
安徽新安江干流滩涂湿地草本植物区系及物种多样性
2013-09-11 08:38杨文斌周守标
生态学报 2013年5期
杨文斌,刘 坤,周守标
(1.安徽师范大学环境科学与工程学院,芜湖 241003;2.安徽师范大学生命科学学院,芜湖 241000)
湿地与森林、海洋一起并称为全球三大生态系统,它广泛分布于世界各地,是地球上最富生物多样性的生态景观和人类最重要的生存环境之一[1-2]。湿地位于水陆交界处,是在水陆交互作用下形成的独特的生态系统,在保护生物多样性、涵养水源、净化水体、蓄洪防洪、提供休闲旅游场所等方面起着极其重要的作用[3]。湿地植物作为湿地生态系统食物链中的生产者,影响着湿地生态系统的生存和发展,也在一定程度上决定着湿地动物的种类和数量。分析湿地草本植物的区系组成及其物种多样性对湿地保护、特别是水资源的保护和湿地动物多样性保护和研究具有重要的意义,也可为受损湿地植被恢复提供可靠的依据[4]。滩涂湿地是平原河床季节性淹水的微地形,因濒临的水体不同,可分为内陆湖泊滩涂湿地、沿河(江)滩涂湿地及沿海滩涂湿地等[5]。河流滩涂湿地作为一种沿江滩涂湿地,也可归类为河流湿地中的洪泛平原湿地。虽然目前国内有关滩涂湿地的研究较多,但对于新安江干流滩涂湿地植被的相关研究很少,有关植物区系的研究更未见报道。新安江干流为千岛湖的重要水源补给河流,也是我国首个跨省流域水环境补偿试点流域。本文通过对新安江干流(安徽段)草本植物区系及其物种多样性等方面的研究,旨在为本区域湿地植物资源的保护、合理利用和生态管理以及湿地植被恢复等提供相关的科学依据。
1 研究区概况
新安江干流安徽段位于皖境最南端,属钱塘江水系,居流域的上游。新安江流域位于29°25'—30°16'N、117°38'—118°56'E,东南以天目山脉、白际山脉为界与浙江省毗邻,西南与江西省鄱阳湖水系接壤,北以长江、新安江分水岭为界。新安江干流跨皖浙两省,发源于安徽黄山市休宁县海拔1630 m的六股尖,总长约230 km,流域面积6456 km2。新安江自西向东流经安徽省黄山市的休宁县、屯溪区和歙县,在歙县的街口镇进入浙江省后入淳安、建德市的千岛湖(即新安江水库)。新安江干流安徽段湿地所在区域的气候类型为亚热带湿润季风气候,气候温暖,四季分明,冬无严寒,夏无酷暑,雨量充沛,湿度较大。年平均气温15.6℃,变化范围在15.0—16.2℃之间,极端最高温度39.5℃,极端最低-12.2℃,≥0℃年均积温为8756℃,≥10℃年均积温7125℃;年平均降水量约1780 mm,变化范围1636—1910 mm之间,雨量集中在4—7月;年均蒸发量约1260 mm,变化范围在1122—1389 mm之间;相对湿度较高,无霜期220 d左右,地域性小气候突出,春夏期间多云多雾、多雨。湿地区四周山体岩石主要由花岗岩组成,并兼有千枚岩、板岩、砂砾岩、石灰岩块等,山地土壤主要为黄粽壤、红壤和黄壤。
2 研究方法
2.1 样地设置与调查方法
2011年7—8月,在滩涂湿地植物最佳生长季节,对新安江干流安徽段滩涂湿地植物进行野外调查。由于滩涂湿地上偶有乔木和灌木,因此,本文只对江段滩涂湿地草本植物进行调查研究。依据不同干扰类型和环境梯度,分别在江段上、中、下游共选取8个滩涂湿地(即8个样地)进行调查研究,研究区域及样地设置见图1,各样地基本状况见表1,群落类型采取优势种命名的原则。参照一些学者对滩涂湿地调查取样的方法[2,6],根据滩地湿地具体环境和岸线情况,在每个样地靠近河岸线的草本群落内平行于岸线设置3条样线,在每条样线上间隔一定距离设置10个1 m×1 m的样方,共240个样方,调查内容包括:草本植物的种类、数量、盖度、多度和平均高度等,同时测定记录各样地的生境因子(包括地势、土壤类型等)。

图1 研究区域及样地设置示意图Fig.1 The diagram of study area and plots
2.2 区系分析
植物分类、鉴定及生态类型划分以《安徽植植物志》、《中国湿地植被》为标准[7-8]。区系成分分析以《中国种子植物属的分布区类型》和《中国种子植物区系统计分析》为依据[9-10],参照潘云芬等[4]和金孝锋等[11]的植物区系研究方法进行分析。在进行科属种的区系分析时,种下分类群均视作种级看待。

表1 各样地基本概况Table 1 Characteristics of the eight plots
2.3 统计分析
参考方精云等[12]、娄彦景等[13]和王立龙等[2]的物种多样性的测度方法,对样地草本植物多样性的测定采用以下指数和方法:
(1)重要值Ⅳ

(2)物种丰富度

(3)α多样性测定方法

式中,Pi为第i种的重要值;S为样地内出现的物种数。
(4)β多样性测定方法

式中,a和b分别为两群落的物种数,c为两群落的共有物种数。
3 结果与分析
3.1 滩涂湿地草本植物区系的组成
根据本次群落调查和初步统计,安徽新安江流域滩涂湿地共有草本被子植物48科131属194种(含种下分类单位,下同),其中双子叶植物37科88属129种,单子叶植物11科43属65种。另外,调查中有蕨类植物4科4属5种,不在本文中予以讨论。
3.1.1 滩涂湿地草本植物科的分析
(1)科的大小分析
科的大小按种数多少统计,见表2。
含10种以上的科有4科,计49属,85种,分别占属、种总数的37.4%和43.8%。这些科包含了本地区近半的属种,其中禾本科(Gramineae)、莎草科(Cyperaceae)、菊科(Compositae)和蓼科(Polygonaceae)的一些种类是滩涂草本群落的优势种,说明新安江流域滩涂湿地草本被子植物区系中优势科明显。
10种以下的小科和极小科有44科,82属,109种,分别占科、属、种总数的91.7%、62.6%和56.2%。这些类群在群落中虽然不占主导地位,但体现了该区植物区系的复杂性和多样性。

表2 新安江干流湿地植物科内种的组成Table 2 The composition of species in families of wetland plants in Xin'anjiang River
(2)科的分布区类型
世界分布的科主要有禾本科、莎草科、菊科、浮萍科(Lemnaceae)等21科,占43.8%;其次为泛热带分布的科,主要有大戟科(Euphorbiaceae)、鸭跖草科(Commelinaceae)、雨久花科(Pontederiaceae)、水鳖科(Hydrocharitaceae)等19科,占39.6%;北温带分布的科主要有蓼科(Polygonaceae)、毛茛科(Ranunculaceae)、十字花科(Cruciferae)、伞形科(Umbelliferae)、桔梗科(Campanulaceae)等5科,占10.4%;其它的分布区类型都只有少数一两个科。可见,组成新安江干流滩涂湿地草本种子植物科的分布区类型比较集中,主要为世界分布和泛热带分布(表3)。
3.1.2 滩涂湿地草本种子植物属的分析
(1)属的大小分析
安徽新安江干流滩涂湿地共有草本种子植物131属,含11种的属有蓼属(Polygonum),含8种的属有莎草属(Cyperus),含6种的属有大戟属(Euphorbia),含2—5种的属有31属,单种属97属。寡种属(2—5种)和单种属共128个,含169种,占总属数的97.7%和总种数的87.1%(表4)。97个单种属中,其中单型属有大薸属(Pistia)、黑藻属(Hydrilla)、鹅肠菜属(Malachiyum)、泥胡菜属(Hemistepta)等4属。由此可见,本区系属的组成较为分散,以寡种属和单种属为主,植物成分较为复杂(表4)。

表3 新安江干流滩涂湿地草本种子植物科的分布区类型Table 3 The areal-types of spermatophytic families in wetlands of Xin'anjiang River

表4 新安江干流湿地植物属内种的组成Table 4 The composition of species in genera of wetland plants in Xin'anjiang River
(2)属的分布区类型
参照吴征镒[9]关于中国种子植物属的分布区类型的划分,新安江干流滩涂湿地草本种子植物的131个属中,除了地中海区、西亚至中亚分布、东亚分布和中国特有分布外,其它12个分布区类型在本区域均有代表(表5)。
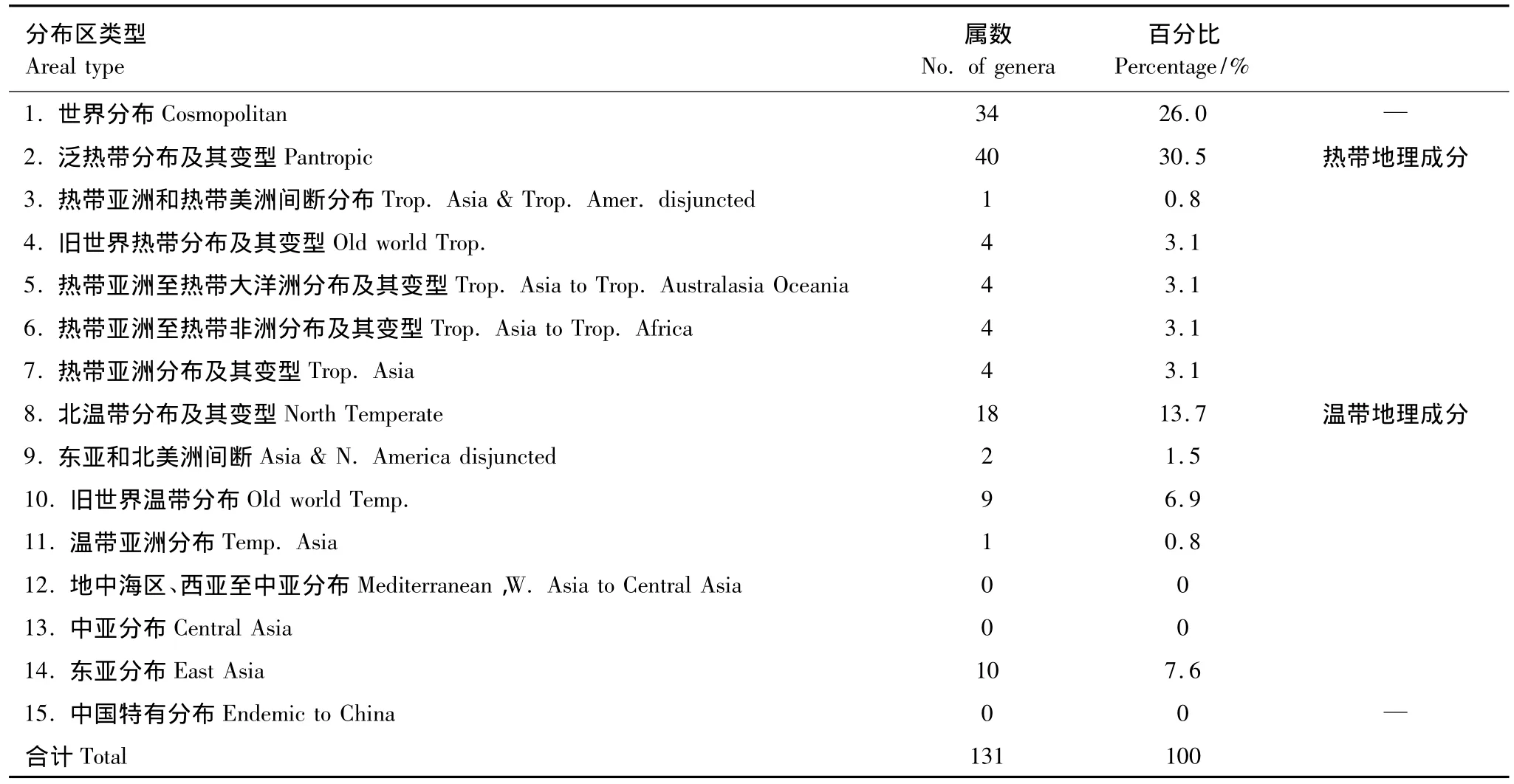
表5 新安江干流滩涂湿地草本种子植物属的分布区类型Table 5 The areal-types of spermatophytic genera in Xin'anjiang River wetlands
世界分布的共有34属,主要有莎草属、蓼属、苔草属(Carex)、蔊菜属(Rorippa)、酸模属(Rumex)、碎米荠属(Cardamine)等。
热带分布的属共57属,占属总数的43.5%。其中泛热带分布的属有40属,占属总数的30.5%,常见的有狗牙根属(Cynodon)、雀稗属(Paspalum)、水蜈蚣属(Kyllinga)、莲子草属(Alternanthera)、白茅属(Imperata)、假稻属(Leersia)、飘拂草属(Fimbristylis)、狗尾草属(Setaria)、合萌属(Aeschynomene)等,这些属在本区植被中起着比较重要的作用。热带亚洲和热带美洲间断分布的仅凤眼莲属(Eichhornia)1属。旧世界热带分布的有爵床属(Rostellularia)、乌蔹苺属(Cayratia)、水鳖属(Hydrocharis)、雨久花属(Monochoria)等4属。热带亚洲至热带大洋洲分布的有假俭草属(Eremochloa)、结缕草属(Zoysia)、黑藻属(Hydrilla)和通泉草属(Mazus)等4属。热带亚洲至热带非洲分布的有大豆属(Glycine)、荩草属(Arthraxon)、芒属(Miscanthus)和黄瓜属(Cucumis)等4属。热带亚洲分布的有翅果菊属(Pterocypsela)、鸡矢藤属(Paederia)、蛇莓属(Duchesnea)和芋属(Colocasia)等4属。
温带分布的属共40属,占30.5%,较低于热带分布属。北温带分布的属有24属,主要有稗属(Echinochloa)、蒿属(Artemisia)、委陵菜属(Potentilla)、风轮菜属(Clinopodium)等。东亚和北美洲间断分布的属有菰属(Zizania)和莲属(Nelumbo)2属。旧世界温带分布的共13属,主要有鹅观草属(Roegneria)、天名精属(Carpesium)、菱属(Trapa)和水芹属(Oenanthe)等。温带亚洲分布的仅马兰属(Kalimeris)1属。东亚分布的共10属,主要有黄鹌菜属(Youngia)、泥胡菜属(Hemistepta)、鸡眼草属(Kummerowia)和石荠苧属(Mosla)等。
新安江干流滩涂湿地草本种子植物区系地理成分复杂,分布区类型多样,过渡性明显。总体上热带成分43.5%较多于温带性成分30.5%,植物区系的热带、亚热带特征显著。这表明新安江干流地区地处亚热带,为热带向温带过渡地区,植物区系明显具有热带、亚热带与暖温带的多重性质,但以亚热带性质为主。
3.2 滩涂湿地草本植物物种多样性分析
3.2.1 不同江段滩涂湿地草本植物物种组成的变化
由表6可见,新安江干流滩涂湿地草本植物主要以禾草型的湿生草本植物为主,主要的湿地草本植物有狗牙根(Cynodon dactylon)、双穗雀稗(Paspalum distichum)、假俭草(Eremochloa ophiuroides)等。尤其是狗牙根,在所调查的滩涂湿地中它几乎都扮演着重要的角色,为群落优势种或建群种,这可能一方面是由于狗牙根等根茎型禾草具有发达的根状茎,繁殖较快,且较耐水淹;另一方面由于新安江滩涂湿地主要为沙壤土,比较适宜根茎型禾草的生长。研究发现,江段上游样地Ⅰ中植物群落以狗牙根、假俭草和双穗雀稗为优势种,三者重要值之和达44.9%;样地Ⅱ中植物群落以狗牙根、双穗雀稗和碎米莎草(Cyperus iria)为优势种,三者重要值之和达42%。在江段上游两个样地中重要值排第一位的均为狗牙根,说明狗牙根在江段上游区域的植被组成中占有非常重要的位置。江段中游样地Ⅲ中植物群落以假俭草、五节芒(Miscanthus floridulus)和狗牙根为优势种,三者重要值之和达41.5%;样地Ⅳ中植物群落以双穗雀稗、菰(Zizania latifolia)和假俭草为优势种,三者重要值之和达39.7%;样地Ⅴ中植物群落以假俭草、狗牙根和结缕草(Zoysia japonica)为优势种,三者重要值之和达53.1%;假俭草在江段中游3个样地中均为优势种,说明假俭草在江段中游滩涂湿地的植被组成中占有比较重要的位置。
江段下游样地Ⅵ中植物群落以双穗雀稗、水蓼(Polygonum hydropiper)和莎草(Cyperus rotundus)为优势种,三者重要值之和达60.3%;样地Ⅶ中植物群落以双穗雀稗、假稻(Leersia japonica)和喜旱莲子草(Alternanthera philoxeroides)为优势种,三者重要值之和达38.4%;样地Ⅷ中植物群落以狗牙根、双穗雀稗和喜旱莲子草为优势种,三者重要值之和达42.7%;双穗雀稗在江段下游3个样地中均为优势种,说明双穗雀稗在江段下游滩涂湿地的植被组成中占有非常重要的位置。
3.2.2 不同江段滩涂湿地草本植物物种丰富度的变化
这里的物种丰富度指样地内出现的物种数。不同江段滩涂湿地所调查的8个样地中出现的物种数如图2所示,可以看出,新安江干流上游滩涂湿地草本植物群落的物种丰富度较下游的高,群落的物种丰富度呈逐渐减小的格局。江段上游样地Ⅱ中草本植物群落的物种丰富度最高,出现了71种草本植物;江段下游样地Ⅵ中草本植物群落的物种丰富度最低,仅出现了35种草本植物。新安江干流下游滩涂湿地草本植物较上游物种丰富度低,一方面可能是下游滩涂湿地的淹水期普遍较上游滩涂湿地的淹水期长,很多草本植物不适宜在下游滩涂湿地生长繁殖;另一方面,由于新安江下游沿岸人口密度较大,滩涂湿地受干扰比较严重,主要表现为人为活动和放牧干扰。调查发现,一些外来入侵种成为了江段下游滩涂湿地的优势物种,如喜旱莲子草在江段下游样地Ⅶ和样地Ⅷ中重要值均较高,分布为0.087和0.142;苍耳(Xanthium sibiricum)在样地Ⅵ中的重要值也高达0.084。此外,本次调查在江段下游还首次发现了安徽新分布的一种外来入侵物种——大戟科飞扬草(Euphorbia hirta),其在下游滩涂湿地及岸边分布范围较大,在局部区域已经形成优势种群,需要引起相关管理部门的重视。
3.2.3 不同江段滩涂湿地草本植物物种α多样性的变化
新安江干流不同江段滩涂湿地草本植物群落的Simpson指数、Shannon-Wiener指数和Pielou均匀度指数变化如图2。可以看出,Simpson指数和Shannon-Wiener指数的变化格局与丰富度的变化格局是比较一致的,均表现为上游>中游>下游。江段上游滩涂湿地样地Ⅱ植物群落为狗牙根+双穗雀稗+碎米莎草群落,Simpson指数和 Shannon-Wiener指数最高,分别为0.913和2.769;江段下游样地Ⅵ中草本植物群落的Simpson指数和Shannon-Wiener指数最低,分别为0.825和1.932。Pielou均匀度指数变化格局与Simpson指数和Shannon-Wiener指数的变化格局不完全一致,但总体上变化趋势是一致的,表现为上游>中游>下游。
江段下游滩涂湿地应该原属于草本植物丰茂区域,但由于江段下游沿岸周边人口较多,江段下游滩涂湿地也就成为了当地农民活动和放牧的重要区域,调查中发现一些滩涂湿地被开垦种植农作物,这就大大降低了滩涂湿地的物种多样性,此外,由于原始的植被遭到破坏,也在一定程度上增加了局部的水土流失,严重影响了湿地的原生态景观。在湿地区域进行适量牛羊等家畜放牧,增加了湿地生态保育区的生物多样性美感,营造了一种人与自然和谐共存的景观效果[2]。但由于过度的放牧干扰,使该区域草本植物动态变化较大,假俭草、狗牙根等适口性较好的草本植物在放牧胁迫下虽然具有一定补偿能力,但随着放牧强度的增加,牧草类草本生物多样性骤减[2,14],放牧对植物种群的影响最终都将反映在植物的构件和种群结构上,植物对适度放牧具备一定的再生恢复能力,但超过了一定限度,植物将大幅度减少,不再具备恢复能力,从而引起植被的退化,对植被造成了严重的不利影响[15-16]。研究表明,频繁的人类活动以及人类活动导致的土地利用方式的改变可能增加外来种迁入和定居的机会,并促进外来种的扩散和传播[17-20]。由于江段下游当地居民在滩涂湿地上开垦种植农作物,并进行过度放牧,江段下游滩涂湿地受到了较大程度的人为干扰,湿地中外来入侵种的数量明显多于江段上游和中游,且一些外来入侵种如喜旱莲子草、苍耳等已经取代一些本土植物成为江段下游滩涂湿地的优势种。这不仅影响了植物景观多样性,还降低了群落物种多样性,进而对滩涂湿地生态系统的结构、功能及生态环境产生严重的干扰与危害。

图2 上游至下游滩涂湿地草本植物群落α多样性的变化Fig.2 α diversity of herbaceous plant community from upstream to downstream
3.2.4 不同江段滩涂湿地草本植物物种β多样性的变化
β多样性用相异性系数和Cody指数来表达[13]。研究表明,新安江干流不同江段滩涂湿地草本植物群落的相异性系数和Cody指数的变化趋势基本相同(图3),均呈现上升的格局。二者的最小值(0.25和17.5)均出现在江段上游样地Ⅰ和样地Ⅱ群落之间,最大值均出现样地Ⅶ和样地Ⅷ植物群落之间(0.69和29)。
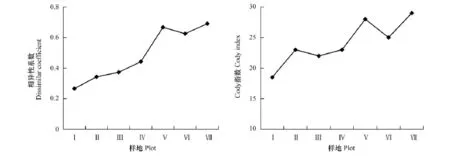
图3 上游至下游滩涂湿地草本植物群落β多样性的变化Fig.3 β diversity of herbaceous plant community from upstream to downstream
4 讨论
本文对新安江干流安徽段滩涂湿地草本植物群落物种组成及多样性进行了分析和研究,结果表明新安江干流滩涂湿地共有草本植物48科、131属、194种,其中双子叶植物37科、88属、129种,单子叶植物11科、43属、65种;该区草本植物区系地理成分复杂,分布区类型多样,物种多样性高,优势科明显,主要包括禾本科、莎草科、菊科和蓼科;科内种的组成比较复杂,较少的科含有较多的种,较多的科含有较少的种;属的组成较为分散,寡种属(2—5种)和单种属占总属数的97.7%和总种数的87.1%。
通过对新安江干流不同滩涂湿地草本植物群落物种多样性研究发现,江段上、中、下游不同滩涂湿地各植物群落的物种数量、以及建群种、优势种和伴生种的种类等组成特征具有较大差异,如假俭草在江段中游3个滩涂湿地中为建群种,处于绝对优势,但在江段下游滩涂湿地中很少见。α多样性是指某个群落或生境内部的物种多样性[21-22]。本研究得出江段干流不同流域滩涂湿地草本植物群落的α多样性格局与丰富度的格局比较一致,均表现为上游﹥中游﹥下游。造成这种α多样性格局与丰富度的变化格局的原因,主要于各滩涂湿地植物群落的生境以及受干扰程度有关。一方面可能是下游滩涂湿地的淹水期普遍较上游滩涂湿地的淹水期长,很多草本植物不适宜在下游滩涂湿地生长繁殖;另一方面由于江段下游滩涂湿地受干扰比较严重,主要表现为人为开垦湿地种植农作物和过度放牧。
β多样性可以定义为群落间的多样性,也可以定义为沿着某一环境梯度物种替代的程度或速率、物种周转率等[21-22]。相异性系数和Cody指数分别从上述两个角度反映物种多样性沿环境梯度的分布格局及变化规律,均很好地表达了β多样性的含义[13]。本研究得出的相异性系数最小值出现在江段上游样地Ⅰ和样地Ⅱ植物群落之间,最大值出现江段下游样地Ⅶ和样地Ⅷ植物群落之间。出现这种格局的原因,可能与滩涂湿地的地形地貌,淹水程度以及人为干扰程度有关,具体原因尚需进一步研究。本研究得出的Cody指数的变化格局说明,由江段上游至下游不同滩涂湿地草本植物种的替代总体上呈现逐渐加快的趋势,而物种的替代与生态位的状况有关[13],本研究中出现的这种格局可能与不同滩涂湿地土壤基质化学成分和湿地水位等有关,但也可能是由人为活动干扰造成的,具体原因尚需进一步研究。
通过实地调查发现,新安江干流下游部分滩涂湿地的植物多样性已经遭到了一定程度的人为活动的影响甚至破坏,部分区域外来物种入侵比较严重,这严重影响了滩涂湿地的生物多样性景观和微生态系统。因此,保护和恢复新安江干流下游滩涂湿地植物多样性已迫在眉睫。一方面要加强宣传,提高沿岸居民保护环境的意识,停止对滩涂湿地的开垦,另一方面,建议相关管理部门组织当地居民对滩涂湿地的一些外来入侵植物采取人工刈除,以免其入侵其它区域,造成更为严重的后果。从而使江段下游滩涂湿地植物多样性得到较大提高,再现江段上游那种人与自然和谐相伴的生态景观。
[1] Wall G.Implications of global climate change for tourism and recreation in wetland areas.Climatic Change,1998,40(2):371-389.
[2] Wang L L,Lu L,Dai J S.The herbaceous plant diversity dynamics under different disturbance and its flora in the conservation area of Taiping Lake National Wetland Park.Journal of Natural Resources,2010,25(8):1306-1319.
[3] Shen Q,Liu K,Li S Y,Zhang J,Jiang Y P,Ge Y,Chang J.Relationships of plant composition,water level and solar radiation in Xixi wetland,Hangzhou,China.Journal of Plant Ecology,2008,32(1):114-122.
[4] Pan Y F,Xu Q,Cheng Y Q,Yin L,Wang C L.Herbaceous seed flora of wetlands in Shengjin lake nature reserve of Anhui province.Wetland Science,2008,6(2):304-309.
[5] Song X Z.Characteristics and Functions of Vegetation on Down-Reach Floodplain of Yellow River in He'nan[D].Beijing:Chinese Academy of Forestry,2005,1-2.
[6] Wu T G,Wu M,Xiao J H.Dynamics of community succession and species diversity of vegetations in beach wetlands of Hangzhou bay.Chinese Journal of Ecology,2008,27(8):1284-1289.
[7] Editorial Committee of Anhui flora.The Flora of Anhui.Hefei:Anhui Science and Technology Press,1992.
[8] Lang H Q,Zhao K Y,Chen K L.Wetland Vegetation in China.Beijing:Science Press,1999.
[9] Wu Z Y.The areal-types of Chinese genera of seed plants.Acta Botanica Yunnanica,1991,13(Ⅳ):1-139.
[10] Li X W.Floristic statistics and analyses of seed plants from China.Acta Botanica Yunnanica,1996,18(4):363-384.
[11] Jin X F,Ding B Y,Zheng C Z,Ye Z L,Chen X R.The floristic analysis of seed plants in Baishanzu nature reserve from Zhejiang province.Acta Botanica Yunnanica,2004,26(6):605-618.
[12] Fang J Y,Shen Z H,Tang Z Y,Wang Z H.The protocol for the survey plan for plant species diversity of China's mountains.Biodiversity Science,2004,12(1):5-9.
[13] Lou Y J,Zhao K Y,Ma K P.Change in floristic composition and species diversity of plant community along environment gradient in Honghe National Nature Reserve,China.Acta Ecologica Sinica,2007,27(9):3883-3891.
[14] Pan S W,Wang H Y,Du G Z,Lei S Q,Wei S Q.Effects of compensation capacity and palatability on the dominance of plant species in grazing community.Chinese Journal of Applied Ecology,2008,19(8):1682-1687.
[15] An Y,Li B,Yang C,Yan Z J,Han G D.Influence of grazing rate on population structure of Stipa Grandis.Acta Phytoecologica Sinica,2002,26(2):163-169.
[16] Li Y Q,Zhao J B.Effects of overgrazing on ecological and environmental construction and measurement.Journal of Desert Research,2005,25(3):404-408.
[17] Ding J Q,Mack R N,Lu P,Ren M X,Huang H W.China's booming economy is sparking and accelerating biological invasions.BioScience,2008,58(4),317-324.
[18] Weber E,Sun S G,Li B.Invasive alien plants in China:diversity and ecological insights.Biological Invasions,2008,10(8):1411-1429.
[19] Hellmann J J,Byers J E,Bierwagen B G,Dukes J S.Five potential consequences of climate change for invasive species.Conservation Biology,2008,22(3):534-543.
[20] Yang B,Yangjin Z G,Pan X Y,Xu H G,Li Bo.Alien terrestrial herbs in China:diversity and ecological insights.Biodiversity Science,2010,18(6):660-666.
[21] Magurran A E.Ecological Diversity and Its Measurement.New Jersey:Princeton University Press,1988.
[22] Ma K P.The measurement of community diversity//Qian Y Q,Ma K P,eds.Principles and Methodologies of Biodiversity Studies.Beijing:Chinese Science and Technology Press,1994:141-165.
参考文献:
[2] 王立龙,陆林,戴建生.太平湖国家湿地公园生态保育区草本植物区系及其在不同干扰下的多样性动态.自然与资源学报,2010,25(8):1306-1319.
[3] 沈琪,刘珂,李世玉,张骏,蒋跃平,葛滢,常杰.杭州西溪湿地植物组成及其与水位光照的关系.植物生态学报,2008,32(1):114-122.
[4] 潘云芬,徐庆,程元启,尹莉,王翠莲.安徽升金湖自然保护区湿地草本种子植物区系研究.湿地科学,2008,6(2):304-309.
[5] 宋绪忠.黄河下游河南段滩地植被特征与功能研究[D].北京:中国林业科学研究院,2005:1-2.
[6] 吴统贵,吴明,萧江华.杭州湾滩涂湿地植被群落演替与物种多样性动态.生态学杂志,2008,27(8):1284-1289.
[7] 安徽植物志编委会.安徽植物志.合肥:安徽科学技术出版社,1992.
[8] 郎惠卿,赵魁义,陈克林.中国湿地植被.北京:科学出版社,1999.
[9] 吴征镒.中国种子植物属的分布区类型.云南植物研究,1991,13(增刊Ⅳ):1-139.
[10] 李锡文.中国种子植物区系统计分析.云南植物研究,1996,18(4):363-384.
[11] 金孝锋,丁炳扬,郑朝宗,叶珍林,陈小荣.浙江百山祖自然保护区种子植物区系分析.云南植物研究,2004,26(6):605-618.
[12] 方精云,沈泽昊,唐志尧,王志恒.“中国山地植物物种多样性调查计划”及若干技术规范.生物多样性,2004,12(1):5-9.
[13] 娄彦景,赵魁义,马克平.洪河自然保护区典型湿地植物群落组成及物种多样性梯度变化.生态学报,2007,27(9):3883-3891.
[14] 潘声旺,王海洋,杜国祯,雷抒情,魏世强.补偿能力和适口性对放牧群落植物优势度的影响.应用生态学报,2008,19(8):1682-1687.
[15] 安渊,李博,杨持,闫志坚,韩国栋.不同放牧率对大针茅种群结构的影响.植物生态学报,2002,26(2):163-169.
[16] 李瑜琴,赵景波.过度放牧对生态环境的影响与控制对策.中国沙漠,2005,25(3):404-408.
[20] 杨博,央金卓嘎,潘晓云,徐海根,李博.中国外来陆生草本植物:多样性和生态学特性.生物多样性,2010,18(6):660-666.
[22] 马克平.生物群落多样性的测度方法//钱迎倩,马克平.生物多样性研究的原理和方法.北京:科学出版社,1994:141-165.
猜你喜欢
中国水利(2020年23期)2020-12-24
科学导报(2020年27期)2020-05-13
世界热带农业信息(2019年11期)2019-01-05
照相机(2017年10期)2017-11-22
华东师范大学学报(自然科学版)(2017年1期)2017-02-27
中国资源综合利用(2016年6期)2016-01-22
中国资源综合利用(2016年3期)2016-01-22
学习月刊(2015年23期)2015-07-09
水电站机电技术(2014年3期)2014-09-26
山西大同大学学报(自然科学版)(2014年6期)2014-01-23
