细菌纯培养物。
1.2.2 蛇足石杉根际细菌产铁载体细菌的初筛
将细菌纯培养物接种到CAS检测平板上,37℃培养48 h,经过培养的CAS检测平板上,分泌铁载体的细菌菌落周围会出现明显的橙黄色晕圈,并可通过晕圈直径大小来初步判断菌株产铁载体能力的强弱。
1.2.3 蛇足石杉根际细菌产铁载体细菌的复筛
挑取晕圈比较明显的菌落,接种于液体牛肉膏蛋白胨培养基中发酵培养,37℃,150 r/min振荡培养48 h,然后将菌悬液8000 r/min离心15 min后,得到细菌发酵上清液。分别取发酵上清液与CAS检测液各3 mL,充分混匀。1 h后采用721型分光光度计在630 nm波长处测定吸光值(As),并取双蒸水作对照调零。另取3 mL CAS检测液与3 mL未接种的牛肉膏蛋白胨液体培养基上清液充分混匀,同上测定吸光值即为参比值(Ar)。按照文献[10]对铁载体活性单位的定义,根据公式计算菌株产铁载体的量:
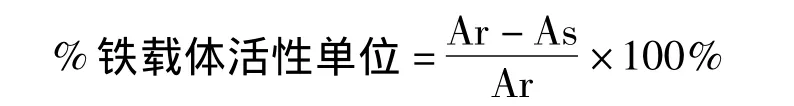
1.2.4 细菌菌落形态、革兰染色与显微形态观察
挑取纯化后的单菌落,四分体划线接种到牛肉膏蛋白胨平板上,37℃培养24 h,进行菌落形态观察并拍照,然后挑取单菌落进行革兰染色和显微形态观察。
1.2.5 细菌基因组DNA提取和16S rDNA基因的扩增 酶解法提取菌体中的基因组DNA:取50 mg菌体加1×TE稀释成2 mg/mL的溶菌酶500 μL放入37℃摇床1~2 h,加入50 μL 20%SDS和5 μL浓度为20 μg/mL蛋白酶 K混匀1 min,置55℃烘箱保温60 min。结束后加入550 μL苯酚∶氯仿∶异戊醇(25∶24∶1),混匀抽提,12000 r/min离心10 min,取上清液。上清液加入800 μL 无水乙醇、80 μL 3mol/L 乙酸钠(pH 4.8~5.2)室温放置 15 min。12000 r/min 离心10 min,除上清液,加入200 μL 70%乙醇洗盐1~2次,12000 r/min低温离心5 min,弃乙醇。55℃干燥后加入1×TE 50 μL溶解DNA,-20℃保存备用。采用细菌16S rDNA PCR扩增的通用引物(正向引物:5'-CCGGATCCAGAGTTTGATCCTGGTCAGAACGAACGC-3';反向引物:5'-CGGGATCCTACGGCTACCTTGTTACGACTTCACCCC-3'),以细菌基因组DNA为模板进行PCR扩增。PCR反应程序:94℃预变性5 min;30个循环:94℃变性30 s,54℃退火30 s,72℃延伸90 s;72℃延伸10 min,4℃保存。1%琼脂糖凝胶电泳检测PCR扩增产物,将PCR产物送上海生工生物工程有限公司测序。
1.2.6 16S rDNA基因系统发育分析 将 PCR扩增产物进行部分测序,测序结果利用BLAST软件(http//www.ncbi.nlm.nih.gov/BLAST/)在GenBank、EMBL及DDBJ等数据库中进行相似性搜索,选取同源性最高的且有效发表的菌株序列作为参比对象,用CLUSTAL W软件进行多序列比对,计算测试菌株和参比菌株之间的序列相似性,采用 Neighbor-Joining法,利用 MEGA5软件(http://www.Megasoftware.net)构建系统发育树[11],自展数(bootstrap)为 1000。
2 结果与分析
2.1 蛇足石杉根际铁载体细菌的分离与筛选
用牛肉膏蛋白胨培养基分离蛇足石杉根际细菌,依据菌落形态、颜色、大小及革兰染色结果,挑选100株不同细菌。将100株细菌培养于CAS检测平板上,经过24 h培养后,得到1株产铁载体能力较强的菌,命名为JSX 389。其周围出现橙黄色的晕圈(图1C),表明该菌株具有产铁载体的能力。为了鉴定该菌株产铁载体能力,按1.2.3中的方法对该菌产铁载体能力进行定量检测;在波长630 nm 处As的光吸收值为0.167,而 Ar为0.341,可算得 As/Ar=0.490,(Ar-As)/Ar=51.02%,说明该菌株具有较强的产铁载体能力[8,12]。
2.2 细菌菌落形态、革兰染色与显微形态观察结果
按照2.1方法对菌株 JXS 389进行菌落形态、革兰染色与显微形态观察,结果如图1A、B所示。从图1中可以看出菌株JXS 389菌落为乳白色、湿润光滑微向上突起菌落;革兰染色后,显微观察菌体呈紫色,细杆状,表明该菌为革兰阳性杆菌。

图1 产铁载体菌形态特征及CAS检测筛选Fig.1 The morphology and screening of siderophore producing property by CAS detecting plates of strain JX S389
2.3 JSX 389的16S rDNA基因系统发育分析
对可产铁载体细菌JSX 389的16S rDNA全基因进行测序,得到总长为1515 bp的16S rDNA序列,并将测序结果提交GenBank,序列接收号为KC192729。根据16S rDNA基因序列构建的JSX 389的系统发育树见图2。从JSX 389系统发育树可以看出,菌株JSX 389与Lysinibacillus属菌株聚类在一个大枝上,JSX 389应属于Lysinibacillus属;JSX 389和有效发表菌株Lysinibacillus sinduriensis BLB-1 KCTC 13296T和Lysinibacillus massiliensis 4400831T以较高的自展值(99)聚在一个小枝上,且JSX 389与Lysinibacillus sinduriensis BLB-1 KCTC 13296T的序列相似性为98%,与Lysinibacillus massiliensis 4400831T的序列相似性为96.5%,按照16S rDNA序列相似程度小于97%可归于不同物种标准[13-14],菌株JSX 389可能与Lysinibacillus sinduriensis BLB-1 KCTC 13296T属同一物种。但是 Lysinibacillus sinduriensis BLB-1 KCTC 13296T分离自海洋,而JSX 389分离自蛇足石杉根际土壤,且能产生铁载体,具体分类地位还有待进一步研究。

图2 菌株JSX 389的系统进化树Fig.2 The phylogenetic tree of strain JSX 389
3 结论与讨论
本研究从蛇足石杉根际土样中分离了1株具产铁载体能力的细菌JSX 389,并对其菌落形态特征、革兰染色和显微形态特征进行观察,显示JSX 389为革兰阳性的杆菌。JSX 389的16S rDNA基因系统发育分析表明:JSX 389应为Lysinibacillus属菌株,因JSX 389和有效发表菌株Lysinibacillus massiliensis 4400831T聚在一个小枝上,且JSX 389与Lysinibacillus sinduriensis BLB-1 KCTC 13296T的序列相似性为98%,JSX 389可能与Lysinibacillus sinduriensis BLB-1 KCTC 13296T属同一物种,但后者分离自海洋,JSX 389分离自蛇足石杉根际土壤,且能产生铁载体,具体分类地位还有待研究。产铁载体细菌在环境中能够迅速稳定地分泌铁载体,这不仅可以实现可利用铁源的自足,还能为植物根际周围小生境提供过量的铁载体,为其他利用同源铁载体植物的生长提供便利,同时它也可与植物病原微生物争夺有限的铁营养,从而抑制病原微生物的生长[15]。JSX 389为蛇足石杉根际细菌,产生铁载体的能力较强,能否像其他植物根际铁载体细菌具有促生长作用,促进蛇足石杉的生长,还有待进一步深入研究。
[1]鲁翠涛,梅兴国,钟凡.千层塔生物学特性的初步研究[J].中国野生植物资源,2002,21(4):33-35.
[2]Shelly MP.Detection,isolation and characterization of siderophores[J].Methods in Enzymology,1994,235:329-344.
[3]David AH,Amy EW,Alison B,et al.Competition among marine phytoplankton for deferent chelated iron species[J].Letters to Nature,1999,400(6747):858-861.
[4]于柏峰,张慧云,刘冰,等.铜绿假单胞菌致病力和致病机理研究进展[J].微生物学杂志,2004,24(1):52-53.
[5]刘健,李俊,葛诚.微生物肥料作用机理的研究新进展[J].微生物学杂志,2001,21(1):33 -36,46.
[6]余贤美,郑服丛.嗜铁素在促进植物生长及病害防治等方面的应用[J].中国农学通报,2007,23(8):507-510.
[7]沈萍,范秀容,李广武.微生物学实验[M].北京:高等教育出版社,1999:69.
[8]王平,董飚,李阜棣,等.小麦根圈细菌铁载体的检测[J].微生物学通报,1994,21(6):323-326.
[9]赵翔,谢志翔,陈绍兴,等.适合高铁载体细菌筛选、检测体系的改进与探析[J].微生物学通报,2006,33(6):13-18.
[10]Machuca A,Milagres AMF.Use of CAS-Agar plate modified to study the effect of different variables on the siderophore production by Aspergillus[J].Letters in Applied Microbiology,2003,36(3):177-181.
[11]Tamura K,Peterson D,Peterson N,et al.MEGA5:Molecular Evolutionary Genetics Analysis using Maximum Likelihood,Evolutionary Distance,and Maximum Parsimony Methods[J].Molecular Biology and Evolution,2011,28(10):2731-2739.
[12]孙磊,邵红,刘琳,等.可产生铁载体的春兰根内生细菌多样性[J].微生物学报,2011,51(2):189-195.
[13]肖怀东,陈义光,刘祝祥,等.湛江硇洲岛海葵相关可培养细菌系统发育多样性研究[J].微生物学报,2009,49(2):246-250.
[14]Stackebrandt E,Goebel BM.Taxonomic note:a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology[J].International Journal of Systematic Bacteriology,1994,44(4):846-849.
[15]赵翔,陈绍兴,谢志雄,等.高产铁载体荧光假单胞菌Pseudomonas fluorescens sp-f的筛选鉴定及其铁载体特性研究[J].微生物学报,2006,46(5):691-695.
猜你喜欢
疯狂英语·初中天地(2022年1期)2022-07-07 08:37:54
环境保护与循环经济(2021年7期)2021-11-02 08:10:42
湖北农机化(2020年4期)2020-07-24 09:07:26
林业科技情报(2020年3期)2020-03-02 13:21:36
生物工程学报(2019年11期)2019-12-09 13:09:12
西南农业学报(2019年6期)2019-07-18 05:32:16
中成药(2018年5期)2018-06-06 03:12:19
中成药(2018年3期)2018-05-07 13:34:27
中国当代医药(2015年36期)2015-03-11 20:03:28
中国民族民间医药·上半月(2014年2期)2014-11-27 21:13:55