
黄土高原不同侵蚀类型区生物结皮中蓝藻的多样性
2013-09-15 06:18:34杨丽娜赵允格王爱国
生态学报 2013年14期
杨丽娜,赵允格,* ,明 姣,王爱国
(1.中国科学院水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室,杨凌 712100;2.西北农林科技大学水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室,杨凌 712100;3.西北农林科技大学林学院,杨凌 712100)
生物土壤结皮(简称生物结皮),是指由藻类、苔藓、地衣、真菌及细菌等与土壤相互作用形成的有机复合体[1],是干旱半干旱地区普遍存在的地被物。土生藻类是指分布在土壤生境的藻类[2],是生物结皮的重要组分[3]。由于土生藻类能够在条件恶劣的环境下生长、繁殖,如干旱、营养贫瘠、高温、大风和强紫外线辐射等极端环境,通过自身的活动影响并改变着环境,因此,是重要的拓荒植物,具有许多重要的生态功能。土生藻类不仅可以通过自身的固氮作用和光合固碳作用,为土壤提供必要的营养物质,还可以通过藻丝体和分泌的多糖物质捆绑、黏结土壤颗粒,形成致密的抗蚀层,从而稳定和保护土壤表面免受水蚀风蚀影响[4]。蓝藻作为干旱半干旱地区土生藻类的优势类群,是主要的固氮类群,在生物结皮组成及生态功能中占据重要地位,尤其是占优势的丝状种类,不仅是重要的碳源、氮源,还能分泌胞外聚合物,黏结土壤颗粒,对土壤颗粒起到束缚作用;此外,蓝藻的藻丝体交织形成密集的网状结构在降雨出现时会迅速吸水、膨胀,能降低土壤的渗透性和地表的径流速度,显著提高了土壤抗侵蚀性能[5]。可见,蓝藻在干旱半干旱地区的生态环境的维持和改良过程中发挥着重要的作用[4,6-7]。
在我国的黄土高原地区,生物结皮是以退耕还林、封山禁牧为主的生态恢复工程的实施后广泛发育的地被物,盖度可以达到60%—70%[8]。近年来,有关该区的生物结皮及其生态功能已经展开了不少研究。如肖波,赵允格等[9-11]研究了生物结皮对土壤理化性质、导水入渗的影响,以及人工培育条件下生物结皮的养分积累与水土保持效应,研究发现生物结皮不仅对土壤养分有明显的累积作用,还可以降低土壤导水能力,减小水分入渗,并极显著地降低土壤侵蚀。此外,赵允格[12]研究了生物结皮光合作用对光温水的响应,发现在水分、光照充足的条件下,苔藓结皮的光合固碳能力显著高于藻结皮,认为不同类型的生物结皮光合固碳潜力主要取决于其生物组成。但是迄今有关该区生物结皮的生物组成的研究仍比较薄弱,生物结皮中藻类组成、分布及多样性鲜见报道。尤其是作为优势类群的蓝藻,它的种类组成、优势种方面的研究至今未有报道,蓝藻在不同侵蚀类型区生物结皮中的区系组成和分布情况如何,有无变化尚不清楚。一定程度上妨碍了人们对该区生物结皮生态功能的进一步认识。
为此,本文以黄土高原不同侵蚀类型区退耕撂荒地上自然发育的生物结皮为研究对象,通过野外调查、采样和室内培养、镜检相结合,初步分析了黄土高原不同侵蚀类型区生物结皮中蓝藻种类组成及优势种,旨在进一步理解黄土高原不同侵蚀类型区藻类在该区生态恢复中的重要作用,为生物结皮在水土流失和荒漠化防治中的应用奠定理论基础。
1 材料与方法
1.1 研究区概况
黄土高原位于北纬32°—41°,东经107°—114°之间。东起太行山,西到青海日月山,南界秦岭,北抵鄂尔多斯高原的区域,跨山西省、陕西省、甘肃省、青海省、宁夏回族自治区及河南省等省区,总面积64.87万km2,海拔1500—2000 m[13]。20世纪70年代,唐克丽等前辈在野外考察的基础上,初步将黄土高原按照主要的侵蚀营力划分为水蚀区、水蚀风蚀交错区及风蚀区[14]。其中,水蚀风蚀交错区位于北纬35°20'—40°10'、东经103°33'—113°53'之间,其范围大致自水蚀地区北部的神池、灵武、兴县、绥德、吴旗、庆阳、固原、定西、东乡一线以北到长城沿线以南一带,面积约17.8万km2,占黄土高原地区考察面积62.4万km2的28.56%。该区属半干旱草原地带,气候变化剧烈,植被稀疏,地形和产沙地层复杂,风沙地貌和流水侵蚀地貌交错分布,成为黄土高原土壤侵蚀最为强烈的地区。该区以北为风蚀区,其面积占总面积的25.08%;以南为水蚀区,其面积占总面积的 46.36%[14-15]。
1.2 调查及采样
样品采集于2011年8月进行,在黄土高原垂直等降雨量梯度线递减的方向从东南到西北分别选取水蚀、水蚀风蚀交错及风蚀侵蚀区的典型县(相对应的分别为陕西安塞县、吴起县、宁夏回族自治区盐池县),于各典型县选取封禁良好的小流域或退耕地作为采样点采集生物结皮样品(采样点概况见表1)。每个侵蚀类型区选取5个样地,样地间距大于1000 m。由于当地破碎的地形,一般面积大于300 m2的退耕地即可作为一个单独样地。每个样地布设4个5 m×5 m的大调查样方,调查高等植被群落组成、盖度。植被调查结束后,采用25点样方法调查样地生物结皮组成及盖度,每个大调查样方随机设10个小样方。之后每样地随机采集生物结皮样10个,每区采集生物结皮样品50个,三区共计150个样品。采集时用无菌铲尽可能地选择完好原状的结皮采集于直径9 cm无菌培养皿中,每次采样后用70%酒精擦拭采样工具,以防交叉污染。样品于采集地风干,快速运回实验室。
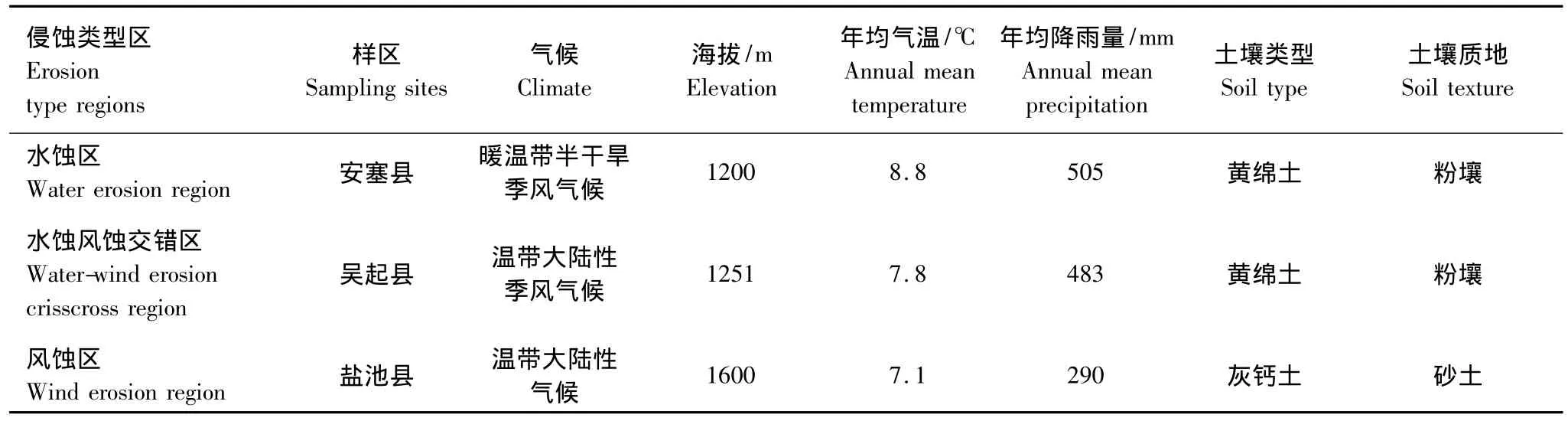
表1 采样区概况Table 1 General situation of the sampling sites
1.3 研究方法
1.3.1 蓝藻多样性
蓝藻多样性采用Shannon-Weiner指数表达:
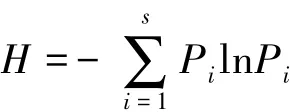
式中,S为物种数目,Pi为种i的个体在全体个体中的比例,H为物种的多样性指数[16]。
1.3.2 蓝藻种类组成
采用显微镜直接观察和室内培养箱培养观察相结合的方法。直接观察是取适量生物结皮样品,一部分放入试管内,用玻璃棒将其捣碎并轻微研磨,然后用无菌蒸馏水稀释,制成均一的土壤悬浊液,取一滴制作成临时装片在数码显微镜下观察并拍照,另一部分在适当水温条件下培养箱内培养2—3 d,取样研磨加水制成临时水装片,在显微镜下观察拍照,二者结果相结合得出最终镜检结果。培养观察采用BG-110蓝藻固体培养基和Bristol、BBM两种液体培养基。约10 g的样品,研磨过筛,一部分用无菌水稀释成不同浓度梯度的土壤悬液,接种于已灭菌的固体培养基;另一部分加入已灭菌的液体培养基,充分振荡,将接种后的固体培养基和液体培养基置于人工气候箱内进行培养,经过2—3周后进行观察并鉴定[17]。根据有关参考书[18-20]对蓝藻进行种类鉴定。
1.3.3 蓝藻优势种的确定
采用直接观察法。每个样品经研磨后,称取1 g土,加10 mL无菌蒸馏水,摇匀,取1滴均匀的土壤水溶液制成临时水装片,每个样品取3个临时装片,每个装片观察10个视野,对蓝藻进行拍照,统计不同种类并计数,最后根据出现频率的大小确定优势种[21]。
2 研究结果
2.1 黄土高原不同侵蚀类型区生物结皮中蓝藻的种类组成
经初步观察统计,黄土高原3个侵蚀类型区共发现蓝藻门植物4科10属54种,隶属于色球藻科(Chroococcaceae)、颤藻科(Oscillatoriaceae)、念珠藻科(Nostocaceae)、伪枝藻科(Scytonemataceae),色球藻属(Chroococcus)、集胞藻属(Synechocystis)、单歧藻属(Tolypothrix)、伪枝藻属(Scytonema)、鞘丝藻属(Lyngbya)、颤藻属(Oscillatoria)、席藻属(Phormidium)、微鞘藻属(Microcolus)、念珠藻属(Nostoc)、鱼腥藻属(Anabaena)(表2)。研究区蓝藻存在明显的科属现象,优势科为颤藻科,优势属为颤藻属。然而,色球藻科、念珠藻科和伪枝藻科三者之间并无明显差异。其中,以丝状蓝藻为主,占总数的87%。可见,蓝藻在黄土高原生物结皮中大都以群体形式存在,且丝状种类占绝对优势。
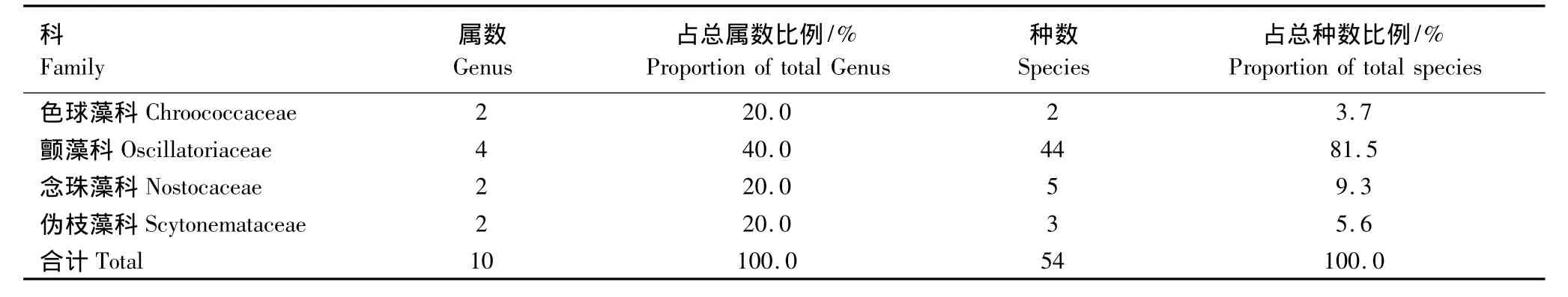
表2 黄土高原地区蓝藻科、属、种及其比例Table 2 The statistics of the number and proportion of Family,Genus,Species of cyanobacteria on the Loess Plateau region
2.2 不同侵蚀类型区蓝藻多样性、种类组成及优势种差异
2.2.1 蓝藻多样性特征
研究区蓝藻的Shannon-Weiner多样性指数表明,在黄土高原不同的侵蚀类型区生物结皮中,蓝藻的多样性水蚀风蚀交错区最高,水蚀区次之,风蚀区最低,多样性指数依次为2.22,2.20和2.14。图1是不同侵蚀区蓝藻多样性指数,由图1可见,水蚀风蚀交错区和水蚀区的蓝藻多样性指数之间差异不显著(P>0.05),但与风蚀区之间均达到显著性差异(P<0.05)。
黄土高原3个侵蚀类型区蓝藻的丰富度与多样性指数略有差异。图2是不同侵蚀类型区蓝藻的丰富度,由图2可见,3个侵蚀类型区的蓝藻丰富度差异显著(P<0.05),其中,水蚀风蚀交错区蓝藻丰富度最高,有39种,占黄土高原蓝藻总丰度的72.2%;水蚀区26种次之,风蚀区20种最少(图2)。
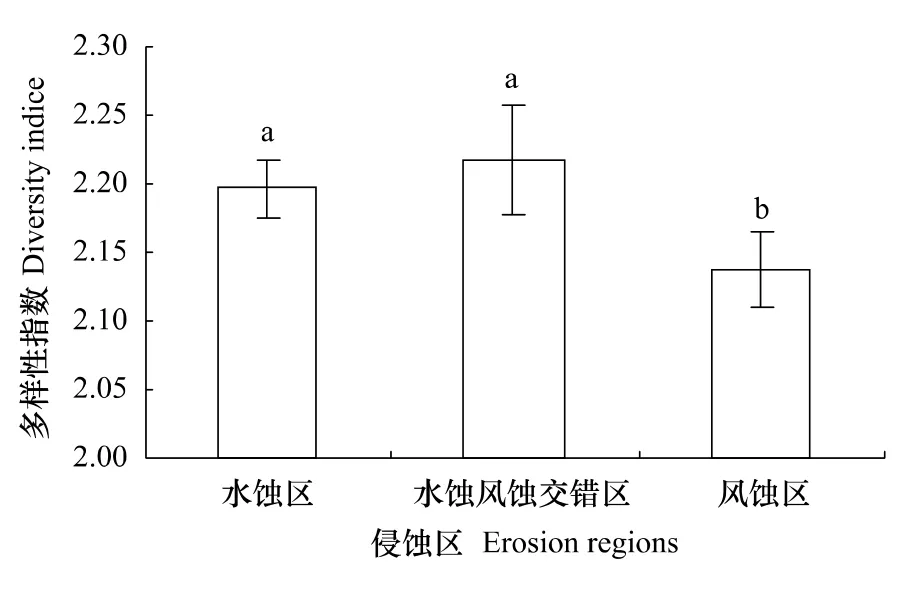
图1 不同侵蚀区蓝藻多样性指数Fig.1 Diversity indice in different erosion regions图中的不同字母表示处理间差异达到5%显著水平
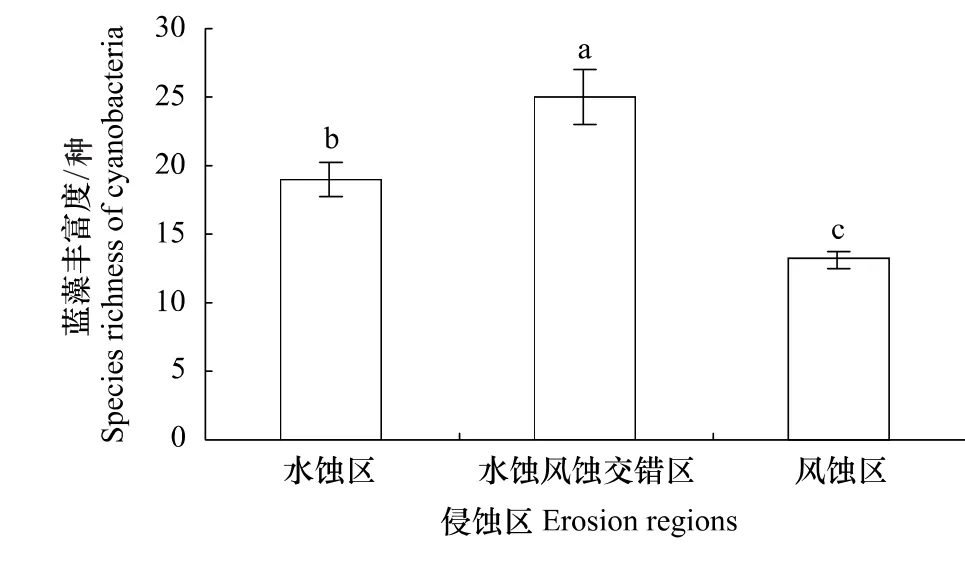
图2 不同侵蚀区蓝藻丰富度Fig.2 Species richness of cyanobacteria in different erosion regions图中的不同字母表示处理间差异达到5%显著水平
2.2.2 不同侵蚀类型区蓝藻优势种差异
3个侵蚀类型区之间不仅蓝藻多样性及丰富度有差异,优势种及种类组成也存在差异(表3)。水蚀风蚀交错区蓝藻优势种为阿氏鞘丝藻(Lyngbya allorgei),亚优势种为狭细席藻(Phormidium angustissimum);水蚀区为含钙席藻(Phormidium calciola),亚优势种为小席藻(Phomidium tenue);风蚀区为颗粒颤藻(Oscillatoria granulata),亚优势种为具鞘微鞘藻(Microcolus vaginatus)和沼地微鞘藻(Microcolus paludosus)。
3个区的优势种虽均属颤藻科,但却分属于不同的属,水蚀风蚀交错区的阿氏鞘丝藻属于鞘丝藻属,水蚀区的含钙席藻属于席藻属,风蚀区的颗粒颤藻属于颤藻属。可见,颤藻科蓝藻在黄土高原地区生物结皮发育的区域普遍存在,且种类繁多。
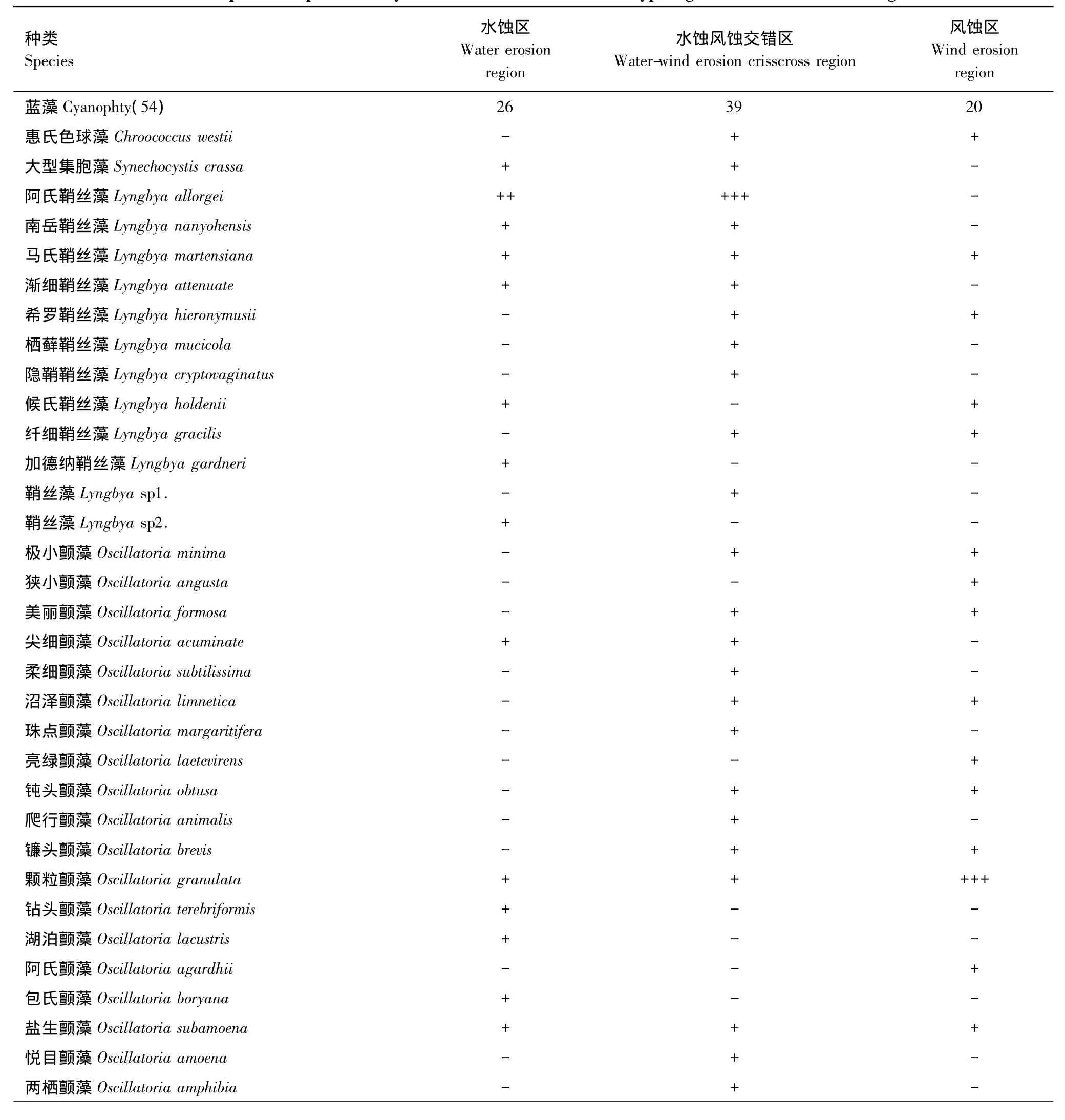
表3 不同侵蚀类型区生物结皮中蓝藻的种类组成Table 3 Species composition of cyanobacteria in different erosion type region in the Loess Plateau region
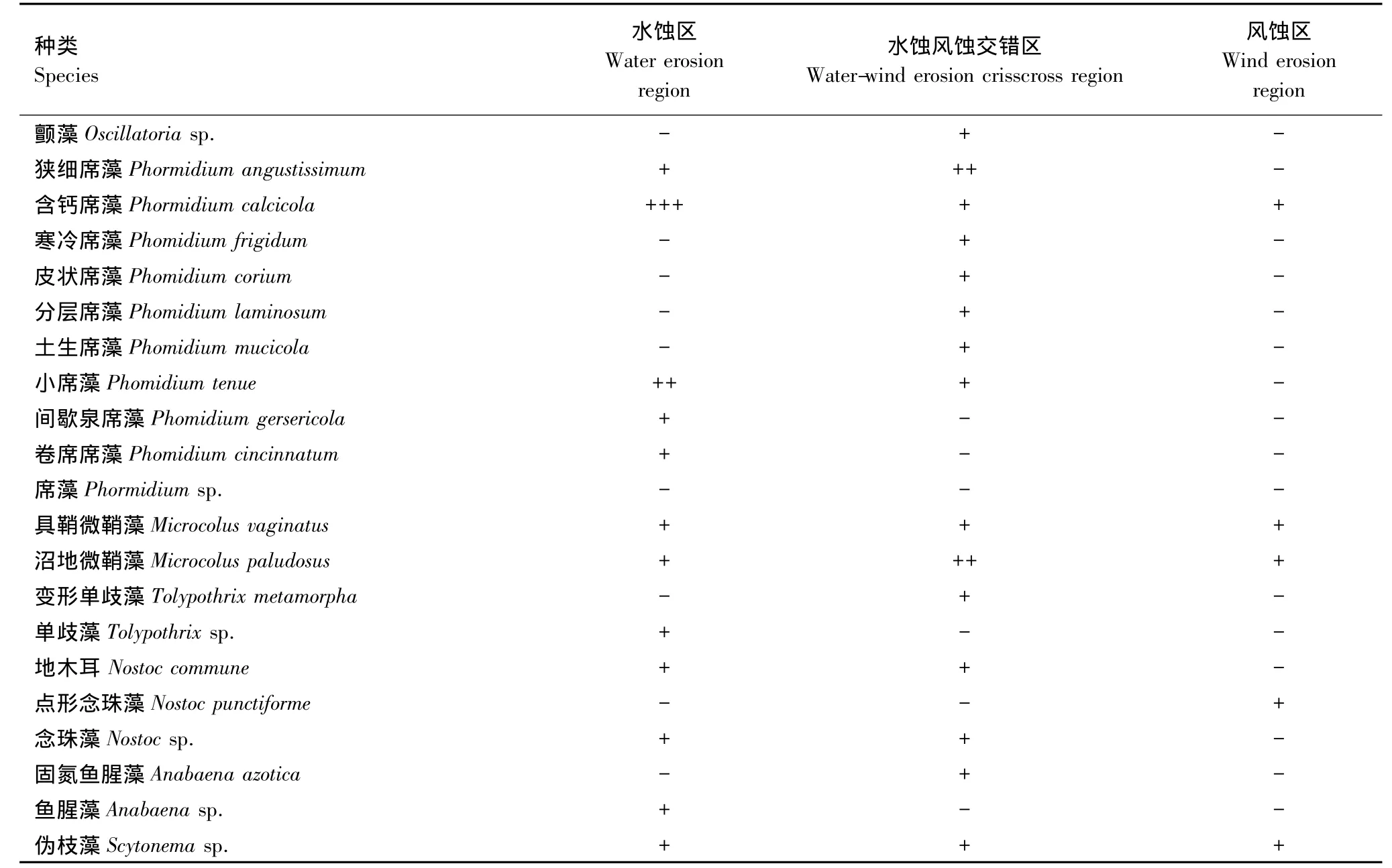
续表
2.3 蓝藻在黄土高原不同侵蚀类型区的选择性分布
从黄土高原3个侵蚀类型区的蓝藻分布看,有些种类的蓝藻对不同区域不具选择性,主要是颤藻科的种类,属于该地区的常见种。同时,还存在一些对区域具一定选择性的蓝藻,属该区特有种。不同侵蚀类型区蓝藻的特有种与常见种详见表4。
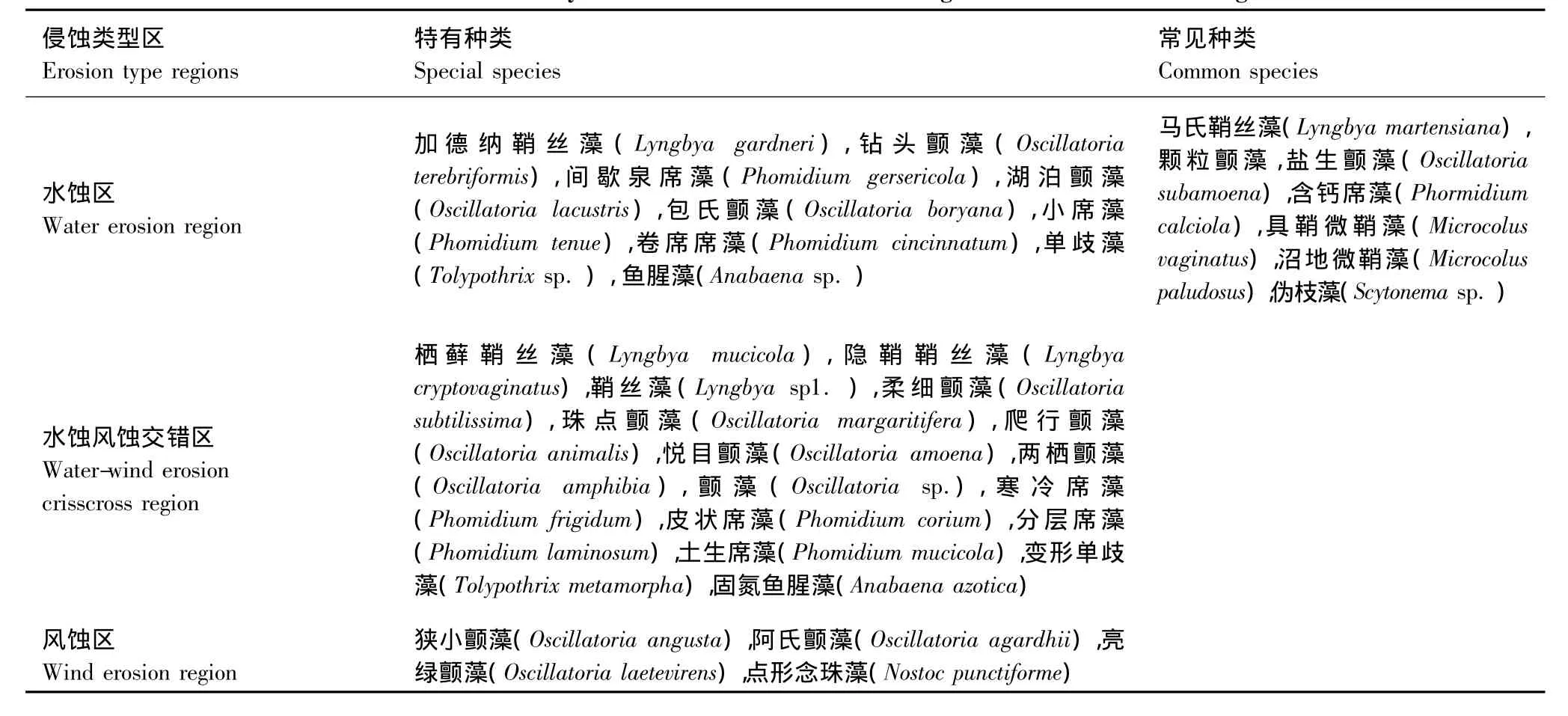
表4 蓝藻在黄土高原不同侵蚀类型区的分布Table 4 Distribution of cyanobacteria in different erosion region in the Loess Plateau region
3 分析与讨论
3.1 蓝藻在土壤生境中的适应性
蓝藻作为干旱半干旱地区土壤生境先锋植物中的主要类群,具有独特的形态结构、生理特性和生态适应性。在形态结构方面,蓝藻不仅细胞壁厚,细胞外被还有较厚的粘胶层或具色泽的胶鞘,这些结构都使蓝藻具有很强的保护性生理反应及生态适应性。当处于高温、干旱、强光辐射等不利环境时,能避免自身受到伤害[22],这可能是蓝藻在黄土高原地区分布广泛,种类繁多的原因。此外,一些丝状蓝藻(如颤藻科的很多种类)可以游动,使得它们可以避开强光干燥的环境以便处于更有利的环境[23],这可能是丝状蓝藻在黄土高原地区占绝对优势的原因。蓝藻的生态分布与土壤pH值、质地、温度、水分等多种因子相关,中性和碱性土壤有助于蓝藻的分布,质地较粘,颗粒较细的土壤上分布的蓝藻种类也较多[3]。在黄土高原地区,土壤pH值通常偏碱性(pH值一般为8.7左右),土壤颗粒较细,为蓝藻的广泛分布提供了良好的土壤生境,这可能是蓝藻在黄土高原地区普遍存在,且种类繁多的原因之一。
3.2 黄土高原不同侵蚀类型区与荒漠地区蓝藻多样性异同
在黄土高原地区,生物结皮中蓝藻存在明显的优势科属现象,优势科为颤藻科,优势属为颤藻属、鞘丝藻属、席藻属,其中又以丝状种类占优势,这与荒漠地区研究结果相一致[17,24]。胡春香研究发现兰州北山生物结皮中的蓝藻亦有较明显的优势科属现象,优势科是颤藻科和色球藻科,优势属都是颤藻属、席藻属和鞘丝藻属,都以丝状蓝藻占优势[25],与本文结果接近。张丙昌研究发现新疆古尔班通古特沙漠生物结皮中蓝藻也存在明显的优势科属现象,优势科是颤藻科和色球藻科,优势属是颤藻属、鞘丝藻属和色球藻属,色球科的球形种类相对较多[17],与黄土高原地区略有差异。这可能与蓝藻中球形种类具有更强的抗逆和固土性能有关[2,26],古尔班通古特沙漠地区太阳辐射强烈,风力威胁大,条件恶劣,与之相适应,只有这些抗逆性强的球形种类存活下来,成为了古沙漠的优势类群。而在黄土高原地区由于水分、温度、养分等条件较古沙漠优越,为丝状种类的生长、繁殖提供了适宜的条件,因此,球状种类较少,色球藻科不占优势。丝状种类占绝对优势这一特点与蓝藻的游动习性有关[23],为了与复杂的侵蚀环境相适应,具有强大缠绕、捆绑作用的丝状蓝藻最终适应了黄土高原三大侵蚀类型区脆弱的生态环境。此外蓝藻分泌的胞外多糖可以黏结、固定土粒,最终不仅产生了减少水蚀、风蚀的生态效应[17],同时为自身的生存改善了环境,进一步确定了蓝藻在黄土高原侵蚀类型区具有的重要水土保持效应。
3.3 不同侵蚀类型区生物结皮中蓝藻分布的差异
迄今为止,黄土高原地区生物结皮中蓝藻的研究少见报道。本文的结果初步表明,黄土高原地区三大侵蚀类型区生物结皮中均有多种蓝藻出现,且蓝藻多样性、种类组成与分布存在明显差异。水蚀风蚀交错区的蓝藻多样性、丰富度及球形种类和特有种都最多,水蚀区次之,风蚀区最少。这可能与研究区土壤pH值、质地、土壤理化性质等生境条件有关。张丙昌[27]在古尔班通古特沙漠的典型沙垄上研究发现藻类的种类组成受不同的地貌部位生境条件的差异、降水、气温和光照强度变化的影响,而物种多样性会进一步随之变化。黄土高原地区从水蚀区到水蚀风蚀交错区再到风蚀区,随着降雨量、温度的降低,植被逐渐稀疏,土壤质地由壤质到砂质,土壤pH值逐渐升高,3个侵蚀类型区自然条件之间的这些差异,这可能是影响蓝藻在3个侵蚀类型区分布差异的主要原因。此外,水蚀风蚀交错区蓝藻的特有种相对较多,一定程度上说明交错区生态环境特别是微生境相对多样化,与该区受水蚀、风蚀双重侵蚀作用,生态环境脆弱有着一定的关系。
综上所述,本文初步明确了黄土高原不同侵蚀类型区蓝藻种类组成和分布,要进一步明确该区生物结皮中蓝藻多样性特征仍需要从不同地貌、不同土层深度以及季节动态等方面对蓝藻种类组成及分布特征进行全面深入研究。
致谢:中国科学院安塞水土保持综合试验站对本研究给予支持,Jayne Belnap润色英文摘要,特此致谢。
[1] Belnap J,Lange O L.Biological Soil Crusts:Structure,Function,and Management.Berlin:Springer-Verlag,2003.
[2] Hu C X,Liu Y D,Song L R.New development of soil algae research.Acta Hydrobiologica Sinica,2002,26(5):521-528.
[3] Chen L Z,Liu Y D,Li D H,Shen Y W,Xie Z M.The research process of desert algae and crust.Science Foundation of China,2003,(2):90-93.
[4] Zhang B C,Zhang Y M,Zhao J C.Composition and ecological distribution of the algae living in the Gurbantunggut Desert of Xinjiang.Acta Botanica Boreali-Occidentalia Sinica,2005,25(10):2048-2055.
[5] Verrecchia E,Yair A,Kidron G J,Verrechia K.Physical properties of the psammophile cryptogamic crust and their consequences to the water regime of sandy soils,north-western Negev Desert,Israel.Journal of Arid Environments,1995,29(4):427-437.
[6] Hu C X,Zhang D L,Liu Y D.The newly progress of algae study of microbiotic crust in arid area.Progress in Natural Science,2003,13(8):791-795.
[7] Belnap J.Surface disturbances:their role in accelerating desertification.Environmental Monitoring and Assessment,1995,37(1/3):39-57.
[8] Zhao Y G,Xu M X,Wang QJ,Shao M A.Impact of biological soil crust on soil physical and chemical properties of rehabilitated grassland in hilly Loess Plateau,China.Journal of Natural Resources,2006,21(3):441-448.
[9] Xiao B,Zhao Y G,Shao M A.Artificial cultivation of biological soil crust and its effects on soil and water conservation in water-wind erosion crisscross region of Loess Plateau,China.Acta Agrestia Sinica,2008,16(1):28-33.
[10] Zhao Y G,Xu M X,Wang Q J,Shao M A.Physical and chemical properties of soil bio-crust on rehabilitated grassland in hilly Loess Plateau of China.Chinese Journal of Applied Ecology,2006,17(8):1429-1434.
[11] Xiao B,Zhao Y G,Shao M A.Effects of biological soil crust on saturated hydraulic conductivity in water-wind erosion crisscross region,North of Shaanxi Province,China.Transactions of the Chinese Society of Agricultural Engineering,2007,23(12):35-40.
[12] Zhao Y G,Xu M X,Belnap J.Response of biocrusts'photosynthesis to environmental factors:a possible explanation of the spatial distribution of biocrusts in the Hilly Loess Plateau region of China.Acta Ecologica Sinica,2010,30(17):4668-4675.
[13] The National Development and Reform Commission,Ministry of Water Resources,Ministry of Agriculture.The National Forestry Bureau.Planning brief about comprehensive treatments on the Loess Plateau.Macroeconomic Management,2011,(2):76-76.
[14] Tang K L.Importance and urgency of harnessing the interlocked area with both water and wind erosion in the Loess Plateau.Soil and Water Conservition in China,2000,(11):11-12,17-17.
[15] Tang K L,Hou Q C,Wang B K,Zhang P C.The environment background and administration way of wind-water erosion crisscross region and shenmu experimental area on the Loess Plateau.Memoir of Northwestern Institute of Soil and Water Conservation,Academia Sinica and Ministry of Water Resources,1993,(18):2-15.
[16] Sun R Y,Li Q F,Niu C J.Basic Ecology,Beijing:Higher Education Press,2002:143-144.
[17] Zhang B C,Zhang Y M,Zhao J C,Zhang R C.Study on cyanobacteria of biological soil crusts in Guerbantunggut Desert,Zhungar Basin,Sinkiang.Geography and Geo-Information Science,2005,21(5):107-109.
[18] Zhu H R.Flora Algarum Sinicarum Aquae Dulcis(TomusⅡ,Chroococcophyceae).Beijing:Science Press,1991:1-147.
[19] Zhu H R.Flora Algarum Sinicarum Aquae Dulcis(TomusⅨ,Cyanophyta).Beijing:Science Press,2007:1-312.
[20] Hu H J,Wei Y X.The Freshwater Algae of China—Systematics,Taxonomy and Ecology.Beijing:Science Press,2006:1-950.
[21] Zhang B C,Zhao J C,Zhang Y M,Li M,Zhang J.Vertical distribution of algae in different locations of sand dunes in the gurbantunggut desert,Xinjiang,China.Journal of Plant Ecology,2008,32(2):456-464.
[22] Hu C X,Liu Y D.Primary succession of algal community structure in desert soil.Acta Botanica Sinica,2003,45(8):917-924.
[23] Belnap J,Harper K T,Warren S D.Surface disturbance of cryptobiotic soil crusts:nitrogenase activity,chlorophyll content,and chlorophyll degradation.Arid Soil Research and Rehabilitation,1994,8(1):1-8.
[24] Hu C X,Liu Y D,Song L R.Species compositon and distribution of algae in shapotou area,Ningxia Hui autonomous region,China.Acta Hydrobiologica Sinica,1999,23(5):443-448.
[25] Hu C X,Zhang B C,Ma H Y,Liu Y D,Zhang D L.Species composition and community structure of terrestrial algae in the biological crusts of Lanzhou Northern Hill.Journal of Northwest Normal University:Natural Science Edition,2003,39(1):59-63.
[26] Hu C X,Liu Y D,Song L R,Zhang D L.Effect of desert soil algae on the stabilization of fine sands.Journal of Applied Phycology,2002,14(4):281-292.
[27] Zhang B C,Zhang Y M,Wang J Z.Spatio-temporal distribution of algae relating to biological soil crusts in the Gurbantunggut Desert,Xinjiang,China.Journal of Desert Research,2011,31(4):919-926.
参考文献:
[2] 胡春香,刘永定,宋立荣.土壤藻研究新进展.水生生物学报,2002,26(5):521-528.
[3] 陈兰周,刘永定,李敦海,沈银武,谢作明.荒漠藻类及其结皮的研究.中国科学基金,2003,2:90-93.
[4] 张丙昌,张元明,赵建成.古尔班通古特沙漠生物结皮藻类的组成和生态分布研究.西北植物学报,2005,25(10):2048-2055.
[6] 胡春香,张德禄,刘永定.干旱区微小生物结皮中藻类研究的新进展.自然科学进展,2003,13(8):791-795.
[8] 赵允格,许明祥,王全九,邵明安.黄土丘陵区退耕地生物结皮对土壤理化性状的影响.自然资源学报,2006,21(3):441-448.
[9] 肖波,赵允格,邵明安.黄土高原侵蚀区生物结皮的人工培育及其水土保持效应.草地学报,2008,16(1):28-33.
[10] 赵允格,许明祥,王全九,邵明安.黄土丘陵区退耕地生物结皮理化性状初报.应用生态学报,2006,17(8):1429-1434.
[11] 肖波,赵允格,邵明安.陕北水蚀风蚀交错区两种生物结皮对土壤饱和导水率的影响.农业工程学报,2007,23(12):35-40.
[12] 赵允格,许明祥,Belnap J.生物结皮光合作用对光温水的响应及其对结皮空间分布格局的解译——以黄土丘陵区为例.生态学报,2010,30(17):4668-4675.
[13] 国家发展改革委,水利部,农业部,国家林业局.国家发展改革委等部门印发《黄土高原地区综合治理规划大纲(2010—2030年)》.宏观经济管理,2011,(2):76-76.
[14] 唐克丽.黄土高原水蚀风蚀交错区治理的重要性与紧迫性.中国水土保持,2000,(11):11-12,17-17.
[15] 唐克丽,侯庆春,王斌科,张平仓.黄土高原水蚀风蚀交错带和神木试区的环境背景及整治方向.中国科学院水利部西北水土保持研究所集刊(神木水蚀风蚀交错带生态环境整治技术及试验示范研究论文集),1993,(18):2-15.
[16] 孙儒泳,李庆芬,牛翠娟.基础生态学.北京:高等教育出版社,2002:143-144.
[17] 张丙昌,张元明,赵建成,张茹春.准噶尔盆地古尔班通古特沙漠生物结皮蓝藻研究.地理与地理信息科学,2005,21(5):107-109.
[18] 朱浩然.中国淡水藻志.第二卷,色球藻纲.北京:科学出版社,1991:1-147.
[19] 朱浩然.中国淡水藻志.第九卷,蓝藻门,藻殖段纲.北京:科学出版社,2007:1-312.
[20] 胡鸿钧,魏印心.中国淡水藻类——分类、区系与生态.北京:科学出版社,2006:1-950.
[21] 张丙昌,赵建成,张元明,李敏,张静.新疆古尔班通古特沙漠南部沙垄不同部位藻类的垂直分布特征.植物生态学报,2008,32(2):456-464.
[24] 胡春香,刘永定,宋立荣.宁夏沙坡头地区藻类及其分布.水生生物学报,1999,23(5):443-448.
[25] 胡春香,张斌才,马红樱,刘永定,张德禄.兰州北山生物结皮中陆生藻种类组成与群落结构.西北师范大学学报;自然科学版,2003,39(1):59-63.
[27] 张丙昌,张元明,王敬竹.古尔班通古特沙漠南缘典型沙垄藻类的时空分布.中国沙漠,2011,31(4):919-926.
猜你喜欢
农业工程学报(2022年5期)2022-06-22 10:22:40
农技服务(2021年10期)2021-12-02 08:53:35
中国房地产业·上旬(2019年12期)2019-03-24 09:44:41
烟台果树(2019年1期)2019-01-28 09:34:46
现代园艺(2018年3期)2018-02-10 05:18:21
现代农业科技(2014年11期)2014-09-24 11:08:11
水土保持研究(2014年2期)2014-05-07 11:07:32
黑龙江水利科技(2012年3期)2012-08-15 00:49:58
环境影响评价(2012年1期)2012-01-31 08:36:10
自然资源遥感(2012年1期)2012-01-05 07:57:30
