
多重耐药菌中β-内酰胺酶的基因型研究*
2013-10-11 05:50陈定强彭玉婷罗毓婷徐韫健林勇平广州医学院第一附属医院检验科510120
检验医学与临床 2013年1期
陈定强,彭玉婷,罗毓婷,徐韫健,林勇平,杨 羚(广州医学院第一附属医院检验科 510120)
细菌产生β-内酰胺酶是革兰阴性杆菌耐β-内酰胺类抗菌药物最普遍的机制,β-内酰胺酶相关基因赋予细菌编码对应酶的能力[1]。由于β-内酰胺类抗菌药物在临床上的广泛使用,产β-内酰胺酶的菌株也变得越来越常见,并在选择和进化压力中产生多种不同的酶基因型[2]。这些基因还往往与其他耐药相关基因(如喹诺酮类耐药基因和磺胺类耐药基因等)共同存在,导致细菌产生多重耐药性[3]。本研究收集59株临床分离的多重耐药菌,通过多重聚合酶链反应(PCR)法检测其携带的β-内酰胺酶基因类型,分析β-内酰胺酶基因在多重耐药菌中的分布特征,为进一步研究和控制细菌耐药性提供实验依据。
1 材料与方法
1.1 菌株来源 研究菌株为2009~2010年广州医学院第一附属医院临床分离的59株多重耐药菌,包括29株大肠杆菌和30株肺炎克雷伯菌。其中分离自尿液标本的32株,痰液标本19株,血液标本7株,脓液标本1株。细菌鉴定与药敏试验的质控菌株为大肠杆菌ATCC25922和金黄色葡萄球菌ATCC25923。
1.2 仪器与试剂 VITEK2全自动微生物分析仪、细菌鉴定卡与药敏卡为BioMerieux公司产品,PCR仪为Eppendorf公司产品,核酸电泳仪与凝胶成像仪为BioRad公司产品。MH培养基和药敏纸片由Oxoid公司提供。PCR扩增反应混合体系为Takara公司产品,DNA分子标记物购自Tiangen公司,琼脂糖购自Biowest公司。
1.3 细菌总DNA的提取 采用煮沸法提取细菌总DNA。挑取平板上的一个单菌落,重悬于300μL双蒸水中。置沸水浴中孵育10min后,13 500r/min离心5min,吸取上清液至新的微量离心管中,作为PCR反应的DNA模板。
1.4 PCR引物 PCR引物序列见表1,所有引物由上海英潍捷基公司合成。
1.5 β-内酰胺酶基因的检测 按照文献[4]报道的方法,用单重和多重PCR法对21种β-内酰胺酶基因进行检测。PCR反应在20μL体系中进行,包括2×PCR扩增反应混合体系10 μL,细菌总DNA模板1μL,PCR引物各1μL,并用双蒸水补至20μL。PCR反应的条件为:95℃预变性2min,然后95℃变性30s,退火温度见表1,时间为30s,72℃ 延伸1min,进行35个循环,最后延伸7min。PCR产物在2%的琼脂糖凝胶中电泳40min,溴化乙锭溶液染色后用凝胶成像系统观察结果。
2 结 果
2.1 59株多重耐药菌的药敏结果 59株多重耐药菌包括29株大肠埃希菌与30株肺炎克雷伯菌,对氨苄西林的耐药率为100%,对头孢他啶的耐药率为91.5%,对庆大霉素的耐药率为66.1%,对环丙沙星的耐药率为72.9%,值得注意的是3.4%及1.7%的菌株分别对厄他培南和亚胺培南耐药,显示耐碳青霉烯类药物菌株的出现。对其他抗菌药物的耐药情况见表2。
2.2 β-内酰胺酶基因型分布 在21种β-内酰胺酶基因型中,有10种在多重耐药菌株中被检出(表3)。阳性率最高的是TEM,为54.2%(32/59);SHV 的阳性率为45.8%(27/59);CTX-M-G1、G2和G9的检出率分别为32.2%(19/59)、23.7%(14/59)和52.5%(31/59),说明CTX-M 型β-内酰胺酶分布非常广泛;另外,分别有4株和2株肺炎克雷伯菌检出DHA和MOX基因,CMY、OXA-1和ACT基因则各有1株检出。未检到含GES、VEB、IMP、VIM、KPC、CTX-M-G8/25、ACC、FOX、PER、OXA-48等基因的菌株。
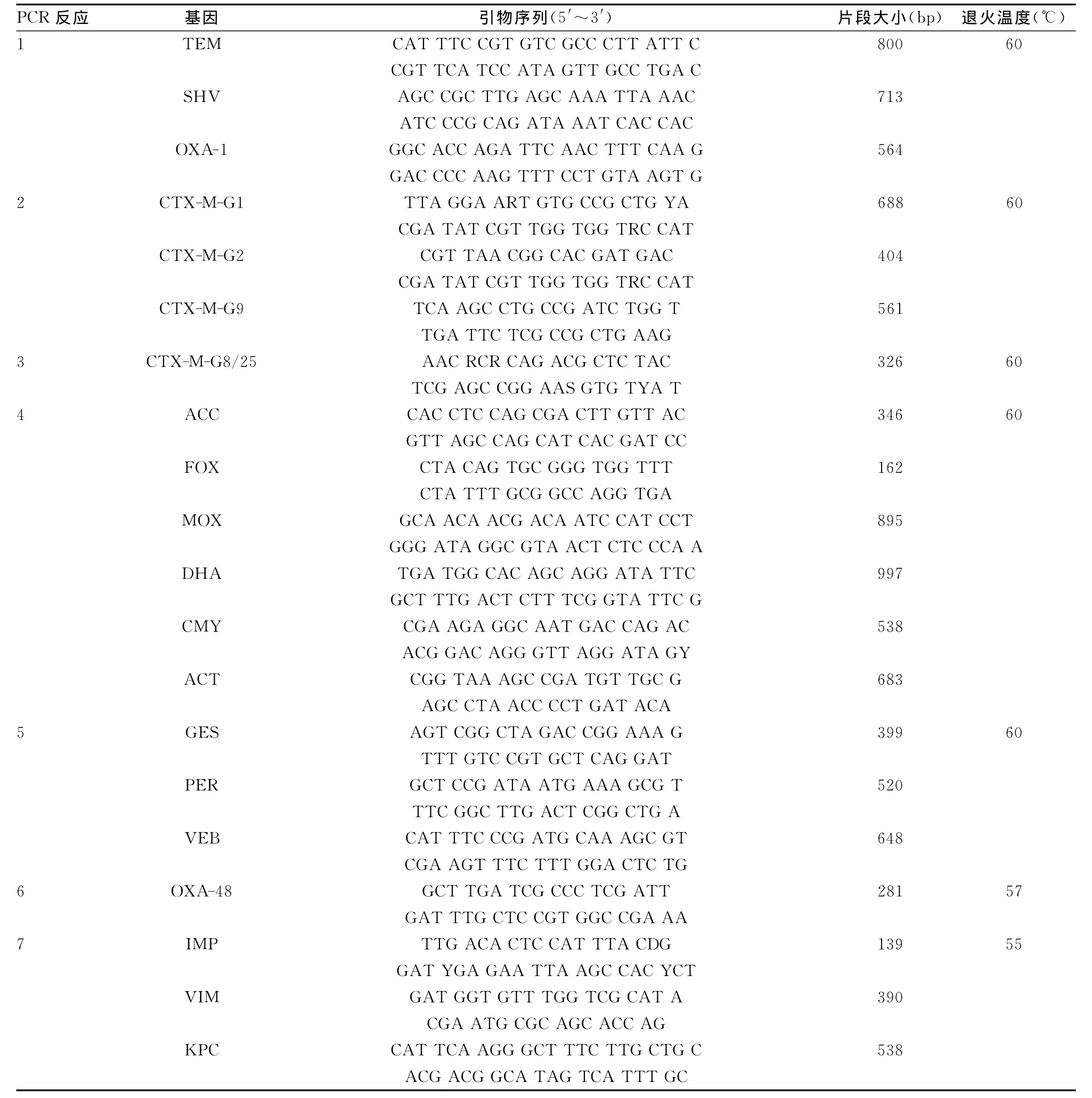
表1 PCR引物序列和反应条件
2.3 多种β-内酰胺酶基因型共存情况分析 见表4。59株多重耐药菌中有67.8%(40/59)的菌株携带两种以上的β-内酰胺酶基因型,其中肺炎克雷伯菌携带两种以上基因型菌株更为常见,达76.7%(23/30),值得注意的是,有2株肺炎克雷伯菌携带5种β-内酰胺酶基因型。
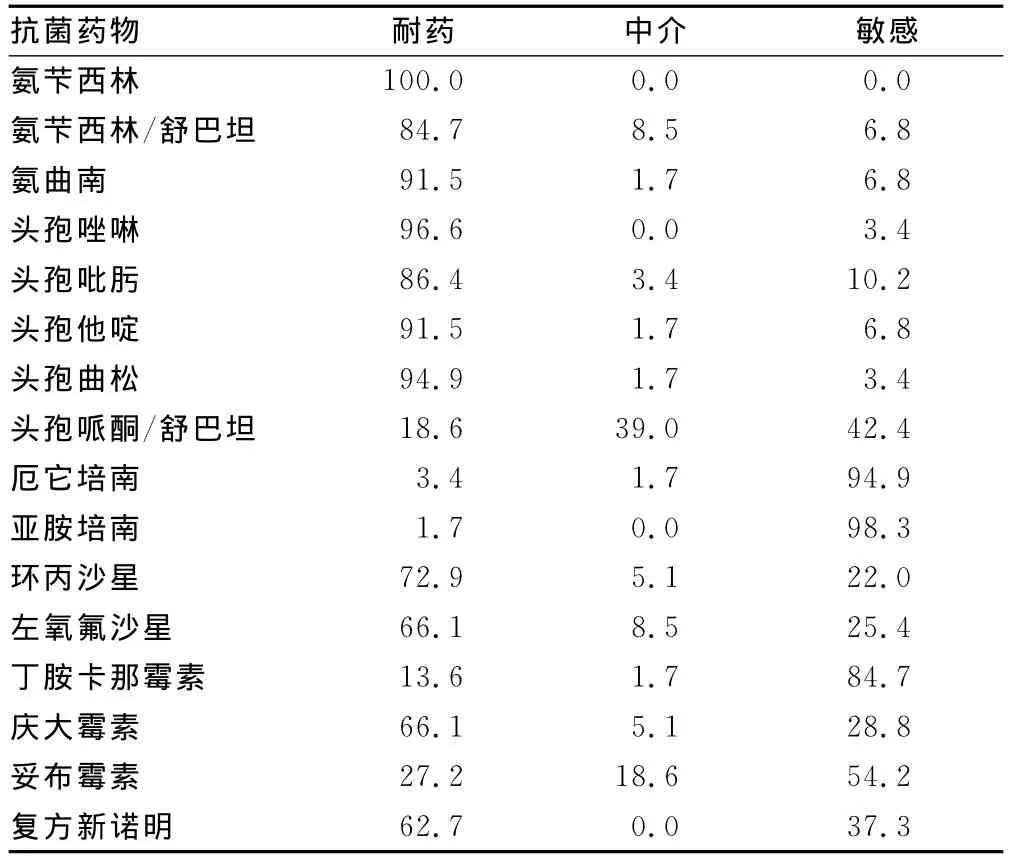
表2 59株多重耐药菌株的药敏结果(%)

表3 59株多重耐药菌中β-内酰胺酶基因型分布情况(n)
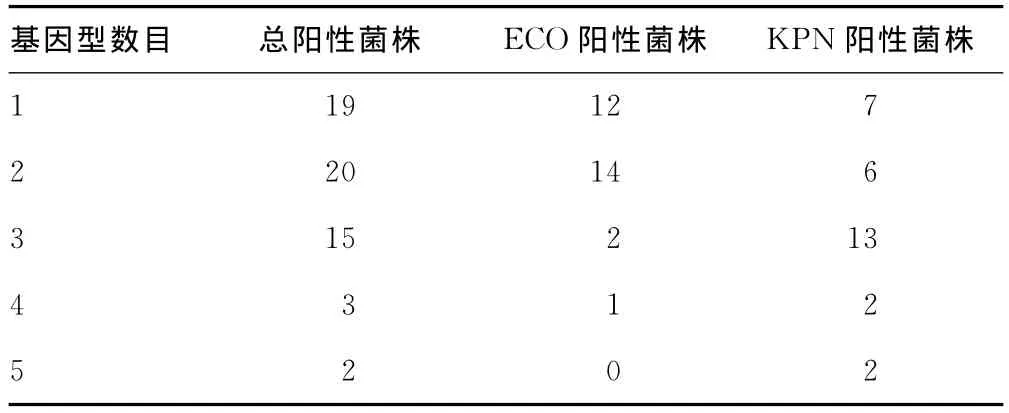
表4 多种β-内酰胺酶基因型共存的菌株数目(n)
3 讨 论
大肠杆菌和肺炎克雷伯菌是临床感染常见的革兰阴性菌,近年来由于抗菌药物的广泛应用,多重耐药菌株的出现越来越普遍[5]。β-内酰胺类抗菌药物是临床治疗细菌感染的最常用抗菌药物,但β-内酰胺酶的产生和传播令细菌对这类抗菌药物的耐药性不断增强[6]。本研究对59株多重耐药的大肠杆菌和肺炎克雷伯菌的药敏试验结果分析显示,这些菌株对多种青霉素类和头孢菌素类药物的耐药性非常严重,而且出现了耐碳青霉烯类药物的菌株。有文献报道,产β-内酰胺酶的细菌往往可同时携带对其他种类抗菌药物耐药的基因,这些菌株易发展成多重耐药菌,给临床感染治疗带来很大的困难[7]。本研究中收集的59株多重耐药菌对喹诺酮类和磺胺类药物的耐药率均较高,也证实了这一点。
对21种β-内酰胺酶基因的检测结果显示,本研究收集的多重耐药菌株携带其中10种。对大肠杆菌和肺炎克雷伯菌的检出率分析显示,大肠杆菌仅检出4种β-内酰胺酶基因,而肺炎克雷伯菌则检出10种,由此显示后者的β-内酰胺酶基因型更具多样性。多重耐药的肺炎克雷伯菌可携带多种耐药基因,且往往是定位在可转移的质粒上,可通过在肠杆菌科细菌间的接合作用引起这些基因的扩散,进一步加大了预防控制的难度[8]。虽然研究菌株中包含了耐碳青霉烯类药物的菌株,但并未检出VIM、IMP和KPC等碳青霉烯酶,说明这些菌株中碳青霉烯类药物耐药性可能与其他因素有关,需要进一步的研究以阐明其耐药机制。
本研究还发现多重耐药菌中同时携带多种β-内酰胺酶基因的情况十分普遍。不同的β-内酰胺酶有不同的水解底物,对仅产一种β-内酰胺酶基因的菌株来说,可以选择其不能水解的β-内酰胺类抗菌药物进行治疗。但多种基因型共存于同一菌株时,细菌往往对所有青霉素类和头孢菌素类药物都耐药,大大增加了治疗的难度。如何控制和治疗这些菌株引起的感染,已经成为临床抗菌治疗和医院感染控制迫切需要解决的大问题。
[1] Bush K.Alarming beta-lactamase-mediated resistance in multidrug-resistant Enterobacteriaceae[J].Curr Opin Microbiol,2010,13(5):558-564.
[2] 谭文彬.细菌耐药的基因机制研究进展[J].中国病原生物学杂志,2009,4(7):543-544.
[3] Nikaido H.Multidrug resistance in bacteria[J].Annu Rev Biochem,2009,78:119-146.
[4] Dallenne C,Da Costa A,Decre D,et al.Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae[J].J Antimicrob Chemother,2010,65(3):490-495.
[5] Bonomo RA,Szabo D.Mechanisms of multidrug resistance in Acinetobacter species and Pseudomonas aeruginosa[J].Clin Infect Dis,2006,43(Suppl 2):49-56.
[6] 刘云红.临床常见耐药性细菌的耐药机制[J].中国现代医药杂志,2009,11(6):129-130.
[7] Ogbolu DO,Daini OA,Ogunledun A,et al.High levels of multidrug resistance in clinical isolates of Gram-negative pathogens from Nigeria[J].Int J Antimicrob Agents,2011,37(1):62-66.
[8] Woodford N,Turton JF,Livermore DM.Multiresistant Gram-negative bacteria:the role of high-risk clones in the dissemination of antibiotic resistance[J].FEMS Microbiol Rev,2011,35(5):736-755.
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23
现代临床医学(2022年3期)2022-06-06
当代医药论丛(2022年4期)2022-03-20
昆明医科大学学报(2021年8期)2021-08-13
中国现代医药杂志(2020年10期)2020-12-14
天津医科大学学报(2019年3期)2019-08-13
中国乳品工业(2017年4期)2017-05-15
上海农业学报(2017年3期)2017-04-10
作物研究(2014年6期)2014-03-01
中国乳业(2013年8期)2013-08-31
