
梭梭土壤种子库空间分布的尺度依赖性研究
2013-09-14 09:19陈云龙宋于洋周朝彬
水土保持研究 2013年4期
陈云龙,宋于洋,周朝彬
(1.石河子大学 农学院,新疆 石河子832003;2.农九师林业工作管理站,新疆 塔城834601)
空间格局表现出的方向和尺度依赖性已成为现代生态学家的共识[1-3]。对空间格局进行各向异性和多尺度研究有助于更细致地分析空间分布格局及其与环境之间的关系[4],解决空间格局的尺度依赖问题并获得直观的结果[5]。地统计学中变异函数与方向、抽样尺度有关,因此可用于分析空间异质性的各向异性和尺度依赖性。除此之外,小波分析及其改进后的角小波分析(angular wavelet analysis)、扩展最近邻体分析法(extended nearest neighbor analysis,ENNA)等方法也常用于空间格局的各向异性和尺度依赖性分析[6-7]。近年来,国内外学者对空间格局的各向异性和尺度依赖性进行了有益的探讨[8-10],但总体来看还处于初步探索阶段。
土壤种子库是指存在于土壤上层凋落物和土壤中全部存活种子的总和[11]。受母树、地被覆盖物等各种因素的影响,在方向和尺度的共同作用下,种子库具有明显的各向异性和尺度依赖性[10,12]。林分中土壤种子库是林木更新过程的一个重要阶段,是林分动态中主要的生态学过程之一[13]。已有研究表明,种子在土壤中的空间分布具有高度异质性[14]。种子库的空间格局及其生态学过程对苗木的更新格局有决定性的影响[15-16]。因此,研究种子库格局特征及其受控因素对认识林木更新机制有重要意义。
近年来国内外学者进行了很多关于荒漠生态系统种子库的研究[17-23],这些研究在很大程度上是定性或概括性描述。对于土壤种子库空间分布的各向异性和尺度依赖性研究仍很缺乏[10]。有关古尔班通古特沙漠不同地区梭梭土壤种子库空间分布的各向异性和尺度依赖性仍未见报道。
梭梭(Haloxyl on Ammodendr on)为藜科,梭梭属植物,具有很强的抗干旱、耐盐碱能力,是气候干旱、盐碱土广泛分布的新疆荒漠区的优势种群,对维护当地生境的稳定具有重要的生态价值[24]。本文采用地统计学理论与方法,对梭梭土壤种子库空间分布格局进行各向异性和尺度依赖性的研究,旨在为古尔班通古特沙漠梭梭种子库分布格局与梭梭种群更新及恢复提供一定的科学依据。
1 实验材料与方法
1.1 样地设置
于2012年3月初在古尔班通古特沙漠南缘选择典型的沙丘作为研究地点,采用相邻网格法,在精河(44°52′N,83°35′E)、奎屯(44°34′N,84°77′E)和五家渠(44°39′N,87°34′E)迎风坡分别设置3个样地(表1)。各样地采用GPS定位(样地采用经纬仪校订均为正南正北方向)、每个样地面积为0.25 h m2(50 m×50 m)。调查时首先将50 m×50 m的样地等距离间隔划分为100个5 m×5 m小样方,先对小样方进行编号,然后在每个小样方中按对角线选择3个0.2 m×0.2 m取样点,对土壤分0—1,1—2 c m和2—3 c m 3层进行取样,再将同土层的三个土样进行混合,共采集土样900个,带回实验室进行种子萌发实验。

表1 所选样地的环境因子及植被状况
1.2 种子萌发
将带回实验室的土样去除枯枝烂根等杂物,将各层土样分别充分混匀后平铺于直径25 c m,高20 c m的花盆内,花盆底部沙层厚度2~3 c m,置于玻璃温室(28℃/20℃)中进行发芽试验,萌发期间定期喷洒适量蒸馏水,使土壤保持湿润状态。每天早晨10:00观察记录梭梭种子萌发情况,将新萌发出土的幼苗移除,辨别鉴定后,统计梭梭幼苗数量。萌发实验从3

式中:Z(xi),Z(xi+h)——区域化随机变量Z在空间位置xi和xi+h上的取值;N(h)——抽样间隔等于h时的点对数;γ(h)——变异函数值(又称为半方差,semivariance)。以半方差γ(h)为Y轴,以抽样间隔h为 X轴,用SPSS 17.0软件绘制散点图[27]。
1.3.2 最优模型拟合 通过半方差公式计算半变异函数值,分别用不同类型的理论模型拟合,得到模型的相关参数。本文采用平均预测误差(ME)越小越好、标准平均值(MS)最接近于0、均方根预测误差(RMSE)最小、平均预测标准差(ASE)最接近于均方根预测误差、均方根标准预测误差(RMSS)越接近于1越好、平均绝对误差(MAE)越小越好的原则评估不同模型的插值效果。本研究中得到的最优理论模型均为球状模型。模型表达式为:月22日开始,至3月3月28日发芽结束[25]。
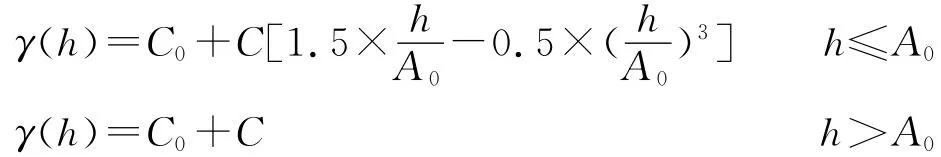
1.3 数据处理
使用SPSS 17.0软件分析不同样地种子平均密度的差异性。
种子库的空间格局应用地统计学方法进行分析[26],在 Arc GIS 9.3中以Geostatisical Anal yst地统计分析模块分析各取样点种子库统计特征、半方差模型,进行空间插值和交互检验。
1.3.1 半方差函数 采用半方差分析(semivariance analysis)的方法研究种子库的土壤空间异质性,半方差函数表达式为:
式中:h——分隔空间两点的矢量,又称滞后距;C0——块金值(不小于0);C——偏基台值(不小于C0);A0——变程,球状模型中A0等于有效变程a。
1.3.3 空间局部插值 本研究采用地统计学中应用极为广泛的普通Kriging法(Ordinary Kriging)进行插值。设一区域内Z(x0)为某一变量在位置x0处的估值,其周围相关范围内的已测定值为Z(xi)(i=1,2,3,…,n),则x0处的估值可用线性组合来估测:
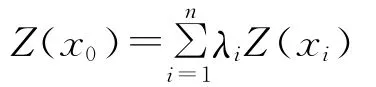
式中:λi——与Z(xi)位置有关的权重系数,表示各测定值对估计值Z(x0)的贡献,其和为1。
2 结果与分析
2.1 种子库统计特征
3个样地梭梭土壤种子库平均密度有显著差异(表2,P<0.01)。奎屯样地种子贮量最高,平均密度达152.58粒/100 m2,分别是五家渠和精河样地的1.75倍和9.51倍。变异系数分析表明,精河样地变异性较小,而奎屯样地变异性较大。

表2 三个样地种子库密度统计特征
2.2 不同尺度下土壤种子库空间结构分析
3个样地梭梭土壤种子库的空间格局表现出明显的尺度依赖性。比较不同尺度上变异函数的理论模型发现,随着观察尺度的增大,3个样地块金值呈现出增大趋势,表明随机因素对种子库空间格局的影响有增大趋势(表3)。当最小抽样尺度分别为5,10,15 m时,奎屯和五家渠样地基台值随最小抽样尺度的增加而呈增加趋势,表明空间异质性程度在提高。奎屯和五家渠样地在最小抽样尺度为20 m时,基台值出现拐点,意味着可能存在明显的不同尺度范围的作用机制,对种子库空间格局产生重要的影响。精河样地的基台值随抽样尺度的变化基本保持不变。
当最小抽样尺度分别为5,10,15,20,25 m 时,奎屯和五家渠样地种子库在5~88.4 m的空间尺度上仍可观察到强烈的空间格局,格局强度的空间自相关变异解释程度分别为46%~50%和81%~85%。精河在抽样尺度从5~25 m范围内变化时,仅观察到中等程度的空间格局,格局强度的31%~34%可由空间自相关变异来解释。
2.3 各向异性下土壤种子库的空间结构分析
对于区域化变量,变异函数不仅与间隔距离有很强的相关性,而且也与方向性有关。当一个变异函数是由某一个特殊方向构造时,称为各向异性变异函数[26]。
各向异性下各土壤种子库空间变异的结构参数见表4。奎屯样地45°和90°方向上块金值比另外两个方向的大,说明梭梭种子库在45°和90°方向上变化的结构性要弱于另外两个方向。较大的块金值表明小尺度上的生态学过程不可忽视,较大的块金值是源于微尺度上的结构变异还是试验误差,尚需进一步缩小取样尺度进行深入研究[28]。五家渠样地0°和90°方向上变化的结构性要弱于另外两个方向,精河样地45°和135°方向上变化的结构性要弱于另外两个方向。比较基台值发现,同一样地不同方向上基台值差异不大,表明各方向上空间异质化程度相似。

表3 3个样地不同抽样尺度下半方差函数的参数

表4 3个样地各向异性下半方差函数的参数
同一样地不同角度之间变程存在差异,某个方向上的变程越长,则该方向上有较强的空间连续性[29]。奎屯样地种子库45°,90°方向上变程大于0°,135°方向,五家渠样地种子库在0°,90°和135°方向上变程大于45°方向,精河样地在0°和90°方向上变程大于另外两个方向。表明了3个样地不同方向上具有空间连续性差异。精河样地在45°和135°方向上、奎屯样地在0°和135°方向上、五家渠样地在0°和90°方向上的空间结构比显著小于另外两个方向,表明由空间自相关部分引起的空间异质性较低。
2.4 克里格插值结果
在土壤种子库变异函数理论结构分析的基础上,采用克里格法对采样区进行最优化内插,应用Arc-GIS 9.3绘制各样地空间格局图(图1)。各样地种子库密度梯度变化均较清晰,种子呈明显聚集分布。精河样地的种子密度低,且分布较均匀,这与样地较平整且精河风大有密切关系。奎屯样地克里格插值图中低密度斑块镶嵌在高密度斑块中,可能和该样地有一部分沙丘上无植被有关。五家渠样地中高密度斑块镶嵌在低密度斑块中,分布均匀斑块边界变化较简单。调查发现,奎屯和五家渠样地中种子高密度斑块处短命植物层发育良好,可有效降低风速,有利于种子停留,表明地被覆盖物显著影响着种子库的空间分布。

图1 各样地种子库空间分布格局
3 讨论
3.1 梭梭种子库空间分布的各向异性
在地统计学中,表示研究对象的空间异质性的基台值是系统或系统属性中最大的变异,由自相关变异和随机变异组成[26]。其中,结构性因素引起自相关变异,随机因素引起随机变异。通常,影响土壤种子库空间分布的结构性因素由母树空间分布、结实能力、比例、土壤微环境、林下杂草和枯落物等组成[30-32],随机因素由风吹、流水、动物摄食和搬运等组成[33-35]。已有研究表明,古尔班通古特沙漠的成年梭梭母树空间位置呈随机分布[36],表明种子源的空间分布是随机的。我们的研究结果显示,梭梭土壤种子库分布格局与母树分布完全不同。3个样地梭梭土壤种子库在不同方向上的空间结构比在0.3~0.88之间,具有中等以上的空间结构变异且各向异性特征明显;空间插值图表明种子库梯度变化大,呈明显聚集分布。这可能是因为梭梭种子离开母树后,在传播过程中受到地表障碍物空间异质性分布的影响而进行了重新分布[32],而地表障碍物如短命植物层受地形、土壤理化性质等的影响,具有较高的空间异质性[37]。在古尔班通古特沙漠中,草本层一般在4—6月份完成其生活史,至11月梭梭种子成熟时,草本植物仅留下部分干枯地上部分。草本植物种类不同导致其地上部分构型有差异,对种子库形成的影响不同。另外,同种草本植物盖度越大,捕获梭梭种子能力越强。3个样地中,精河、五家渠和奎屯的草本植物层盖度依次降低(表1)。另外,精河降水较另两个样地少(表1),短命植物种类最少。奎屯降水丰富,物种数量高于五家渠,丰富的物种和高的盖度营造了异质性强的生境。因此,奎屯种子库空间结构比最高,而精河最低。
梭梭种子具果翅,在离开母树后其空间格局重新分布必然会受到风的影响。Keitt认为,种子传播在盛行风向具有较强的方向依赖性,即具有各向异性特征[38]。古尔班通古特沙漠平均风速在2.0~3.7 m/s间(表1),本研究中选择的3个样地主风向为西北风,也时常出现东风或东北风。研究发现,由于土壤种子库空间分布受母树位置和结实能力、风吹、短命植物层空间分布等的综合影响,梭梭土壤种子库的空间异质性均没有表现出风向的一致性,即在西北和东北方向上没有表现出一致的差异性,各个方向上的空间结构表现出无序特征。风吹作用仅是梭梭种子离开母树后格局发生重新分布的影响因子之一。
3.2 梭梭种子库空间分布的尺度依赖性
生态学中空间分布格局是空间异质性的具体表现,而异质性是尺度的函数,因此,空间格局对尺度有很强的依赖性。从不同抽样尺度上获得信息能够了解研究对象的细节[39]。
从不同的观察尺度上进行深入研究,更容易发现一些潜在的空间结构特征。韩有志和王政权[10]通过分析分维值发现,种子库的空间格局在不同尺度上具有某种等级结构特征。我们的研究发现,奎屯和五家渠样地块金值、基台值随观察尺度的增加均呈增大趋势,表明随尺度的增加,种子库空间异质性呈增加趋势。这是因为奎屯和五家渠样地梭梭种子空间分布,在小尺度上受母树结实能力和地表短命植物层异质性分布的影响,在大尺度上则还可能受到地形、母树的随机分布的影响。大小不同尺度的相互作用机制,加强了种子库分布的空间异质性。因此,随尺度的增加,空间异质性加大。精河样地所处环境水分条件恶劣,植被稀疏,风速大,种子库的空间分布受风力影响,导致5~25 m尺度上空间异质性差异不大。
[1] 郭旭东,傅伯杰,陈利顶,等.河北省遵化平原土壤养分的时空变异特征:变异函数与Kriging插值分析[J].地理学报,2000,11(4):557-563.
[2] Wu J,Shen W,Sun W,et al.Empirical patter ns of the effects of changing scale on landscape metrics[J].Landscape Ecology,2002,17(8):761-782.
[3] 王本洋,余世孝,王永繁,等.种群分布格局的各向异性分析[J].中山大学学报,2006,45(2):83-87.
[4] Lichsten J W,Si mons T R,Shriner S A,et al.Spatial autocorrelation and autoregressive models in ecology[J].Ecological Monographs,2002,72(3):445-463.
[5] 马克明,祖元刚.植被格局的分形特征[J].植物生态学报,2000,24(1):111-117.
[6] Rosenberg M S.Wavelet analysis for detecting anisotropy in point patter ns[J].Jour nal of Vegetation Science,2009,15(2):277-284.
[7] 王本洋,余世孝.种群分布格局的多尺度分析[J].植物生态学报,2005,29(2):235-241.
[8] 郭旭东,傅博杰,马克明,等.基于GIS和地统计学的土壤养分空间变异特征研究:以河北省遵化市为例[J].应用生态学报,2000,11(4):557-563.
[9] 路鹏,彭佩钦,宋变兰,等.洞庭湖平原区土壤全磷含量地统计学和 GIS分析[J].中国农业科学,2005,38(6):1204-1212.
[10] 韩有志,王政权.两个林分水曲柳土壤种子库空间格局的定量比较[J].应用生态学报,2003,14(4):487-492.
[11] Si mpson R L.Ecology of Soil Seed Bank[M].San Diego:Academic Press,1989.
[12] Matlack G R,Good R E.Spatial heterogeneity in the soil seed bank of a mat ure Coastal Plain forest[J].The Jour nal of the Torrey Botanical Society,1990,117(2):143-152.
[13] Nathan R,Safriel U N,Noy-Meir I,et al.Spatiotemporal variation in seed dispersal and recr uit ment near and far from Pinus halepensis trees[J].Ecology,2000,81(8):2156-2169.
[14] Thompson K,Bakker J P,Bekker R M,et al.Ecological correlates of seed persistence in soil in the northwest European flora[J].Jour nal of Ecology,2002,86(1):163-169.
[15] Oakley B B,Franklin J J.Bitter cherry(Pr unus emarginata)distribution,successional dynamics,and implications for the role of the seed bank[J].Canadian Journal Botany,1998,76(10):1725-1732.
[16] Bekker R M,Ver weij G L,Bakker J P,et al.Soil seed bank dynamics in hayfield succession[J].Jour nal of E-cology,2000,88(4):594-607.
[17] 郑翠玲,曹子龙,赵廷宁,等.围封沙化草地土壤种子库动态研究[J].水土保持研究,2005,12(6):169-171.
[18] 刘华,蒋齐,王占军,等.不同封育年限宁夏荒漠草原土壤种子库研究[J].水土保持研究,2011,18(5):96-98.
[19] 曾彦军,王彦荣,南志标,等.阿拉善干旱荒漠区不同植被类型土壤种子库研究[J].应用生态学报,2003,14(9):1457-1463.
[20] 张涛,田长彦,孙羽,等.古尔班通古特沙漠地区短命植物土壤种子库研究[J].干旱区地理,2006,29(5):675-681.
[21] 苏延桂,李新荣,陈应武,等.生物土壤结皮对荒漠土壤种子库和种子萌发的影响[J].生态学报,2007,27(3):938-946.
[22] 仝川,冯秀,仲延凯.内蒙古锡林郭勒克氏针茅退化草原土壤种子库特征[J].生态学报,2009,29(9):4710-4719.
[23] 吴涛,王雪芹,盖世广,等.春夏季放牧对古尔班通古特沙漠南部土壤种子库和地上植被的影响[J].中国沙漠,2009,29(3):499-507.
[24] 黄培祐,李启剑,袁勤芬.准噶尔盆地南缘梭梭群落对气候变化的响应[J].生态学报,2008,28(12):6051-6059.
[25] 王习勇,魏岩,严成,等.温度和盐分对两种梭梭种子萌发的影响[J].干旱区研究,2004,21(S):58-63.
[26] 李哈滨,王政权,王庆成.空间异质性定量研究理论与方法[J].应用生态学报,1998,9(6):651-657.
[27] 冯益明,唐守正,李增元.空间统计分析在林业中的应用[J].林业科学,2004,40(3):149-155.
[28] 王存国,韩士杰,张军辉,等.长白山阔叶红松林表层土壤水分空间异质性的地统计学分析[J].应用生态学报,2010,21(4):849-855.
[29] 余昊,王登元,王运兵,等.春尺蠖种群空间格局的地质统计学分析[J].云南农业大学学报,2006,21(3):303-306.
[30] Matlack G R,Good R E.Spatial heterogeneity in the soil seed bank of amature Coastal Plain f orest[J].The Jour nal of the Torrey Botanical Society,1990,117(2):143-152.
[31] Minore D,Weatherly H G,Cart mill M.Seeds,seedlings,and growth of Pacific yew (Taxus brevif olia)[J].North west Sci.,1996,70(3):223-229.
[32] Russell S K,Schupp E W.Effects of microhabital patchiness on patter n of seed dispersal and seed predation of Cercocar pus ledif olius(Rosaceae)[J].Oikos,1998,81(3):434-443.
[33] Prince M V,Joyner J W.What resources are available to desert granivores:seed rain or soil seed bank[J].E-cology,78(3):764-773.
[34] Nystrand O,Granstrom A.Predation on Pinus syl vestris seeds and juvenile seedlings in Swedish boreal forest in relation to stand dist ur bance by logging[J].Appl.Ecol.,2000,37(3):449-463.
[35] Wenny D G.Seed dispersal,seed predation,and seedling recr uit ment of a Neotropical montane tree[J].Ecol.Monogr.,2000,70(2):331-351.
[36] 宋于洋,李园园,张文辉.梭梭种群不同发育阶段的空间格局与关联性分析[J].生态学报,2010,30(16):4317-4327.
[37] 钱亦兵,吴兆宁,张立运,等.古尔班通古特沙漠植被与环境的关系[J].生态学报,2007,27(7):2802-2811.
[38] Keitt T H.Spectral representation of neutral landscape[J].Landscape Ecology,2000,15(5):479-493.
[39] 肖笃宁,布仁仓,李秀珍.生态空间理论和景观异质性[J].生态学报,1997,17(5):453-461.
猜你喜欢
林业资源管理(2022年6期)2023-01-18
哈哈画报(2022年4期)2022-04-19
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
科学大众(2020年17期)2020-10-27
中学生天地(B版)(2019年4期)2019-05-08
中国棉花加工(2017年2期)2017-06-09
大陆桥视野·上(2016年4期)2016-11-19
大陆桥视野(2016年7期)2016-06-13
