
北长山岛森林乔木层碳储量及其影响因子
2013-09-19 03:05石洪华王晓丽刘振英麻德明
生态学报 2013年19期
石洪华 ,王晓丽,王 嫒,刘振英,麻德明
(1.国家海洋局第一海洋研究所,青岛 266061;2.天津理工大学环境科学与安全工程学院,天津 300384)
森林是陆地生态系统的重要组成部分。森林生态系统作为重要的碳储存库,约占整个陆地生态系统碳库的50%,在储存CO2、缓解全球气候变化过程发挥着重要的作用[1-3]。乔木层、灌草层、枯落物层和土壤层碳储量共同组成森林生态系统碳储量,其中森林乔木层固碳能力不容忽视。由于受到立地条件、根系与土壤的生理生化作用等因素的影响,森林土壤与乔木层碳储量具有密切的相关性[4-5]。估算森林乔木层碳储量,探寻出森林乔木碳储量与土壤环境的相关性,已成为森林生态系统碳循环研究的热点问题。
以往对陆地生态系统的固碳评估研究多集中在农田、陆域森林等领域[4,6-8],而对海岛陆地生态系统固碳研究较少。海岛兼具海、陆双重特性[9],立地条件差,树种相对单一,土壤贫瘠,时常面临大风、海啸和风暴潮等威胁,通常采用大面积人工造林改善海岛生态环境[10-11]。黑松(P.thunbergii)适应性强,具有喜海洋气候、抗海风、耐瘠薄等特性,是山东省长岛县的主要造林树种。海岛人工林的生态环境脆弱,由于海岛四面环海,植被对大风、海啸等影响没有任何阻挡和缓冲,在同等级别的自然灾害下与陆域相比,其受影响的程度往往更大。单一树种对改善立地条件,以及发挥森林多种防护效益都很有限,抵抗病虫害和各种自然灾害的能力较弱。因此,为了丰富海岛森林生态系统类型的多样性,长岛县北长山岛人工林建设主要采用黑松纯林与黑松×刺槐(P.thunbergii×R.pseudoacacia)混交林两种模式,确保海岛人工林生态和防护效能的正常发挥。
本研究以北长山岛黑松、黑松×刺槐混交林为研究对象,利用生物量相对生长方程与样地调查数据相结合的方法,分别研究了黑松纯林与黑松×刺槐混交林乔木层碳储量,利用多元统计方法分析影响海岛森林乔木层碳储量的因子,旨在为庙岛群岛陆地植被固碳能力的评估提供参考资料。
1 材料与方法
1.1 研究区域概况
北长山岛(37°57'36″—37°59'40″N,120°40'50″—120°43'54″E)位于山东省长岛县,面积 8.25 km2,海岸线长 15.68 km,地貌以低丘陵为主,是庙岛群岛第二大岛。该地区属于东亚季风型气候区,年均气温为12.0℃,1月平均气温最低为-1.6℃,8月平均气温最高为24.5℃。年均降水量537.1 mm,降水多集中在6—9月。日照充足,年均日照时数2612 h。土壤主要有棕壤、褐土、潮土三大类,以棕壤土分布面积最大,土层厚度约为30 cm,多砂砾,土质较差。植被类型主要是松树及槐树林,以松树为主,该岛松林面积为庙岛群岛之首。从植被分布来看,下部主要是松树和以松、槐为主的阔叶针叶混交林。黄土坡及沟谷内,以槐树为主的阔叶针叶混交林。林下分布各种灌木、耐碱草本植物,尚有果林和农作物分布。该岛现有林木绝大部分为1949年以后人工植种,目前森林覆盖率约为42.0%,黑松和黑松×刺槐混交林为该岛主要森林类型。
1.2 样地设置
2012年7月,根据北长山岛植被分布特征,依据代表性和可达性原则,共设置13个调查样方,样地面积一般为20 m×20 m,由于立地条件所限制个别样地面积设置为10 m×10 m(图1)。各样地的生境特征见表1。分别调查统计森林立地条件及各样地内乔木的胸径、高度、基径、枝下高、冠幅、株数等指标。
1.3 研究方法
1.3.1 人工林固碳能力评估
根据文献[12-13]中给定的相对生长方程适用范围、模拟效果等因素,选择适合本研究区域内黑松和刺槐生物量相对生长方程(表2),计算出北长山岛黑松、黑松×刺槐混交林乔木层各器官的生物量和总生物量。

图1 北长山岛森林乔木层调查样方分布Fig.1 Locations of the sample plots in the Beichangshan Island

表1 样地基本概况Table 1 The basic characteristics of the sampling stations
植被生物量乘以含碳率可得到植被碳储量。在估算森林生态系统植被碳储量的研究中,国内外研究者通常采用0.50或0.45的生物量与碳的换算系数,该值在不同树种间变化范围为0.4—0.5之间[14-17]。通过查阅文献,本文选择的生物量与碳的换算系数为 0.45[12,14,18]。
1.3.2 土壤样品采集及分析
在每个样方内分别选取3个取样点,采用多点混合取土样方法进行。北长山岛森林土壤砂砾较多,土壤厚度约为30 cm,森林土壤取表层土。在每个取样点采用标准环刀(100 cm3)进行取样,然后将环刀内土壤在105℃高温下烘至恒重,称量并计算土壤含水率和土壤容重。将同一样地内3个点取到的土壤均匀混合,作为该样地的土壤样品。样品采集后,带回实验室,自然风干,除去其中草根、大石砾等杂质,过2 mm钢筛,磨细待测。
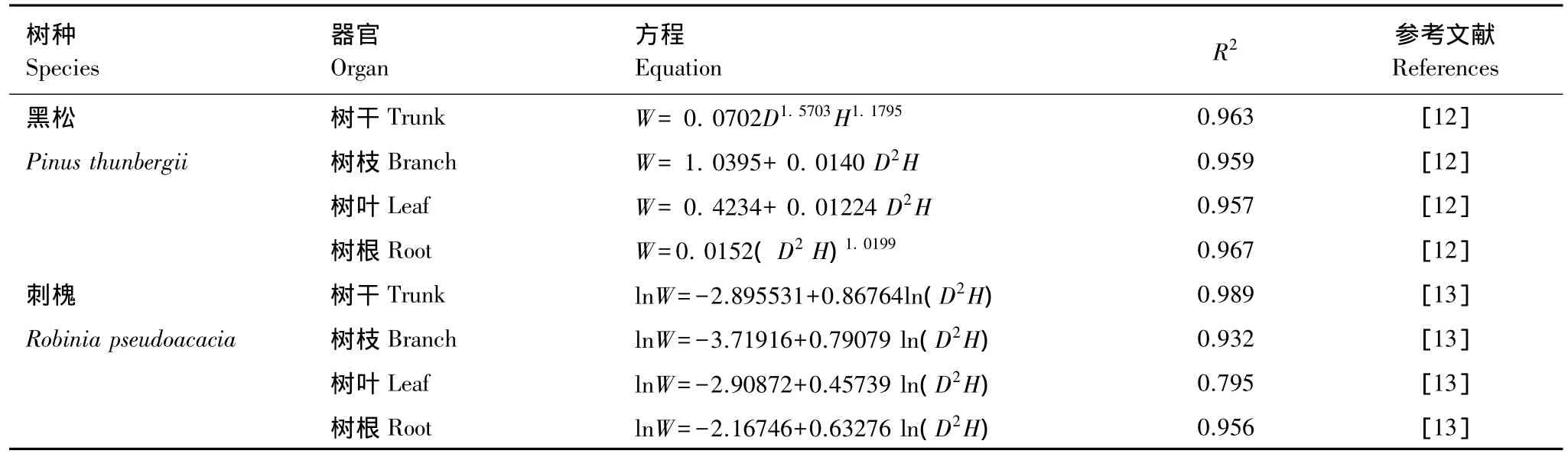
表2 样树各器官生物量(W)与胸径(D)、树高(H)的相对生长方程Table 2 Organ biomass allometric equations with variances of diameter at breast height(D)and height(H)of P.thunbergii and R.pseudoacacia
土壤理化性质按照中国土壤学会编写的土壤农业化学分析方法进行[19]。土壤质地和土壤比表面积采用激光粒度分析仪(Mastersizer 2000,Malvern Instruments Co.,UK)测定;土壤有机碳含量采用灼烧法测定[20]。森林植物群落土壤级配指标,即不均匀系数Cu与曲率系数Cs计算采用如下公式[21]:

式中,Cu为不均匀系数;d10、d60为土壤颗粒累积百分含量为10%、60%所对应的粒径。Cs为曲率系数;d30为土壤颗粒累积百分含量为30%所对应的粒径。
为便于分析立地条件对海岛森林乔木层碳储量影响,本研究对坡向进行转换。在转换过程中,设定以正南方向为0°,正西、正东分别为-90°、90°,坡向转换系数计算采用如下公式:

式中,C为坡向转换系数;α为坡向偏角
1.4 数据处理和统计分析
采用单因素方差分析(one-way ANOVA)比较两种树种及各器官之间的差异;采用因子分析(Factor Analysis Method)提取乔木层碳储量主要影响因子。所有数据采用SPSS18.0进行处理分析。
2 结果与分析
2.1 乔木层碳储量
在北长山岛人工林中,黑松林各器官的碳储量(除树皮外)均高于黑松×刺槐混交林(表3)。两种树种各器官碳储量之间的差异明显(P<0.05),均以树干最高,树根次之,树枝和树叶最低,说明树干碳储量的积累是乔木层碳储量增加的重要影响因素。
黑松的乔木层碳储量显著高于黑松×刺槐混交林。两种林型乔木不同器官的碳储量均为:树干>树根>树枝>树叶(表3)。北长山岛黑松纯林乔木层碳储量为84.00 t/hm2,接近于世界平均水平(86.00 t/hm2)[22],高于广西西南桦乔木碳储量28.541 t/hm2[23],高于长沙市区马尾松林乔木层碳储量32.42 t/hm2[24];黑松×刺槐混交林碳储量为29.60 t/hm2,略高于广西西南桦×红椎混交林乔木层碳贮量29.144 t/hm2[23]。
表3 黑松、黑松×刺槐混交林乔木层碳储量(±sD)Table 3 Carbon storage in P.thunbergii and P.thunbergii×R.Pseudoacacia(±sD)

表3 黑松、黑松×刺槐混交林乔木层碳储量(±sD)Table 3 Carbon storage in P.thunbergii and P.thunbergii×R.Pseudoacacia(±sD)
组分Component黑松P.thunbergii黑松×刺槐P.thunbergii×R.pseudoacacia碳储量/(t/hm2)Carbon storage /%碳储量/(t/hm2)Carbon storage /%树干 Trunk 33.77±22.86 40.21 13.42±8.80 45.34树枝 Branch 16.27±12.02 19.37 5.42±3.51 18.30树叶 Leaf 13.97±10.51 16.63 3.89±2.64 13.14根系 Root 19.99±15.37 23.80 6.87±4.53 23.23总计Total 84.00 100.00 29.60 100.00
2.2 土壤对森林乔木层碳储量的影响
2.2.1 森林土壤质地对乔木层碳储量的影响
北长山岛土壤颗粒(<2 mm)砂粒含量变化范围为82.31%—90.90%,粉砂粒含量变化范围为9.02%—17.38%,黏粒含量较低,变化范围为0.05%—0.30%,土壤类型以砂土为主(表4)。研究区域颗粒级配指标颗粒不均匀系数Cu与曲率系数Cs同时满足Cu>5且Cs在1—3范围内,森林土壤级配良好;北长山岛黑松、黑松×刺槐混交林土壤颗粒分散度Se较低,偏度Srk大(表5),颗粒分布不均衡,反映了2种森林表层土壤流失程度相对较高。由于北长山岛森林土壤含石率(>2 mm)较高(质量百分比约为30%—50%),土壤颗粒相对石头而言含量较少,而地表侵蚀时较细颗粒首先流失,北长山岛森林土壤虽级配良好但土壤流失程度较高。
森林土壤与植被生长的作用是相互的。北长山岛黑松纯林土壤质地部分为砂质壤土,而黑松×刺槐混交林土壤均为砂土(表4)。在土壤改良方面黑松纯林可能要优于黑松×刺槐混交林;且黑松林固碳能力高于黑松×刺槐混交林,是庙岛群岛不可替代的、具有良好生态功能的理想树种。

表4 北长山岛表层土壤质地Table 4 Soil texture in the Beichangshan island

表5 土壤颗粒(<2 mm)级配状况及群体特征值Table 5 The soil structure and population characteristics in the Beichangshan island
为找出海岛森林乔木层碳储量影响因子,以不同的环境因子(坡度、海拔)及土壤颗粒含量为参数计算出相关系数矩阵(表6)。乔木层碳储量与环境因子及土壤颗粒含量具有相关性。因子分析提取3个影响因子,因子1中土壤砂粒含量(-0.898)、粉砂粒含量(0.893)、黏粒含量(0.922),因子2中坡度(0.770)、坡向(-0.722)、海拔(0.946)具有较高的负载,因子3中坡向(0.350)、砂粒含量(-0.385)、粉砂粒含量(0.390)具有较高的负载(表7)。由于因子3中各因素负载均小于0.5,与因子1、2相比较低,因此只对因子1、因子2进行多元线性回归分析。多元线性回归分析表明,对乔木层碳储量的影响中因子1(Beta=0.290)相较于因子2(Beta=0.019)更为重要。
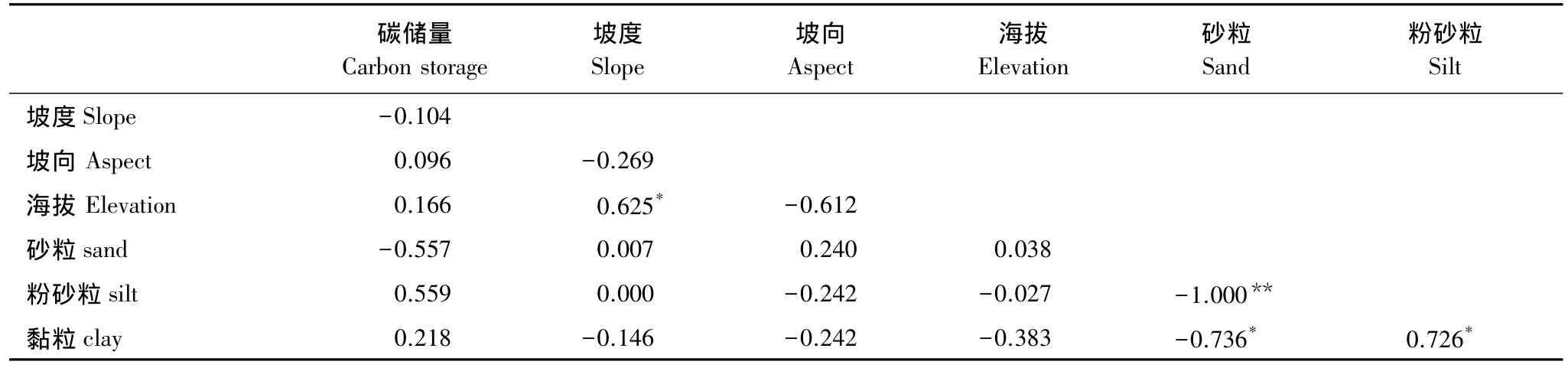
表6 森林乔木碳储量与土壤质地、环境因子相关系数矩阵Table 6 Correlation coefficient matrix of arbores carbon storage and different soil texture and environmental parameters

表7 乔木层碳储量控制变量因子分析旋转因子负荷矩阵Table 7 Rotated component matrix derived from a factor analysis of variables controlling arbore carbon storage
2.2.2 土壤理化性质对森林乔木层碳储量的影响
土壤养分状况是影响植物生长、繁殖和生态系统固碳能力等的重要因素[5,25-26]。由图2可知,就北长山岛表层土而言,所测定的土壤理化指标均存在显著差异(P<0.001),这种差异是由不同树种生理特性以及林下环境特征所引起。森林群落冠层具有遮阴作用,北长山岛土壤含水量变化范围为1.29%—3.51%。森林土壤的8个监测样地中,有7个样地土壤偏酸性,pH值变化范围为4.99—8.2。由于森林土壤表层覆盖有部分凋落物,北长山岛森林土壤有机碳含量变化范围为10.56—17.81 g/kg,普遍高于北长山岛上的农田和村落土壤,主要是因为森林生态系统土壤表面覆盖有凋落物,导致森林土壤有机质含量高于其他类型土壤。北长山岛虽为海岛,但其土壤含盐量较低,变化范围为0.04%—0.206%。造成这种差异的原因可能为海岛土壤含石率高,土质松散,加之调查期(7月份)降雨充足,使得土壤盐分渗入深层土壤,从而其表层土盐分含量较低;另一方面,海岛人工造林可截留部分海盐,缓解海盐胁迫,对改良土壤具有一定的显著效果[10]。
以森林土壤理化性质为参数计算出相关系数矩阵(表8),找出森林乔木层碳储量影响因子。乔木层碳储量与土壤具有密切的相关性。因子分析提取两个影响因子,因子1中土壤全氮(0.842)、总有机碳(0.899)、全碳(0.990),因子2中全磷(0.931)、碳氮比(-0.925),因子3中含水量(0.694)、pH值(0.744)、含盐量(-0.666)具有较高的负载(表9)。多元线性回归分析表明,对乔木层碳储量的影响中因子3(Beta=-0.694)最为重要,其次为因子1(Beta=-0.192)、因子2(Beta=-0.106)。
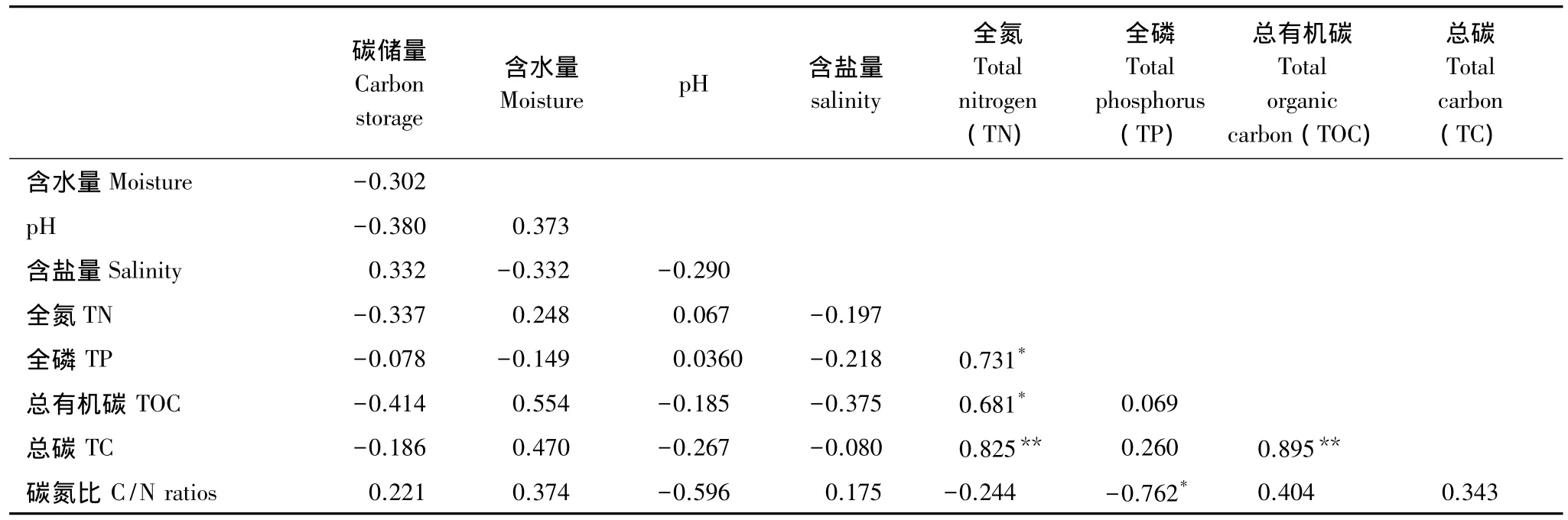
表8 森林乔木碳储量与土壤理化性质相关系数矩阵Table 8 Correlation coefficient matrix of arbores carbon storage and different soil physical-chemical properties

图2 森林土壤理化性质Fig.2 Soil physical-chemical properties in the forest of Beichangshan Island

表9 乔木层碳储量控制变量主成分分析旋转因子负荷矩阵Table 9 Rotated component matrix derived from a factor analysis of variables controlling arbore carbon storage
3 结论与讨论
本研究采用生物量相对生长方程与样地调查数据相结合的方法,分别研究了黑松纯林与黑松×刺槐混交林乔木层碳储量。北长山岛黑松纯林乔木层碳储量高于黑松×刺槐混交林,也高于其它地方的一些常见树种,造成这种差异的主要原因是黑松适应性强,具有喜海洋气候、抗海风、耐瘠薄等特性,较其他树种更适宜在海岛地区生长,因此具有较高的碳储量。
乔木层碳储量受到降雨、气温、坡度、海拔等多种环境因子的影响[27-29]。降雨量等气候特征对土壤母质的风化、土壤形成和土壤有机碳的分解起着重要的作用,进而影响森林乔木层碳储量[30-31]。Brown等[32]的研究结果,降水量在400—3200 mm时,降水和森林植被碳密度之间呈正相关,超过3200 mm时则呈负相关。北长山岛地区气候湿润,北长山岛降雨多集中7—8月份,充沛的降雨量促进土壤碳的分解进而加快了碳周转速率。有研究表明,海岛地区森林乔木层碳储量随坡度等级的增加而增大[27]。由于受到海风的胁迫,雨水冲刷等作用导致了山坳地区土壤厚度大于山顶,加之山坳中水热条件较好,土壤不易流失,利于植被的生长,森林乔木层具有较高的碳储量。
森林土壤对乔木层碳储量影响较大。森林乔木层碳储量与土壤质地密切相关,不同质地土壤特性不同。土壤颗粒组成与土壤养分相关[33-34],对植被的生长、发育具有重要作用,进而影响森林乔木层碳储量。由于海岛地区植被、土壤时常受到海风的影响,局部地段形成风沙堆积。北长山岛森林土壤黏粒含量极低,均在5%以下,土壤类型以砂土类为主(表4),部分为壤土类,级配良好(表5)。砂土类土壤通透性良好,但持水能力较低,微生物活性强,进而促进土壤氮矿化,有利于植被根系对养分的吸收,进而提高了树木的光合速率。黑松纯林、黑松×刺槐混交林乔木层根系碳储量分别为19.99、6.87 t/hm2(表3),占到各自乔木层总碳储量的1/5,说明植被根系对土壤养分吸收较好,根系较为发达,这与土壤质地密不可分。
土壤理化性质通过影响根系与土壤的生理生化作用进而影响植被的生长。氮、磷是植物生长所必须的大量营养元素,其丰缺及供给状况直接影响着植物的生长水平。北长山岛土壤全N、全P含量较低(图2)。全N含量均低于2 g/kg,而在意大利0—20 cm表层土的21个监测样地中,仅有2个样地的土壤全N含量低于2 g/kg[35],更是低于日本22a柳杉林土壤全N含量(2.03 g/kg)[36],因此,北长山岛森林土壤全N含量水平低于国外其他国家的土壤全N水平,对乔木层碳储量影响不是非常明显(表8)。凋落物是土壤-植物系统碳循环的联结库,对表层土壤养分循环产生重要影响[37]。通过现场调查发现,北长山岛森林林下凋落物覆盖少,这也是导致北长山岛森林土壤全N含量较低的原因之一。由图2可知,对于同种林型的森林土壤N、P含量低,总有机碳相对较高(17.81 g/kg),森林乔木具有较高的碳储量(122.91 t/hm2)。因此,土壤中有机碳和养分的合理、协调增加有利于根系的生长,进而提高森林乔木层碳储量。
森林系统水分循环是植被赖以生存的重要环节之一。土壤水更是植被生长、发育所需用水的主要来源,但过高的含水量会导致植被根系的腐烂不利于植被的生长。大多数土壤养分元素的有效性受土壤pH值影响[38-39]。由图2可知,北长山岛8个监测样地中只有1个样地土壤偏碱性(pH值=7.7)且森林乔木层碳储量较低(28.52 t/hm2)。北长山岛森林乔木层碳储量与土壤pH值基本呈负相关。因此,夏季适宜的土壤含水量及pH值有利于森林乔木碳储量的提高。
北长山岛夏季土壤含盐量较低(图2),部分黑松林土壤质地为砂质壤土,可见,海岛人工林的建设对于涵养淡水、过滤海盐、防风固沙等起到重要作用。虽然北长山岛黑松乔木层碳储量高于黑松×刺槐混交林,黑松更适宜在海岛地区生长,是海岛地区不可替代的、具有良好固碳功能的理想树种。单一树种在抵抗病虫害和自然灾害的能力较弱,为了丰富海岛森林生态系统类型的多样性,还应积极抚育其他适宜海岛地区生长的优良树种。混交林是一种较好的森林经营模式,合理的混交比例与种植密度可以提高资源的利用率,增加林分生长量进而提高生态系统的碳储量。
[1] Lorenz K,Lal R.Carbon Sequestration in Forest Ecosystems.New York:Springer-Verlag,2010.
[2] Dixon R K,Brown S,Solomon A M,Houghton R A,Trexier M C,Wisniewski J.Carbon pools and flux of global forest ecosystems.Science,1994,263(5144):185-190.
[3] Houghton R A.Balancing the global carbon budget.Annual Review of Earth and Planetary Sciences,2007,35(1):313-347.
[4] Wang W X,Shi Z M,Luo D,Liu S R,Lu L H,Ming A G,Yu H L.Carbon and nitrogen storage under different plantations in subtropical south China.Acta Ecologica Sinica,2013,33(3):925-933.
[5] Hu H F,Liu G H.Dynamics of soil physical-chemical properties and organic carbon content along a restoration chronosequence in Pinus tabulaeformis plantations.Acta Ecologica Sinica,2013,33(4):1212-1218.
[6] Moroni M T.Aspects of forest carbon management in Australia:A discussion paper.Forest Ecology and Management,2012,275:111-116.
[7] Nunery J S,Keeton W S.Forest carbon storage in the northeastern United States:Net effects of harvesting frequency,post-harvest retention,and wood products.Forest Ecology and Management,2010,259(8):1363-1375.
[8] Sharma C M,Baduni N P,Gairola S,Ghildiyal S K,Suyal S.Tree diversity and carbon stocks of some major forest types of Garhwal Himalaya,India.Forest Ecology and Management,2010,260(12):2170-2179.
[9] Shi H H,Zheng W,Ding D W,Lü J B.Valuation and ecosystem services of typical island.Marine Environmental Science,2009,28(6):743-748.
[10] Han G X,Zhang Z D,Wang G M,Mao P L,Liu S J,Xue Q Z.Growth dynamics and quantitative population characteristics of young trees in coastal Pinus thunbergii windbreak forest in northern Shandong Peninsula.Chinese.Journal of Ecology,2009,28(6):1013-1020.
[11] Donato D C,Kauffman J B,Mackenzie R A,Ainsworth A,Pfleeger A Z.Whole-island carbon stocks in the tropical Pacific:Implications for mangrove conservation and upland restoration.Journal of Environmental Management,2012,97:89-96.
[12] Xu J W,Wang W D,Wang Y H,Li C R,Kang Z.The mathematical model estimating the biomass of black pine plantation in sandy coastal.Journal of Shandong Forestry Science and Technology,2004,154(5):47-49.
[13] Bi J,Huang Z Z,Wang Z L.Studies on biomass dynamic of black loclust tree.Journal of Hebei Forestry College,1993,8(4):278-282.
[14] Wang X K,Feng Z W,Ouyang Z Y.Vegetation carbon storage and density of forest ecosystems in China.Chinese Journal of Applied Ecology,2001,12(1):13-16.
[15] Yim M H,Joo S J,Nakane K.Comparison of field methods for measuring soil respiration:A static alkali absorption method and two dynamic closed chamber methods.Forest Ecology and Management,2002,170(1/3):189-197.
[16] Liu H,Hou L,Lei R D.Carbon storage and carbon density of Pinus tabulaeformis and Pinus armandii forests at Huoditang forest region in Qinling Mountain.Chinese Journal of Eco-Agriculture,2007,15(1):5-8.
[17] Fang J Y,Chen A P,Peng C H,Zhao S Q,Ci L J.Changes in forest biomass carbon storage in China between 1949 and 1998.Science,2001,292(5525):2320-2332.
[18] Crutzen P J,Andreae M O.Biomass burning in tropics:impact on the atmospheric chemistry and biogeochemical cycles.Science,1990,250(4988):1669-1678.
[19] Lu R K.Analytical Methods of Soil Agrochemistry.Beijing:Chinese Agriculture Science and Technology Press,1999.
[20] Wiesmeier M,Prietzel J,Barthold F,Spörlein P,Geuß U,Hangen E,Reischl A,Schilling B,von Lützow M,Kögel-K I.Storage and drivers of organic carbon in forest soils of southeast Germany(Bavaria)-Implications for carbon sequestration.Forest Ecology and Management,2013,295:162-172.
[21] Liu X,Wang L,Zhang G C,Liu X R,Liang H.Structural property of soil from different forest types on rocky mountainous area in middle of Shandong Province.Journal of Soil Water Conservation,2005,12(6):49-52.
[22] Liu G H,Fu B J,Fang J Y.Viewpoints on the study of economic thresholds of cotton bollworm.Acta Ecologica Sinica,2000,20(5):733-740.
[23] He Y J,Qin L,Li Z Y,Shao M X,Liang X Y,Tan L.Carbon storage capacity of a Betula alnoides stand and a mixed Betula alnoides×Castanopsis hystrix stand in Southern Subtropical China:a comparison study.Acta Ecologica Sinica,2012,32(23):7586-7594.
[24] Wu T,Peng C H,Tian D L,Yan W D.Spatial distribution of carbon storage in a 13-year-old Pinus massoniana forest ecosystem in Changsha City,China.Acta Ecologica Sinica,2012,32(13):4034-4042.
[25] Bassem D,Jean-Pierrre C,Jerome L,Bruno M.Changes in soil carbon and nitrogen following tillage conversion in a long-term experiment in Northern France.Agriculture,Ecosystems and Environment,2013,169:12-20.
[26] Allison S D,Gartner T B,Mack M C,McGuire K,Treseder K.Nitrogen alters carbon dynamics during early succession in boreal forest.Soil Biology& Biochemistry,2010,42(7):1157-1164.
[27] Wang X L,Wang A,Shi H H,Peng S T,Gong L P,Qin X B.Carbon storage of Pinus thunbergii and Robinia pseudoacacia plantations on Nanchangshan Island,Changdao County of Shandong Province,China.Chinese Journal of Applied Ecology,2013,24(5):1263-1268.
[28] Wu D,Shao Q Q,Li J,Liu J Y.Carbon fixation estimation for the main plantation forest species in the red soil hilly region of southern-central Jiangxi Province,China.Acta Ecologica Sinica,2012,32(1):142-150.
[29] Watanabe Y,Masunaga T,Owusu-Sekyere E,Buri M M,Oladele O I,Wakatsuki T.Evaluation of growth and carbon storage as influenced by soil chemical properties and moisture on teak(Tectona grandis)in Ashanti region,Ghana.Journal of Food Agriculture& Environment,2009,7(2):640-645.
[30] Renan U G,Natali H,Ruben M,Àngels X,Yuri Y.Forest management for timber and carbon sequestration in the presence of climate change:The case of Pinus Sylvestris.Ecological Economics,2013,88:88-96.
[31] Chen S T,Huang,Y,Zou J W,Shi Y S.Mean residence time of global topsoil organic carbon depends on temperature,precipitation and soil nitrogen.Global and Planetary Change,2013,100:99-108.
[32] Brown S,Lugo A E.Biomass of tropical forests:a new estimate based on forest volumes.Science,1984,223(4642):1290-1293.
[33] Haile S G,Nair V D,Nair P K R.Contribution of trees to carbon storage in soils of silvopastoral systems in Florida,USA.Global Change Biology,2010,16(1):427-438.
[34] Harrison-Kirk T,Beare M H,Meenken E D,Condron L M.Soil organic matter and texture affect responses to dry/wet cycles:Effects on carbon dioxide and nitrous oxide emissions.Soil Biology& Biochemistry,2013,57:43-55.
[35] Allaniello F,Biondi F A,Ferrari C,Mecella G,Nisini L.Forest soil conditions in the CONECOFOR permanent monitoring plots and in the level I network in Italy.Journal of Limnology,2002,61(S1):25-35.
[36] Duan W X,Zhu B,Lu J H,Liu R,Chen F.Dynamics of carbon storage and soil properties in Japanese cedar(cryptomeria fortunei)plantations.Chinese Journal of Applied and Environmental Biology,2007,13(6):777-781.
[37] Wang H Y,Wang W J,Qiu L,Su D X,An J,Zheng G Y,Zu Y Y.Differences in biomass,litter layer mass and SOC storage changing with tree growth in Larix gmelinii plantations in Northeast China.Acta Ecologica Sinica,2012,32(3):833-843.
[38] Wang H,Xing X J,Xu Z C.Relationship between pH value and available nutrients of purple soil in Panxi tobacco-growing areas.Soil and Fertilizer Sciences in China,2007,(6):19-22,49-49.
[39] Khattak R A,Hussain Z.Evaluation of soil fertility status and nutrition of orchards.Soil& Environment,2007,26(1):22-32.
参考文献:
[4] 王卫霞,史作民,罗达,刘世荣,卢立华,明安刚,于浩龙.我国南亚热带几种人工林生态系统碳氮储量.生态学报,2013,33(3):925-933.
[5] 胡会峰,刘国华.人工油松林恢复过程中土壤理化性质及有机碳含量的变化特征.生态学报,2013,33(4):1212-1218.
[9] 石洪华,郑伟,丁德文,吕吉斌.典型海岛生态系统服务及价值评估.海洋环境科学,2009,28(6):743-748.
[10] 韩广轩,张志东,王光美,毛培利,刘苏静,薛钦昭.山东半岛北部黑松海防林幼龄植株生长动态及其种群数量特征.生态学杂志,2009,28(6):1013-1020.
[12] 许景伟,王卫东,王月海,李传荣,康智.沿海沙质岸黑松人工林生物量的估测数学模型.山东林业科技,2004,154(5):47-49.
[13] 毕君,黄则舟,王振亮.刺槐单株生物量动态研究.河北林学院学报,1993,8(4):278-282.
[14] 王效科,冯宗炜,欧阳志云.中国森林生态系统的植物碳储量和碳密度研究.应用生态学报,2001,12(1):13-16.
[16] 刘华,侯琳,雷瑞德.秦岭火地塘林区油松和华山松林的空间分布格局及碳储量与碳密度研究.中国生态农业学报,2007,15(1):5-8.
[19] 鲁如坤.土壤农业化学分析方法.北京:中国农业科技出版社,1999.
[21] 刘霞,王丽,张光灿,刘信儒,梁华.鲁中石质山地不同林分类型土壤结构特征.水土保持学报,2005,12(6):49-52.
[22] 刘国华,付博杰,方精云.中国森林碳动态及其对全球碳平衡的贡献.生态学报,2000,20(5):733-740.
[23] 何友均,覃林,李智勇,邵梅香,梁星云,谭玲.西南桦纯林与西南桦×红椎混交林碳贮量比较.生态学报,2012,32(23):7586-7594.
[24] 巫涛,彭重华,田大伦,闫文德.长沙市区马尾松人工林生态系统碳储量及其空间分布.生态学报,2012,32(13):4034-4042.
[27] 王晓丽,王嫒,石洪华,彭士涛,宫立鹏,覃雪波.山东省长岛县南长山岛黑松和刺槐人工林的碳储量.应用生态学报,2013,24(5):1263-1268.
[28] 吴丹,邵全琴,李佳,刘纪远.江西中南部红壤丘陵区主要造林树种碳固定估算.生态学报,2012,32(1):142-150.
[37] 王洪岩,王文杰,邱岭,苏冬雪,安静,郑广宇,祖元刚.兴安落叶松林生物量、地表枯落物量及土壤有机碳储量随林分生长的变化差异.生态学报,2012,32(3):833-843.
[38] 王晖,邢小军,许自成.攀西烟区紫色土pH值与土壤养分的相关分析.中国土壤与肥料,2007,(6):19-22,49-49.
猜你喜欢
美食(2022年12期)2022-12-09
现代园艺(2020年2期)2020-12-22
美食(2019年7期)2019-09-10
小溪流(故事作文)(2018年10期)2018-12-28
现代园艺(2018年2期)2018-03-15
科学大众·小诺贝尔(2018年12期)2018-02-22
山东林业科技(2016年5期)2016-07-05
亚热带资源与环境学报(2015年4期)2015-01-22
东北师大学报(自然科学版)(2014年1期)2014-02-27
亚热带资源与环境学报(2014年3期)2014-01-23
