
植酸酶对红鳍东方鲀幼鱼生长、消化酶及消化率的影响
2013-09-19 09:07孟祥科孙阳屈菲姜志强毛明光李艳秋吴洪
大连海洋大学学报 2013年4期
孟祥科,孙阳,屈菲,姜志强,毛明光,李艳秋,吴洪
(1.大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连116023;2.辽宁出入境检验检疫局通关处,辽宁大连116001)
近年来,随着鱼粉价格的升高,用植物性蛋白作为鱼粉的替代品得到广泛关注[1-2],但是植物蛋白中存在的多种抗营养因子严重阻碍了水产动物对于营养物质的有效利用[3]。植酸广泛存在于植物性原料中,是一种抗营养因子,它能够无选择性地螯合蛋白,抑制一些酶的活力包括蛋白酶、胰蛋白酶和α-淀粉酶等。鱼类养殖试验已证明[4-7],植酸能够降低磷的利用率,影响鱼类的生长、体组成和饲料效率。植酸在大多数加工过程中不容易被去除,而添加植酸酶可以水解饲料中的植酸和植酸盐,促进动物对磷的吸收,减少磷排放,改善动物生长性能。
近年来,随着畜禽动物饲料中植酸酶应用研究的逐渐深入,水产动物饲料中添加植酸酶的研究报道也相继增加。研究表明,饲料中添加植酸酶能显著改善点带石斑鱼[4]、真鲷[5]、大西洋鲑[6]等鱼类的生长及饲料效率,但饲料中添加植酸酶对红鳍东方鲀Takifugu rubripes生长性能及饲料利用等的研究目前却鲜见报道。红鳍东方鲀营养丰富、肉味鲜美,是一种名贵的海水鱼类。本研究中,以红鳍东方鲀幼鱼为试验对象,在以添加豆粕降低鱼粉用量的基础上添加不同水平的植酸酶,研究植酸酶对东方鲀幼鱼生长、消化酶及消化率的影响,以期为植酸酶在红鳍东方鲀饲料中的合理利用提供参考。
1 材料与方法
1.1 材料
试验用红鳍东方鲀幼鱼购自大连富谷水产有限公司。试验用植酸酶购自山东潍坊苏柯汉生物工程有限公司 (酶活力为5 000 U/g)。试验共制作6组饲料。对照组D1,全鱼粉蛋白组;对照组D2,以豆粕替代30%鱼粉蛋白组;试验组S1~S4,在豆粕替代30%鱼粉蛋白基础上设4个不同的植酸酶添加水平 (500、1 000、1 500、2 000 U/kg)。植酸酶活性定义为:在37℃、pH为5.5反应条件下,从5.0 mmol/L植酸钠溶液中每分钟释放1 μmol/L无机正磷酸根所需植酸酶的量为1个酶活力单位 (U)。
主要原料经粉碎研磨后过80目网筛,混合均匀后用制粒机挤压成软颗粒饲料,饲料规格随鱼的生长阶段而定。饲料分批制作:试验初期饲料粒径为1 mm;待幼鱼平均体质量为10~20 g时调整粒径为2 mm;幼鱼平均体质量大于20 g后再调整粒径为3 mm。制成的饲料于冰箱 (-20℃)中保存备用。试验饲料的组成及营养成分见表1。

表1 试验饲料营养成分Tab.1 The ingredients and appromimate composition of the experimental diets(dry matter)
1.2 方法
1.2.1 饲养管理 试验幼鱼先暂养20 d,期间投喂鱼糜,之后投喂基础饲料驯养10 d,待幼鱼完全适应养殖环境后开始正式试验。试验设6组,每组设3个重复,在18个200 L蓝色方形水槽中,分别投放初始体质量为 (4.58±0.05)g的幼鱼20尾。每天8:00和16:00投喂。投喂量以接近饱食为准。投饲半小时后吸底清除残饵。饲养期间采用流水养殖,水流流速约为3 L/min,24 h微充气。
试验在大连海洋大学农业部北方海水增养殖重点实验室进行,饲养期为60 d。养殖期间水温为20~24℃,溶解氧>5 mg/L,pH为8.0±0.1,盐度为32.6±0.2。
1.2.2 样品的收集 60 d饲养期结束后,停食24 h,从每个水槽中随机取5尾鱼,测定其体长、体质量等,然后在冰盘上解剖,迅速剪取肝胰脏、前肠中段,并用4℃预冷生理盐水冲洗干净,置于超低温冰箱 (-80℃)中保存备用。
在投喂试验饲料饲养20 d,试验鱼生活状态稳定后开始收集粪便。每次投喂后吸净槽底残饵,4~5 h后用虹吸法收集粪便,置于培养皿中于70℃下烘干,待测。粪便样品连续收集15 d。
1.2.3 测定及计算方法
1)生长指标。试验鱼特定生长率 (SGR)、饲料系数 (FCR)、蛋白质效率 (PER)、存活率的计算公式如下:
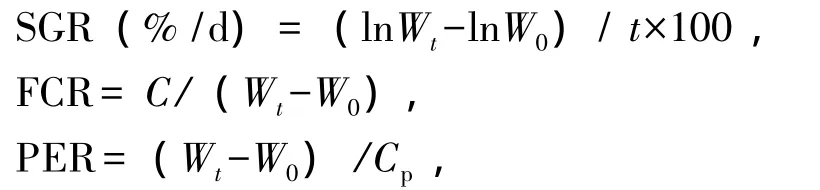
存活率 (%)=存活尾数/总尾数×100,
其中:W0、Wt为试验鱼的初始体质量 (g)和终末体质量 (g);t为试验时间 (d);C为摄饵量(g);Cp为蛋白质摄入量 (干质量,g)。
2)形体指标。试验鱼的肝体比 (HSI)、脏体比 (VSI)、肥满度 (CF)计算公式如下:
HSI(%)=肝脏质量/鱼体质量×100,
VSI(%)=内脏质量/鱼体质量×100,
CF=W/L3×100,
其中:W为试验鱼体质量(g);L为试验鱼长度(cm)。
3)粗酶液的制备。将存放于-80℃下的肝胰脏和肠组织样品在4℃下解冻,剪碎,按1 g组织加入9 mL生理盐水的比例加入预冷的生理盐水,在玻璃匀浆器中匀浆,粗酶液于4℃下以10 000 r/min离心10 min,取上清液备用。
用蛋白质试剂盒 (考马斯亮兰法)测定组织匀浆蛋白质含量。
采用福林-酚试剂法测定蛋白酶活力,其活性单位定义为:在37℃条件下,每分钟水解酪素所产生1 μg酪氨酸为一个酶活力单位 (U)。
用脂肪酶试剂盒 (比浊法)测定脂肪酶活性,其活性单位定义为:在37℃条件下,每克组织蛋白在本反应体系中与底物反应1 min,每消耗1 μmol底物为一个酶活力单位 (U)。
4)表观消化率。采用Cr2O3指示剂法测定消化率,饲料中指示剂掺入量为0.5%,采用湿式灰化定量法测定饲料和粪便中的Cr2O3含量。养分物质消化率 (D)和饲料干物质消化率 (D')的计算公式如下:

其中:A'和A分别为饵料和粪便中的营养物质含量;B和B'分别为饵料和粪便中的Cr2O3的含量。
1.3 数据处理
试验数据以平均值±标准差的形式表示,采用SPSS 16.0软件进行方差分析和Duncan多重比较,显著性水平设为0.05。
2 结果
2.1 植酸酶对红鳍东方鲀幼鱼生长性能的影响
由表2可以看出:经过60 d的饲养试验后,各组红鳍东方鲀幼鱼成活率没有显著差异 (P>0.05);相对于对照组D1,不同处理的豆粕替代30%鱼粉蛋白组 (D2,S1~S4)幼鱼的终末体质量、SGR、PER等均显著降低 (P<0.05),FCR明显升高 (P<0.05);相对于D2组,试验组 (S1~S4)幼鱼的生长性能整体提高,当植酸酶添加水平≥1 000 U/kg时,幼鱼的终末体质量、SGR明显提高 (P<0.05),而FCR则显著降低 (P<0.05)。

表2 植酸酶对红鳍东方鲀幼鱼生长性能的影响Tab.2 Effects of dietary phytase on growth performance in juvenile redfin puffer Takifugu rubripes
2.2 植酸酶对红鳍东方鲀幼鱼形体指标的影响
由表3可以看出:与D1组相比,D2组幼鱼HSI显著升高 (P<0.05);与D2组相比,添加植酸酶对红鳍东方鲀幼鱼HSI、VSI、CF均未产生显著影响 (P>0.05)。

表3 植酸酶对幼鱼肝体比、脏体比及肥满度的影响Tab.3 Effects of dietary phytase on hepato-somatic index(HSI),viscera-somatic index(VSI)and condition factor(CF)in juvenile redfin puffer Takifugu rubripes
2.3 植酸酶对红鳍东方鲀幼鱼消化酶活力的影响
由表4可以看出:与D1组相比,豆粕替代30%鱼粉蛋白后肝胰脏蛋白酶活力明显降低 (P<0.05)。随植酸酶添加量的增加,幼鱼肝胰脏蛋白酶活力呈增加趋势,当植酸酶添加量为1 000、1 500、2 000 U/kg时,肝胰脏中的蛋白酶活力较D2组显著升高 (P<0.05)。本研究结果表明,添加植酸酶能明显提高幼鱼肠道蛋白酶活力,且植酸酶添加量为1 000、1 500 U/kg时,幼鱼肠道蛋白酶活力较D2组分别提高20.33%、16.51%。
与D2组相比,添加植酸酶对各组幼鱼肝胰脏脂肪酶活力未产生显著影响 (P>0.05);随植酸酶的添加,幼鱼肠道脂肪酶活力呈先升高后降低的趋势,在植酸酶添加水平为1 500 U/kg时达到最高,与D1组差异不显著(P>0.05),但与D2组差异显著 (P<0.05)。
各组幼鱼肠道淀粉酶活力随植酸酶添加量的增加呈先升高后降低的趋势,在植酸酶添加量为500 U/kg时达到最高,与 D1组差异不显著 (P>0.05),但与D2组差异显著 (P<0.05);添加植酸梅对幼鱼肝胰脏淀粉酶活性未产生显著影响(P>0.05)。
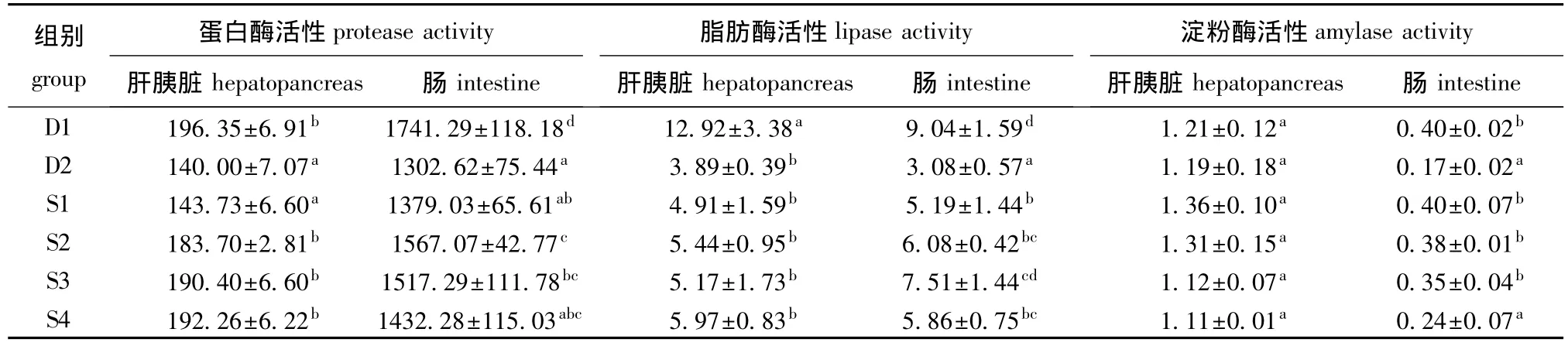
表4 植酸酶对红鳍东方鲀幼鱼消化酶活力的影响Tab.4 Effects of dietary phytase on digestive enzyme activities in juvenile redfin puffer Takifugu rubripes U/g
2.4 植酸酶对红鳍东方鲀幼鱼消化率的影响
由表5可知:红鳍东方鲀幼鱼对干物质及磷消化率较低,而对蛋白质和脂肪的消化率较高。添加植酸酶对幼鱼干物质消化率未产生显著影响 (P>0.05),但随植酸酶添加量的增加,干物质消化率呈升高的趋势,且在1 000 U/kg处达到最大值;植酸酶添加量为1 000 U/kg时,蛋白质消化率较D2组显著提高(P<0.05);植酸酶添加量为1 000、1 500 U/kg时,脂肪消化率较D2组显著提高(P<0.05);添加植酸酶后,磷消化率较D2组显著增加(P<0.05),植酸酶添加水平为500、1 000、1 500、2 000 U/kg时,磷消化率较D2组分别提高 39.55%、47.60%、56.89%、46.79%。

表5 植酸酶对红鳍东方鲀幼鱼消化率的影响Tab.5 Effects of dietary phytase on digestibility in juvenile redfin puffer Takifugu rubripes %
3 讨论
3.1 植酸酶水平对红鳍东方鲀生长性能的影响
有研究资料表明[4-8],在水产动物饲料中添加适量的植酸酶,可以提高其生长性能,促进其生长。植酸酶对水产动物的作用因试验动物种类、植酸酶的含量以及饲料成分等的差异,研究结果各有不同。马恒甲等[7]在低磷全植物性蛋白草鱼基础饲料中添加植酸酶的试验结果表明,添加大于1 000 U/kg的植酸酶可以明显提高草鱼的特定增长率、蛋白转化率,并显著降低饵料系数。陈新宇等[8]在研究植酸酶对凡纳滨对虾生长性能的影响时发现,在总磷含量为1.6%,植酸磷含量为0.3%,非植酸磷含量为1.3%的基础饲料中添加500~2 000 U/kg的植酸酶,对其增重率、饲料利用率和存活率均无显著影响,作者分析原因,可能是其试验基础饲料中的磷含量已经能满足凡纳滨对虾正常的生长需求。本试验结果显示,与D2组相比,饲料中添加1 000 U/kg的植酸酶,能显著提高红鳍东方鲀幼鱼的生长性能,证实了酸性植酸酶在红鳍东方鲀配合饲料中应用的可行性。通过本试验结果推测,试验组幼鱼的磷需求量要大于基础饲料中所提供的总磷水平 (1.47%)。
幼鱼消化系统发育不及成鱼完善,可能会对各种外界因子尤其是饵料因子变化的反应较敏感[9],尤其是肉食性鱼类对饵料蛋白要求高,若饲料中植物蛋白明显增加时,可能会对消化系统发育产生一定影响,肝体比会发生较大变化。本试验中,D2组肝体比与D1组相比明显增加,这与庄平等[10]和Kaushik等[11]研究豆粕替代鱼粉蛋白的试验结果一致。这可能是饲料中豆粕比例较高时,某些抗营养因子 (如植酸、皂苷、低聚糖类等)的存在,引起肝脏消化酶的分泌增加[12],加重了肝脏代谢负担,导致肝脏代偿性增大。本试验结果显示,植酸酶对红鳍东方鲀幼鱼HSI、VSI、CF指标没有显著影响,但随植酸酶添加量的增加,肝体比有一定的降低,但S1~S4组均与D1组差异不显著,说明饲料中添加植酸酶对防止肝脏肿大,降低肝脏负荷有一定作用,但效果不明显。
3.2 植酸酶对红鳍东方鲀幼鱼消化酶活力的影响
鱼类消化酶活力受诸多因素影响,其中包括饲料组成[13],植物性原料中的植酸对水产动物分泌的消化酶如淀粉水解酶、脂肪酶、蛋白水解酶等的活性均有抑制作用。添加植酸酶可以解除植酸对消化酶的抑制作用[14],促进营养物质的消化吸收。
红鳍东方鲀为肉食性鱼类,喜欢摄入蛋白质和脂类含量较高的食物,而且极为贪食,对营养物质有较高的消化和吸收能力。本试验结果显示,所有试验组肝胰脏及肠道蛋白酶活力均较高,且肠道蛋白酶活力高达1 302.62~1 741.29 U/g;相比D2组,试验组 (S1~S4)幼鱼肝胰脏及肠道蛋白酶活力整体提高,表明植酸酶能提高红鳍东方鲀幼鱼肝胰脏及肠道蛋白酶活力,这与朱丽英[15]关于植酸酶对异育银鲫生长、内源酶的影响,姚瑞清等[16]关于植酸酶对奥尼罗非鱼生长、表观消化率与消化酶活性的影响,以及张璐等[17]关于植酸酶和非淀粉多糖酶对鲈生长和消化酶活性的影响等的研究结果相似。
吕林兰等[18]关于植酸酶对异育银鲫的研究结果显示,添加1 000~1 500 U/kg的植酸酶能提高肝胰脏和肠道脂肪酶的活性。王枫[19]在对金鳟的研究中发现,植酸酶的添加导致金鳟肠道和肝胰脏脂肪酶活性略有降低,但差异不显著。本试验结果与上述研究结果不同:添加植酸酶虽然提高了红鳍东方鲀幼鱼肠道脂肪酶活力,但是未对其肝胰脏脂肪酶活力产生明显影响,这可能与肝胰脏脂肪酶主要以酶源的形式储存在肝胰脏中未被激活有关,具体原因有待进一步探究。
本试验中,各组肝胰脏及肠道淀粉酶活力均较低,很好地反映了红鳍东方鲀的食性特点。植酸酶添加量为500 U/kg时,肠道道淀粉酶活力达到最高,随添加量继续增加活力依次降低。相关研究显示,淀粉酶活力会因食物中某些金属离子浓度的提高而受到明显抑制[20]。本试验中,由于植酸酶在红鳍东方鲀幼鱼消化道内,阻碍了植酸与蛋白质的结合,并大量释放与植酸结合的金属离子,导致某些金属离子浓度过高,抑制了肠道淀粉酶活力,因此,会呈现肠道淀粉酶活力先升高后降低的趋势。这与牛继峰等[21]对大口黑鲈的研究结果一致。
3.3 植酸酶对红鳍东方鲀幼鱼消化率的影响
研究表明,当饲料中植物性蛋白质含量过高时,会显著降低鱼类的饲料利用率[22]。本试验结果显示,与D1组相比,D2、S1~S4组红鳍东方鲀幼鱼生长性能及消化率明显降低 (P<0.05),这与宋理平等[23]对宝石鲈及陈京华等[24]对牙鲆的研究结果相似。
植酸分子中的碱性磷酸基团能与蛋白质中质子化的氨基酸、金属离子结合生成螯合物,使蛋白质发生沉淀而降低其利用率及消化酶活力,从而影响饲料的利用率。磷是鱼类生长所必须的营养元素之一[25],植酸酶的添加可以打开饲料中植酸的磷酸磷脂键释放出无机磷,提高磷的利用率,同时消除与植酸结合的蛋白质,有利于动物对其消化吸收。
关于植酸酶对水产动物消化能力的影响,已有一些报道。宋理平等[23]研究显示,一定浓度的植酸酶可以明显提高宝石鲈的干物质、蛋白质、脂肪、钙、磷的表观消化率。舒秋艳等[26]指出,饲料中添加植酸酶可以一定程度上提高青鱼粗蛋白质、粗脂肪、粗灰分及钙表观消化率。本试验结果显示,饲料中添加植酸酶可提高红鳍东方鲀干物质、蛋白质、脂肪、磷消化率,表明植酸酶对其营养物质的利用有一定促进作用。因此,饲料中添加适量的植酸酶可以改善红鳍东方鲀的营养利用率,降低磷排放,节约饲料成本。
[1]Ai Q H,Xie X J.Effects of replacement of fish meal by soybean meal and supplementation of methionine in fish meal/soybean meal-based diets on growth performance of the southern catfish Silurus meridionalis[J].World Aqua Soc,2005,36:498-507.
[2]张娟娟,李小勤,冷向军,等.外源蛋白酶对虹鳟生长和肠道组织结构的影响[J].大连海洋大学学报,2012,27(6):534-537.
[3]Tacon A G J,Jackson A.Utilization of conventional and unconventional protein sources in practical fish feeds[M].Nutrition and feeding in fish.London:Academic Press,1985:211-264.
[4]阮成旭,袁重桂.植酸酶在点带石斑鱼配合饲料中的应用[J].福建水产,2005(1):13-16.
[5]Biswas A K,Kaku H,Ji S C,et al.Use of soybean meal and phytase for partial replacement of fish meal in the diet of red sea bream,Pagrus major[J].Aquaculture,2007,267:284-291.
[6]Sajjadi M,Carter C G.Effect of phytic acid and phytase on feed intake,growth,digestibility and trypsin activity in Atlantic salmon(Salmo salar L.)[J].Aquaculture Nutrition,2004,10:135-142.
[7]马恒甲,叶金云,郭建林,等.饲料中添加植酸酶对草鱼生长、体组成及各组织磷含量的影响[J].上海海洋大学学报,2011,20(6):845-851.
[8]陈新宇,潘庆,毕英佐,等.植酸酶对凡纳滨对虾生长性能及体成分的影响[J].湛江海洋大学学报,2005,25(6):14-18.
[9]崔敏,郭冉,夏辉.用饲料酵母替代鱼粉对大菱鲆幼鱼生长及免疫机能的影响[J].大连海洋大学学报,2012,27(1):58-63.
[10]庄平,陈喜斌,曾翠平,等.中华鲟幼鲟饲料中适宜动植物蛋白比的研究[J].动物营养学报,2002,14(1):61-64.
[11]Kaushik S J,Coves D,Dutto G,et al.Almost total replacement of fish meal by plant protein sources in the diet of a marine teleost,the European seabass Dicentrarchus labrax[J].Aquaculture,2004,230:391-404.
[12]Francis G,Makkar H P S,Becker K.Anti-nutritional factors present in plant-derived alternate fish feed ingredients and their effects in fish[J].Aquaculture,2001,199:197-227.
[13]李卫芬,沈涛,陈南南,等.饲料中添加枯草芽孢杆菌对草鱼消化酶活性和肠道菌群的影响[J].大连海洋大学学报,2012,27(3):221-225.
[14]祁艳霞,陈玉林.植酸酶的作用机理及影响植酸酶活性的因素[J].饲料博览,2004(7):10-11.
[15]朱丽英.植酸酶对异育银鲫生长、内源酶及肌肉成分的影响[D].南京:南京农业大学,2006.
[16]姚瑞清,刘波,吴婷婷,等.植酸酶对奥尼罗非鱼生长、表观消化率与消化酶活性的影响[J].中国饲料,2008(17):30-32.
[17]张璐,艾庆辉,麦康森,等.植酸酶和非淀粉多糖酶对鲈鱼生长和消化酶活性的影响[J].水生生物学报,2009,33(1):82-88.
[18]吕林兰,王爱民.植酸酶对异育银鲫鱼种生长及内源酶的影响[J].粮食与饲料工业,2007(3):36-37.
[19]王枫.植酸酶对金鳟营养物质表观消化率与消化酶活性的影响[D].哈尔滨:东北农业大学,2008.
[20]吴众望,潘鲁青,董双林.9种金属离子对缢蛏消化酶活力的影响[J].中国水产科学,2003,10:297-300.
[21]牛纪锋,吴锐全,谢骏,等.饲料中添加植酸酶对大口黑鲈生长和消化酶活性的影响[J].大连水产学院学报,2010,25(2):132-136.
[22]王广军,吴锐全,谢骏,等.军曹鱼饲料中用豆粕代替鱼粉的研究[J].大连水产学院学报,2005,20(4):304-307.
[23]宋理平,王爱英,张红.饲料中添加植酸酶对宝石鲈生长和饲料利用的影响[J].饲料工业,2010(增刊):83-88.
[24]陈京华,麦康森.不同添加方式植酸酶处理豆粕对牙鲆生长和饲料利用率的影响[J].水生生物学报,2010,34(3):481-488.
[25]孙丽慧,张利民,王际英,等.不同饲料对星斑川鲽幼鱼生长和养殖水环境的影响[J].大连海洋大学学报,2011,26(6):544-549.
[26]舒秋艳,黄峰,侯红利,等.中性耐温植酸酶对青鱼生长养分表观消化率及血清生化指标的影响[J].饲料工业,2010,31(8):12-15.
猜你喜欢
中国饲料(2022年13期)2022-07-07
当代水产(2021年9期)2021-12-02
当代水产(2021年8期)2021-11-04
医学前沿(2021年18期)2021-04-14
中国畜牧杂志(2020年11期)2020-11-18
国外畜牧学(猪与禽)(2020年8期)2020-03-04
生物技术通报(2019年7期)2019-02-21
中国畜禽种业(2018年1期)2018-01-18
畜牧兽医科技信息(2015年5期)2015-12-27
中国民族民间医药·下半月(2014年4期)2014-09-26
