
环境高温与饲粮粗蛋白质水平对肉鸡生产性能、氮代谢和氮排放的影响
2013-09-20 03:05冯京海张敏红刘圈炜姜礼文
动物营养学报 2013年10期
陈 燕 冯京海 张敏红* 刘圈炜 姜礼文
(1.中国农业科学院北京畜牧兽医研究所,北京 100193;2.海南省农业科学院畜牧兽医研究所,海口 571100)
环境高温可引起肉鸡热应激,从而引起肉鸡采食量、体增重等生产性能显著下降[1-2]。而且,规模化、集约化畜禽养殖带来了大量氮排放的问题。据统计,全球畜牧业氮排放量为8.0×107~1.3 ×108t[3],排放的氮在微生物作用下,产生大量氨气,严重污染了生活环境。因此,改善环境高温下动物热应激,提高热应激动物氮利用率、降低氮排放是今后畜牧业面临的重大课题。
随着环境温度升高,热应激使肉鸡肠上皮细胞形态损伤,消化道消化液分泌减少[4]。畜禽消化道内蛋白质消化和氨基酸吸收发生改变[5],Dibner等[6]研究发现,热应激肉鸡的肠道上皮细胞对DL-蛋氨酸的吸收率只有总摄入量的34%。热应激使肉鸡蛋白质分解代谢和糖异生加强,蛋白质合成减弱[7];Geraert等[8]发现,32 ℃ 热应激组肉鸡体蛋白沉积率会显著低于22℃组。由此推测,高温可能导致蛋白质等营养物质消化率下降,排出增加。但有部分研究者得出与以上不同的研究结果,Wilson等[9]报道,高温仅使食糜在消化道内停留时间延长,消化道的蠕动减缓;Chwalibog等[10]发现,15~40℃的环境温度不影响21~42日龄肉鸡体蛋白含量。在肉鸡饲粮蛋白质供给和氮排放研究方面,Zervas 等[11]、Namroud 等[12]、Keshavarz 等[13]和 Lenis等[14]研究均表明,通过降低饲粮粗蛋白质(CP)水平同时补充合成必需氨基酸(EAA),会显著减少畜禽氮排放量。但迄今为止,未见有关高温时通过改变蛋白质供给模式减少肉鸡氮排泄的报道,也未见高温和饲粮蛋白质供给模式对肉鸡氮代谢和氮排放影响是否存在互作的报道。为此,本试验以肉鸡为研究对象,旨在研究高温环境与饲粮CP水平对其氮代谢的影响,从而为高温应激条件下肉鸡饲粮中蛋白质的供给及体内氮的沉积获得具有参考价值的数据,为缓解肉鸡热应激、减少氮排放提供试验依据。
1 材料与方法
1.1 试验饲粮与分组设计
本试验采用2因素3水平试验设计:因子1为饲养方式,设有23℃恒温自由采食组(23℃ ad libtum group,23℃ AL组)、23℃ 恒温配对组(23℃ pair-fed group,23℃ PF组,其饲粮按照28/32℃日变高温自由采食组采食量供给)、28/32℃日变高温自由采食组(28/32℃ ad libtum group,28/32℃ AL组,28℃—32℃—28℃日变循环高温);因子2为饲粮CP水平,设21.19%、18.18%、14.90%3个水平(实测值),通过补充人工合成EAA,使各组饲粮蛋氨酸、赖氨酸、苏氨酸、色氨酸水平满足肉鸡营养需要,且保持一致。试验分组设计见表1。

表1 试验分组设计Table 1 The design of experimental groups
参照 NRC(1994)爱拔益加(Arbor Acres,AA)鸡营养需要及《Arbor Acres肉鸡饲养手册》推荐水平,配制玉米-豆粕型饲粮,添加人工合成EAA,使试验饲粮除CP水平外,各组饲粮 EAA(包括蛋氨酸、赖氨酸、苏氨酸、色氨酸)水平满足肉鸡营养需要,且保持一致(表2)。
1.2 试验动物与饲养管理
选取1日龄健康AA肉鸡620只,公母各占1/2,1~21日龄饲养于同一鸡舍,饲喂相同饲粮,环境温湿度相同。至21日龄,选取体重接近的健康AA肉鸡576只,转入人工环境控制舱内饲养,分为9组,每个组设8个重复,每个重复8只。24 h光照,相对湿度范围为(55±5)%,自由采食和饮水,其他饲养管理与免疫程序按常规进行。21~27日龄预饲 7 d,28~42日龄为正试期(14 d)。
23℃ AL组为23℃恒温环境并自由采食;23℃ PF组为23℃恒温环境,试验期第1天自由采食,从第2天开始每天饲粮投喂量与前1天28/32℃ AL组肉鸡采食量相同;28/32℃ AL组为自由采食,28℃—32℃—28℃日变循环高温,模式为 07:00开始升温,09:00升至32℃,09:00—14:00维持在(32±1)℃ 5 h,14:00开始降温,16:00降至28℃,直至次日07:00,温度维持在(28±1)℃ 15 h。试验在中国农业科学院北京畜牧兽医研究所昌平试验基地动物营养国家重点实验室环控舱中进行。
1.3 样品采集与测定指标
1.3.1 生产性能指标
每日记录肉鸡健康状况、死亡数,于28、35和42日龄断料12 h后以重复为单位对肉鸡进行称量并记录体重(body weight,BW),同时统计各个重复的耗料量,计算试验期平均日增重(average daily gain,ADG)、平均日采食量(average daily feed intake,ADFI)、料 重 比 (feed to gain ratio,F/G)和死亡率(mortality rate,MR)。
1.3.2 生理指标
于肉鸡42日龄(12:00)时每重复随机选取2只(公母各1只),测量并记录各重复肉鸡的肛温(rectal temperature,RT)[用电子显示温度计插入肉鸡泄殖腔(4.0±0.5)cm内测肛门温度]和呼吸率(respiratory rate,RR)(使用秒表和计数器,选取安静状态下的肉鸡,观察肉鸡翅膀及肛门下部 羽毛的起伏动作,一起一伏为1次,记录1 min)。
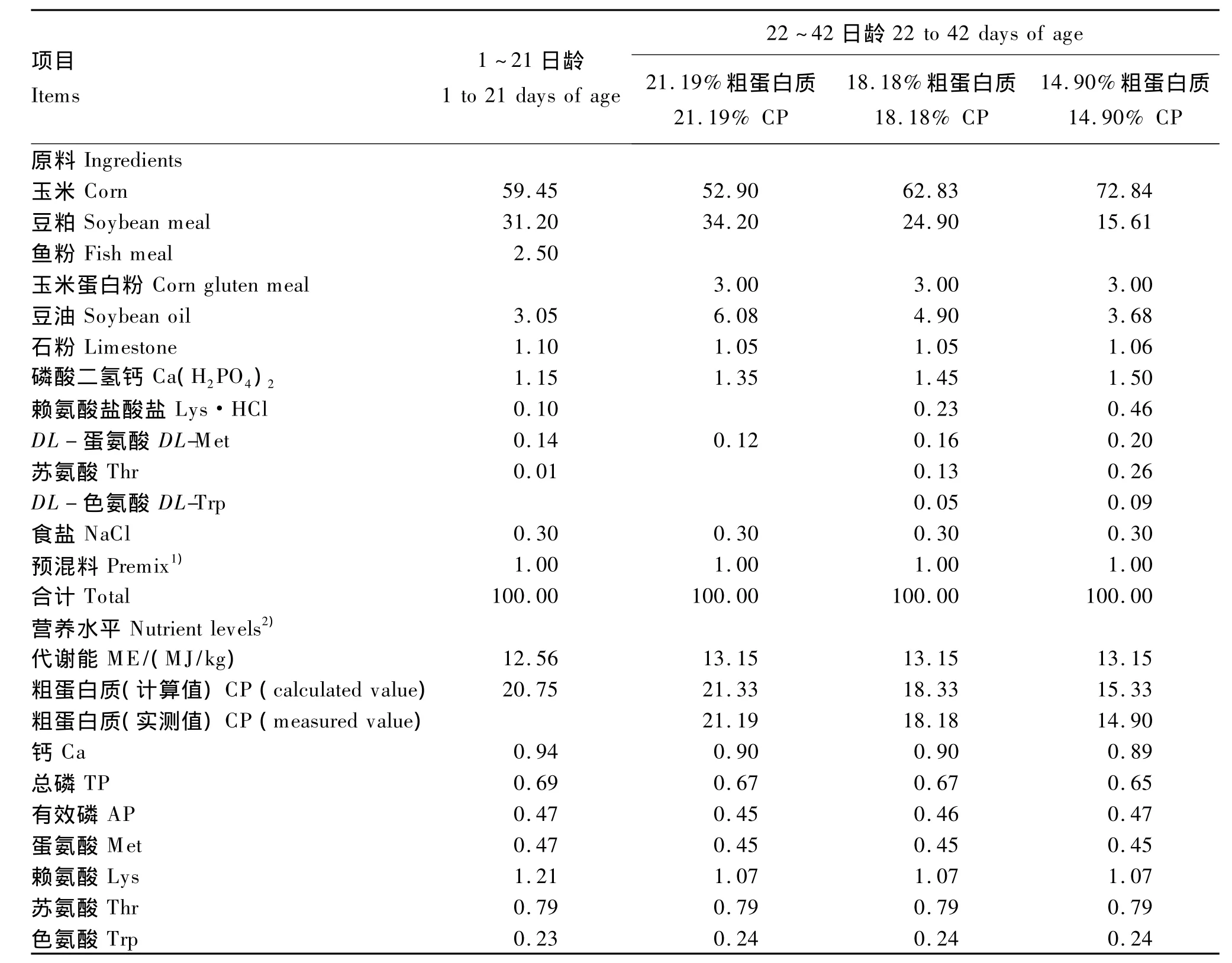
表2 试验饲粮组成及营养水平(风干基础)Table 2 Composition and nutrient levels of experimental diets(air-dry basis)%
1.3.3 血清生化指标
用10 mL无抗凝剂真空采血管和3 mL肝素钠真空采血管各1支,在翅下静脉采血,无抗凝剂真空采血管摆斜面静置4 h,再3 500 r/min离心15 min分离析出的血清,-20℃保存待测。采用两点动力法测定血清尿素氮(urea nitrogen,UN)含量;采用酶比色法测血清尿酸(uric acid,UA)含量;采用双缩脲法测定血清总蛋白(total protein,TP)含量;采用南京建成生物工程研究所的黄嘌呤氧化酶(xanthine oxidase,XOD)测定试剂盒测定血清XOD活性,所用仪器是为日立7020全自动生化分析仪。
1.3.4 排泄物鲜样生化指标
在35、42日龄,以重复为单位收集24 h(每4 h收集1次)全部肉鸡新鲜排泄物,除去毛屑杂物,称重并一式2份,其中一份按照每100 g排泄物样品中加入10 mL 10%H2SO4固氮,最后将24 h收集的排泄物混匀,四分法取样后密封-20℃保存。固氮的排泄物样品在65℃烘干、回潮后称重,粉碎过40目筛,制成风干样品。另一份新鲜排泄物样品用于分析测定UN、UA含量,方法同血清分析方法,所用仪器是为日立7020全自动生化分析仪。单位采食量尿酸(uric acid per feed intake,UADG)、单位采食量尿素氮(urea nitrogen per feed intake,UNFI)含量,用如下方式计算:

1.3.5 氮利用率和排泄物氮排放量
在35、42日龄,采样方法同排泄物鲜样生化指标。用凯氏定氮法测定排泄物风干样品的氮含量,氮利用率(nitrogen utilization,NU)、单位体增重氮排放量(nitrogen excretion per weight gain,NEWG)、单位采食量氮排放量(nitrogen excretion per feed intake,NEFI)用如下方式计算:


1.4 数据处理统计分析
用SAS 9.1软件中的一般线性模型(general linear model,GLM)做双因子方差分析,同时用Duncan氏法多重比较进行差异显著性分析,以P<0.05为差异显著水平。
2 结果与分析
2.1 环境高温与饲粮CP水平对肉鸡生产性能的影响
由表3可知,在相同饲粮CP水平下,28~42日龄,28/32℃ AL组肉鸡 BW、ADG、ADFI均显著低于23℃ AL组(P<0.01),而与23℃ PF组差异不显著(P>0.05),可见环境高温会降低28~42日龄肉鸡的生产性能,且主要是由热应激引起的采食量降低导致的;在相同饲养方式下,21.19%CP组F/G显著低于低CP组(18.18%CP组和14.90%CP组)(P<0.01),对其他生产性能指标影响不显著(P>0.05)。环境高温与饲粮CP水平对死亡率的影响都不显著(P>0.05)。

表3 环境高温与饲粮CP水平对28~42日龄肉鸡生产性能的影响Table 3 Effects of high temperature and dietary CP level on performance of broilers aged 28 to 42 days

续表3
2.2 环境高温与饲粮CP水平对肉鸡生理指标的影响
由表4可知,28/32℃ AL组肉鸡 RT为42.50℃,显著高于23℃ AL组的40.85℃和23℃ PF组的40.11℃(P<0.01);23℃ PF组的RT显著低于23℃ AL组(P<0.01)。
28/32℃ AL组肉鸡RR升高到240.96 br/min,显著高于23℃ AL组的51.70 br/min和23℃PF组的47.16 br/min(P<0.01),同时23℃ PF组的RR也有低于23℃ AL组的趋势(P<0.10)。环境高温与饲粮CP水平对肉鸡RR影响方面存在显著的互作效应(P<0.05),32℃高温下,饲粮CP水平为14.90%时,肉鸡RR显著高于高CP水平(21.19%和18.18%)时(P<0.05)。
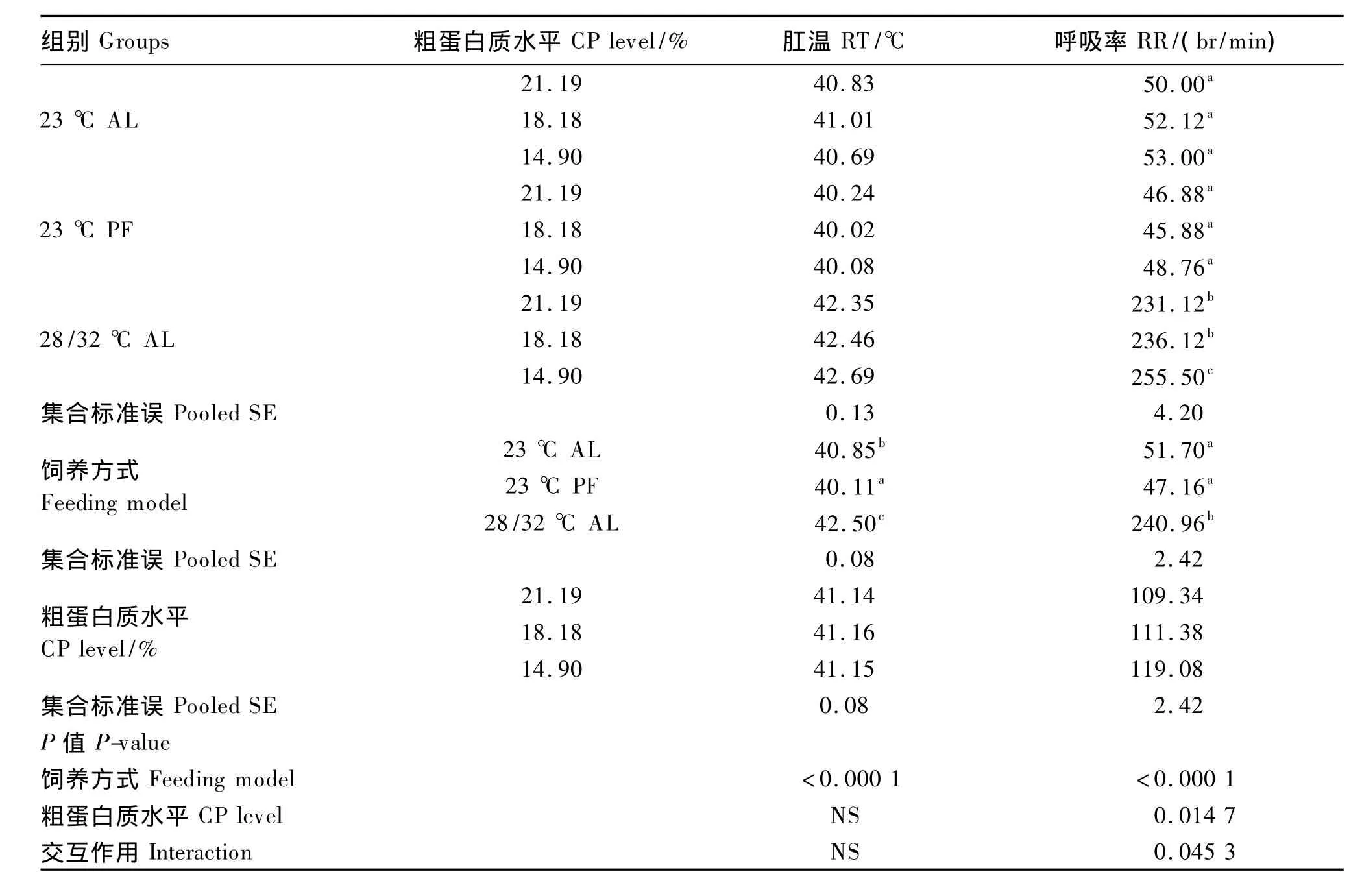
表4 环境高温与饲粮CP水平对28~42日龄肉鸡生理指标的影响Table 4 Effects of high temperature and dietary CP level on physiological indices in broilers aged 28 to 42 days
2.3 环境高温与饲粮CP水平对肉鸡血清生化指标的影响
由表5可知,35日龄时,28/32℃ AL组肉鸡血清UA含量显著高于23℃ AL组和23℃ PF组(P<0.05),23℃ AL组和23℃ PF组之间差异不显著(P>0.05),可见血清UA含量是受环境高温直接影响的,与采食量无关。42日龄时,28/32℃ AL组肉鸡血清TP含量与23℃ AL组无显著差异(P>0.05),但显著低于23℃ PF组(P<0.01);28/32℃ AL组肉鸡血清XOD活性显著高于23℃ AL组(P<0.01),而与23℃ PF组无显著差异(P>0.05)。
35日龄时,18.18%CP组肉鸡血清UA含量显著高于14.90%CP组(P<0.05),与21.19%CP组无显著差异(P>0.05);随着饲粮CP水平变化,血清UN、TP含量和XOD活性无显著变化(P>0.05)。42日龄时,18.18%CP组血清XOD活性显著高于21.19%CP组、14.90%CP组(P<0.05);随着饲粮 CP水平变化,血清 UA、UN、TP含量无显著变化(P>0.05)。

表5 环境高温与饲粮CP水平对35、42日龄肉鸡血清生化指标的影响Table 5 Effects of high temperature and dietary CP level on serum biochemical indices in broilers aged 35 and 42 days
2.4 环境高温与饲粮CP水平对肉鸡排泄物生化指标的影响
由表6可知,35、42日龄肉鸡的排泄物中,28/32℃AL组单位采食量尿酸含量比23℃ AL组显著增加(P<0.05),而23℃ PF组与前2组差异都不显著(P>0.05);28/32℃ AL组单位采食量尿素氮含量比23℃ AL组和23℃ PF组均显著增加(P<0.05)。
35、42日龄时,与21.19%CP组相比,低CP组的单位采食量尿酸、单位采食量尿素氮(除35日龄的18.18%CP组外)含量均显著降低(P<0.05),但2个低CP组之间差异不显著(P>0.05)。
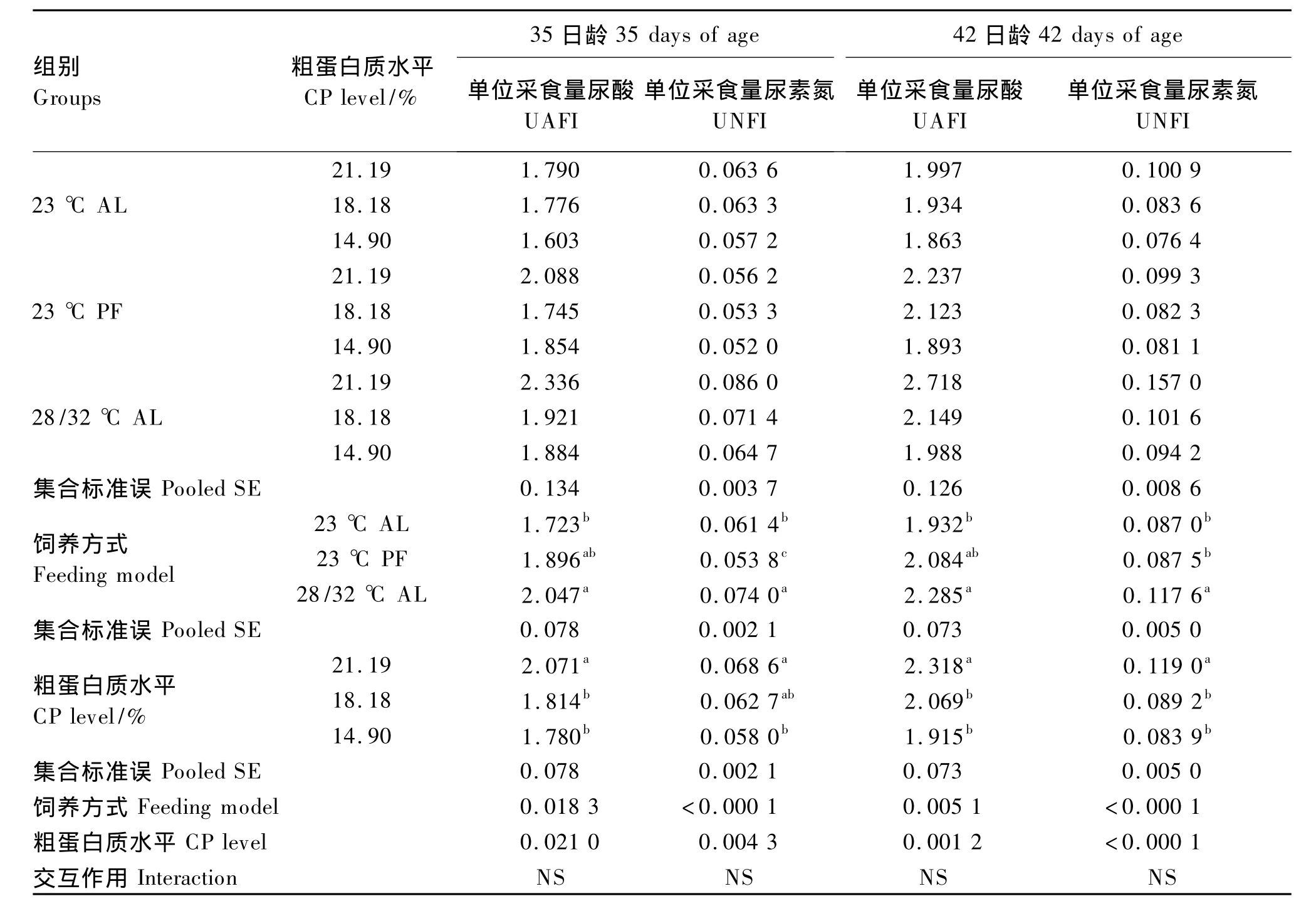
表6 环境高温与饲粮CP水平对35、42日龄肉鸡排泄物生化指标的影响Table 6 Effects of high temperature and dietary CP level on excreta biochemical indices in broilers aged 35 and 42 days mg/g
2.5 环境高温与饲粮CP水平对肉鸡氮排放量的影响
由表7可知,在相同饲粮CP水平下,35日龄时,与23℃ AL组相比,28/32℃ AL组氮利用率显著降低(P<0.01),单位体增重氮排放量(P<0.01)和单位采食量氮排放量(P<0.05)显著增加;而23℃ AL组氮利用率、单位采食量氮排放量均与23℃ PF组差异不显著(P>0.05)。42日龄时,与23℃ AL组相比,28/32℃ AL组氮利用率显著降低(P<0.01)、单位采食量氮排放量显著增加(P<0.05);而28/32℃ AL组肉鸡氮利用率、单位采食量氮排放量均与23℃ PF组差异不显著(P>0.05)。
在相同饲养方式下,随着饲粮CP水平由21.19%下降到14.90%,35、42日龄肉鸡单位体增重氮排放量和单位采食量氮排放量均显著减少(P<0.01),而氮利用率变化不显著(P>0.05)。可见,无论在23℃或者28/32℃时,补充人工合成EAA的同时降低饲粮CP水平可以显著减少氮排放。
3 讨论
3.1 环境高温与饲粮CP水平对肉鸡生产性能的影响
研究表明,32℃以上环境高温会显著降低肉鸡体增重[1]、采食量和生长率[2]。配对试验发现,热应激引起肉鸡生长抑制主要是由于采食量减少而导致的[15]。Smith 等[16]报道,在 35 ℃ 高温下,与自由采食相比,强饲额外16%的饲粮,肉鸡的屠宰率和生产性能呈现显著增长。本试验结果表明,28/32℃ AL组的 28~42日龄肉鸡 BW、ADG、ADFI均显著低于23℃ AL组,而与23℃PF组差异不显著,可见环境高温会抑制生产性能,而这主要是由热应激引起的采食量降低导致的。

表7 环境高温与饲粮CP水平对35、42日龄肉鸡氮排放量的影响Table 7 Effects of high temperature and dietary CP level on nitrogen excretion in broilers aged 35 and 42 days %
研究发现,当降低饲粮CP水平并补充EAA时,Alleman等[17]认为虽然不影响生长率,但 F/G有增加的趋势;而Holsheimer等[18]认为不影响肉鸡生产性能。本试验结果中,随着饲粮CP水平由21.19%下降到14.90%,除F/G显著升高外,其余生产性能指标没有显著变化。可见,无论在23℃或者28/32℃时,在降低饲粮CP水平的同时补充人工合成EAA,不会显著降低肉鸡BW、ADG、ADFI。在生产性能方面,环境高温与饲粮CP水平不存在互作效应。
3.2 环境高温与饲粮CP水平对肉鸡RT和RR的影响
环境高温引起肉鸡热应激,RT、RR会显著升高[19],24、35 ℃时的 RT 与采食量呈线性相关[20],产热量会随着饲喂水平增加[21],本试验中,28/32℃ AL组RT显著高于23℃ AL组和23℃PF组;23℃ PF组的RT显著低于23℃ AL组。Toyomizu等[22]研究报道,肉鸡暴露在38℃高温下 60 min,RR由 61 br/min增加到 261 br/min。本试验中,环境温度达32℃时,42日龄肉鸡RR显著高于23℃ AL组的51.70 br/min,同时显著高于23℃ PF组的47.16 br/min,说明高温下RT、RR升高是受到温度直接影响的,而与高温下采食量降低无关。环境高温与饲粮CP水平在RR上存在互作效应,高温下,14.90%CP组RR显著高于18.18%CP组、21.19%CP组,同时也高于显著高于23℃ AL组和23℃ PF组,估计是低CP水平使代谢减缓的缘故。
本试验中,28/32℃ AL组肉鸡RT、RR显著高于23℃ AL组和23℃ PF组,可见环境高温引起了肉鸡热应激。
3.3 环境高温与饲粮CP水平对肉鸡血清生化指标的影响
家禽合成尿素的能力较弱,氮代谢主要通过嘌呤核苷酸循环,以尿酸作为嘌呤碱代谢的终产物。吴永胜等[23]指出,血清UN含量减少,TP含量提高,表明蛋白质合成增加,动物生长率和饲料转化率高。有研究表明,热应激使肉鸡血清TP含量显著降低,UA、UN 含量显著升高[5,7],这可能是体蛋白合成减少、分解加强,UA排泄增加的结果。UA对清除体内自由基、抵抗热应激引起的氧化应激具有重要作用[24]。有报道,32.5/34.5℃循环高温下10 d,肉鸡血清 TP含量显著低于22℃组[25]。本试验发现,28/32℃下7 d(35日龄时),血清TP含量与23℃下差异不显著,但热应激14 d(42日龄时),TP含量显著低于23℃ PF组,且稍低于23℃ AL组;28/32℃下7 d,血清UA含量显著高于23℃ AL组和23℃ PF组,但14 d血清UA含量差异不显著,这很可能是后期肉鸡机体逐渐适应高温环境所致。
禽类UA需经过XOD的连续氧化作用生成,血清XOD主要由肝脏合成,而直接影响着UA含量,高温环境下,肉鸡受到热应激刺激产生大量自由基[1]。本试验42日龄时,28/32℃ AL组血清XOD活性显著高于23℃ AL组,但与23℃ PF组差异不显著。可见,肉鸡受到热应激影响显著,且可能与采食量下降有关。
De Silva等[26]报道,随着饲粮蛋白质水平由9.6%增加到30.4%,水产动物对蛋白质的消化率显著增加而后有降低趋势。随着饲粮CP水平降低,猪的血浆UN含量显著线性降低[27],氮的利用率显著增加。本试验发现,14.90%CP组的35日龄肉鸡血清UA含量显著低于21.19%CP组、18.18%CP组,可能与低CP组体内蛋白质代谢加强有关。
3.4 环境高温与饲粮CP水平对肉鸡排泄物生化指标的影响
随着环境温度升高,畜禽消化道内蛋白质消化和氨基酸吸收发生改变,蛋白质分解代谢加强、合成减弱[5,7],使得饲粮 CP 消化吸收减少、氮排泄增加。UA是禽类含氮物质代谢终产物,排泄物中含氮物质70% ~80%是以UA形式排放,那么高温下UA排放量显著增加。本试验中,35、42日龄,28/32℃ AL组单位采食量尿酸含量比23℃AL组显著增加,而23℃ PF组与前2组差异都不显著;28/32℃ AL组单位采食量尿素氮含量比23℃ AL组和23℃ PF组显著增加。可见,高温环境导致排泄物单位采食量尿酸增加,是受到热应激和采食量降低的共同影响;而单位采食量尿素氮增加,主要受到热应激影响,而与采食量无关。而环境温度升高使畜禽对蛋白质消化吸收减弱、氮排放增加。
研究报道,环境温度为24℃时,随着饲粮CP水平下降,肉鸡排泄物中UA含量、水分和酸度会下降[12],从而减少含氮化合物代谢终产物的生成和排放。本试验中,在相同温度情况下,补充了合成EAA后,与21.19%CP组相比,低CP组35、42日龄肉鸡的单位采食量尿酸、单位采食量尿素氮均显著降低,但2个低CP组之间差异不显著。可见,环境高温下,通过补充合成EAA、降低饲粮CP水平,同样可以显著降低肉鸡含氮化合物代谢终产物的生成和排放。而在体内氮代谢方面,环境高温和饲粮CP水平不存在互作效应。
3.5 环境高温与饲粮CP水平对肉鸡氮排放量的影响
Chwalibog等[10]认为,环境温度在 15 ~40 ℃的范围内变化不影响21~42日龄肉鸡体蛋白含量,而 Geraert等[8]发现,32 ℃热应激肉鸡体蛋白沉积率会显著低于22℃组。本试验血清指标中,28/32℃ AL组35日龄肉鸡血清UA含量显著高于23℃ AL组和23℃ PF组,42日龄肉鸡血清XOD活性显著高于23℃ AL组,可见环境高温下肉鸡蛋白质分解加强、合成减弱,可能导致氮排放增加。检测氮排放相关指标发现,在相同饲粮CP水平下,与23℃ AL组相比,28/32℃ AL组氮利用率显著降低,单位采食量氮排放量显著增加。但42日龄时,单位体增重氮排放量随环境温度变化无显著变化,推测是由于后期肉鸡对高温环境产生耐受导致。在相同采食量下,35日龄时28/32℃ AL组肉鸡氮利用率显著减少,单位采食量氮排放量显著增加;而42日龄时差异不显著。由此推测,35日龄肉鸡主要受环境温度影响而氮利用率减少、排泄量增加,42日龄时肉鸡对环境高温产生耐受,采食量的影响开始显著。
有研究表明,环境温度为21~24℃时,通过补充合成EAA,降低饲粮CP水平会显著减少畜禽氮排放量[11-13]。Keshavarz 等[13]研究报道,与饲喂16.0% ~16.5%CP的相比,饲喂13.0%CP并添加EAA饲粮的蛋鸡每日氮排泄量可以降低45%,而产蛋性能不会下降。本试验中,相同饲养方式下,无论在23℃或者28/32℃时,随着饲粮CP水平由21.19%降低到14.90%,氮利用率无显著变化,35、42日龄单位体增重氮排泄量和单位采食量氮排泄量均显著下降。可见,在环境高温下,通过改变蛋白质供给模式,即补充合成EAA、降低饲粮CP水平,同样可以显著降低氮排泄量。且在氮排放方面,环境高温与饲粮CP水平不存在互作效应。
4 结论
①日变循环高温导致肉鸡生产性能、氮利用率显著降低,氮排放增加。
②在日变循环高温条件下,补充人工合成EAA,饲粮CP水平降低到14.90%,可以减少氮排放而不影响肉鸡BW、ADG、ADFI。
③在生产性能、氮代谢和氮排放方面,环境高温与饲粮CP水平不存在互作效应。
[1]MUJAHID A,YOSHIKI Y,AKIBA Y,et al.Superoxide radical production in chicken skeletal muscle induced by acute heat stress[J].Poultry Science,2005,84(2):307-314.
[2]KLEIBER M,DOUGHERTY J E.The influence of environmental temperature on the utilization of food energy in baby chicks[J].The Journal of General Physiology,1934,17(5):701-726.
[3]OENEMA O,TAMMINGA S.Nitrogen in global animal production and management options for improving nitrogen use efficiency[J].Science in China Series C:Life Sciences,2005,48(S2):871-887.
[4]林海,杜荣.环境温度对肉鸡消化道内食糜排空和消化酶活性的影响[J].动物营养学报,2001,13(1):49-53.
[5]OSTROWSKI-MEISSNER H T.The physiological and biochemical responses of broilers exposed to short-term thermal stress[J].Comparative Biochemistry and Physiology Part A:Physiology,1981,70(1):1-8.
[6]DIBNER JJ,ATWELL C A,IVEY F J.Effect of heat stress on 2-hydroxy-4-(methylthio)butanoic acid and DL-methionine absorption measured in vitro[J].Poultry Science,1992,71(11):1900-1910.
[7]TEMIM S,CHAGNEAU A M,PERESSON R,et al.Chronic heat exposure alters protein turnover of three different skeletal muscles in finishing broiler chickens fed 20%or 25%protein diets[J].The Journal of Nutrition,2000,130(4):813-819.
[8]GERAERT P A,PADILHA J C F,GUILLAUMIN S.Metabolic and endocrine changes induced by chronic heat exposure in broiler chickens:growth performance,body composition and energy retention[J].British Journal of Nutrition,1996,75(2):195-204.
[9]WILSON E K,PIERSON F W,HESTER P Y,et al.The effects of high environmental temperature on feed passage time and performance traits of White Pekin ducks[J].Poultry Science,1980,59(10):2322-2330.
[10]CHWALIBOG A,EGGUM B O.Effect of temperature on performance,heat production,evaporative heat loss and body composition in chickens[J].Archiv für Geflügelkunde,1989,53(4):179-184.
[11]ZERVAS S,ZIJLSTRA R T.Effects of dietary protein and fermentable fiber on nitrogen excretion patterns and plasma urea in grower pigs[J].Journal of Animal Science,2002,80(12):3247-3256.
[12]NAMROUD N F,SHIVAZAD M,ZAGHARI M.Effects of fortifying low crude protein diet with crystalline amino acids on performance,blood ammonia level,and excreta characteristics of broiler chicks[J].Poultry Science,2008,87(11):2250-2258.
[13]KESHAVARZ K,AUSTIC R E.The use of low-protein,low-phosphorus,amino acid and phytase-supplemented diets on laying hen performance and nitrogen and phosphorus excretion[J].Poultry Science,2004,83(1):75-83.
[14]LENIS N P,VAN DIEPEN H T,BIKKER P,et al.Effect of the ratio between essential and nonessential amino acids in the diet on utilization of nitrogen and amino acids by growing pigs[J].Journal of Animal Science,1999,77(7):1777-1787.
[15]DALE N M,FULLER H L.Effect of diet composition on feed intake and growth of chicks under heat stress.Ⅱ.Constant vs.cycling temperatures[J].Poultry Science,1980,59(7):1434-1441.
[16]SMITH M O,TEETER R G.Influence of feed intake and ambient temperature stress on the relative yield of broiler parts[J].Nutrition Reports International,1987,35(2):299-306.
[17]ALLEMAN F,LECLERCQ B.Effect of dietary protein and environmental temperature on growth performance and water consumption of male broilerchickens[J].British Poultry Science,1997,38(5):607-610.
[18]HOLSHEIMER J P,JANSSEN W M M A.Limiting amino acids in low protein maize-soybean meal diets fed to broiler chicks from 3 to 7 weeks of age[J].British Poultry Science,1991,32(1):151-158.
[19]BORGES S A,DA SILVA A V F,MAJORKA A,et al.Physiological responses of broiler chickens to heat stress and dietary electrolyte balance(sodium plus potassium minus chloride,milliequivalents per kilogram)[J].Poultry Science,2004,83(9):1551-1558.
[20]TEETER R G,SMITH M O,WIERNUSZ C J.Research note:broiler acclimation to heat distress and feed intake effects on body temperature in birds exposed to thermoneutral and high ambient temperatures[J].Poultry Science,1992,71(6):1101-1104.
[21]WIERNUSZ C J,TEETER R G.Feeding effects on broiler thermobalance during thermoneutral and high ambient temperature exposure[J].Poultry Science,1993,72(10):1917-1924.
[22]TOYOMIZU M,TOKUDA M,MUJAHID A,et al.Progressive alteration to core temperature,respiration and blood acid-base balance in broiler chickens exposed to acute heat stress[J].The Journal of Poultry Science,2005,42(2):110-118.
[23]吴永胜,董国忠,王立常,等.饲粮中添加铬-烟酸复合物对肉鸭生产性能、胴体成分及血液生化指标的影响[J].动物营养学报,2000,12(2):20-25.
[24]王讯,马恒东,赵玲,等.鸡体内尿酸生物学功能的研究进展[J].动物医学进展,2005,26(3):41-43.
[25]刘思当,宁章勇,谭勋,等.热应激对肉仔鸡血液生化指标影响的观察[J].中国兽医杂志,2003,39(9):20-23.
[26]DE SILVA S S,PERERA M K.Digestibility in Sarotherodon niloticus fry:effect of dietary protein level and salinity with further observations on variability in daily digestibility[J].Aquaculture,1984,38(4):293-306.
[27]NYACHOTI C M, OMOGBENIGUN F O,RADEMACHER M,et al.Performance responses and indices of gastrointestinal health in early-weaned pigs fed low-protein amino acid-supplemented diets[J].Journal of Animal Science,2006,84(1):125-134.
猜你喜欢
动物营养学报(2022年9期)2022-10-12
动物营养学报(2022年8期)2022-09-01
今日农业(2021年7期)2021-11-27
中国饲料(2021年17期)2021-11-02
煤气与热力(2021年6期)2021-07-28
今日农业(2020年18期)2020-12-14
高师理科学刊(2020年2期)2020-11-26
广东饲料(2016年6期)2016-12-01
中国国情国力(2016年1期)2016-11-26
少儿科学周刊·少年版(2015年11期)2015-12-17
