
黄泥河自然保护区狍冬季卧息地选择
2013-09-07 02:53:02朱洪强葛志勇姜春艳张冬冬张香东常素慧毛之夏
生态学报 2013年7期
朱洪强,葛志勇,刘 庚,姜春艳,张冬冬,张香东,常素慧,毛之夏
(1.吉林农业大学,中药材学院,长春 130118;2.中国科学院东北地理与农业生态研究所,长春 130012)
野生动物对栖息地的选择和利用是一种适应环境的行为,这种行为与栖息地的气候、环境以及动物本身生理状况等条件密切相关[1-2]。卧息地作为栖息地的重要组成部分,一直被看作是贮存能量和反捕食的策略[3],因此在冬季野生动物对卧息地的选择既要能够抵御此时的极端气候和环境,补充此时对能量的巨大消耗,还要躲避天敌的捕杀[4-6]。
狍(Capreolus capreolus)隶属于鹿科狍属,是一种广泛分布于欧亚大陆的中小型鹿科动物[7]。狍是我国重要的野生经济动物,栖息环境多样,在维护生态系统结构,完善生态系统功能和保护生物多样性中具有重要意义[8]。近年来,虽然国内外关于狍卧息地选择的研究已有很多报道,但由于气候及环境条件的不同,各地区之间狍卧息地选择策略存在明显的地区差异性。例如,滕丽微等人对黑龙江省三江自然保护区以及小兴安岭南部低山丘陵区狍冬季栖息地的研究[3,7]以及Mysterud和Østbye对挪威南部狍卧息地选择的研究都表明不同地区的狍在卧息地选择上存在明显的地区差异[9]。在黄泥河自然保护区,狍还是濒危野生动物东北虎(Panthera tigris altaica)等食肉动物的猎物,是大型濒危食肉动物种群恢复的基础[10]。深入研究该地区狍冬季卧息地选择,对于了解该地区狍冬季的生存状况和卧息地质量以及保护和管理该地区的狍资源具有重要意义。在黄泥河自然保护区冬季,1月为最冷月,最深积雪厚度出现在12月—1月,此时保护区内气候和环境最严酷,是决定狍能否顺利度过冬天的最关键时期,此时狍对卧息地的选择最能代表其对冬季恶劣气候和环境的适应。基于此,于2009年12月至2010年1月对黄泥河自然保护区狍冬季卧息地选择进行了调查研究,并期望通过该研究探讨:(1)在该地区冬季狍对卧息地是否具有选择性?(2)如果该地区冬季狍对卧息地具有选择性,哪些生态因子在卧息地选择中起决定作用?
1 研究区域概况
黄泥河自然保护区位于吉林省延边朝族自治州敦化市西北部,地理坐标为127°51'—128°14'E,43°55'—44°06'N,保护区总面积41 583 hm2,是以保护北温带山地森林生态系统和多种珍稀濒危野生动植物为主的省级自然保护区。保护区地势北高南低,地形垂直变化大,北部老白山海拔1 969 m,南部海拔在300 m以下,年平均气温为2.4℃左右,最冷月在1月,极端最低温-39.4℃,最深积雪雪被厚度出现在12月—1月,最深雪被厚度达55 cm以上,年降水量约为632 mm,在保护区内随着地势高低变化,温度和降水呈现显著的垂直变化特征,因而保护区内的植被呈现出明显的垂直地带性。根据黄泥河自然保护区植被分布规律和本次研究特点,我们将研究区域分为3个垂直分布植被带:阔叶林,针阔混交林和针叶林[11]。保护区内植物共计134科863种,其中12种国家级保护植物;脊椎动物74科231种,属于国家级保护动物31种,国家Ⅰ级重点保护野生动物有东北虎、紫貂(Martes zibellina)、金雕(Aquila chrysaetos)和原麝(Moschus moschiferus)。
2 研究方法
2.1 野外调查
2009年12月至2010年1月,根据黄泥河自然保护区狍的数量和分布,在狍活动比较集中的地区采用样线法,即在林相图上随机选取样点,然后沿与等高线垂直方向设置样线47条[2,12-13],样线间距1 500 m以上,样线长3—5 km,样线覆盖了保护区内各主要植被类型[14]。由于保护区内高海拔地区积雪较深,因此样线大部分在海拔900 m以下,仅有一条样线海拔在900 m以上,这条样线最高点海拔为943 m,雪被厚度86 cm,调查时已经无法再向上行进。实地调查过程中发现狍卧迹时,便以该卧迹为中心作一个2 m×2 m和一个10 m×10 m的样方,样方中心用GPS定位。同时,在调查行进过程中,每行走1000 m作一个2 m×2 m和10 m×10 m的样方作为卧迹样方的对照样方,如果此处恰好遇见狍卧迹,则放弃对照样方的数据统计,仅作卧迹样方统计。在黄泥河自然保护区,除狍以外还有野猪(Sus scrofa)、马鹿(Cervus elaphus)和原麝3种有蹄类动物,不同有蹄类的卧迹可根据足迹链特征、粪便形态等加以区分。在每个样方内详细测量记录海拔、坡度、坡向、坡位、隐蔽水平、郁闭度、食物丰富度、灌丛盖度、树胸径、人为干扰距离、林型、裸岩、雪深、倒木和针叶树15类生态因子。总计测量了72个狍卧息地利用样方和109个非利用样方。各类生态因子的划分[2,15]如下:
海拔 样方所处地的海拔高度,由GPS测定。
坡度 用arcgis求出样方所在地坡度,平坡≤10°、10°<缓坡<30°、陡坡≥30°。
坡向 用arcgis求出整个样方所处地的坡向,分为1阳坡(S67.5°E—S22.5°W);2半阴半阳坡(N22.5°E—S67.5°E、S22.5°W—N67.5°W);3 阴坡(N67.5°W—N22.5°E)。
坡位 整个样方所处地的坡位,分为三级,即1上坡位:山坡的上1/3和山顶、2中坡位:山坡的中间1/3部分、3下坡位为山坡的下1/3部分和山谷。
隐蔽水平 在样方中心点处测算4个方向上的可视距离,然后求其平均值。
郁闭度 估算样方内乔木树冠盖度,即乔木树冠对林下层的遮盖率,所有郁闭度的估算均由同一人完成[15]。
食物丰富度 估算样方中狍主要采食食物的覆盖率,主要统计了高度低于1.5 m的榆(Ulmus pumila Linn.)、紫椴(Tilia amurensis Rupr.)、山杨(Populus davidiana)、柳(Salix matsudana)、水曲柳(Fraxinus mandshurica Rupr.)、白桦(Betula platyphylla Suk)、风桦(Betula costata Trautv.)、蒙古栎(Quercus mongolica)、红松(Pinus koraiensis)的嫩枝嫩芽以及可见苔藓等的覆盖率,所有食物丰富度的估算均由同一人完成[16-17]。
灌丛盖度 估算样方内高度低于2.5 m的灌丛以及乔木的覆盖度,用百分率表示,所有灌丛盖度的估算均由同一人完成[15]。
树胸径 在样方中,距中心点最近的一棵高度大于2.5 m的乔木树胸径。
人为干扰距离 用arcgis计算样方中心点到最近人为干扰源(包括主要干道,人类定居点等)的垂直距离。
林型 样方所在地优势乔木的生长型外貌,分为1阔叶林、2针阔混交林、3针叶林。
裸岩 统计样方内是否有裸岩,无记为0,有记为1。
雪深 测量记录样方中心点处的雪深。
倒木 统计样方内是否有树胸径大于15 cm的倒木,无记为0,有记为1。
针叶树 观测记录2 m×2 m样方内是否有针叶树,落叶松不在统计范畴内。无记为0,有记为1。
2.2 数据处理
采用Baileys置信区间法对坡向、坡位、林型、裸岩、倒木和针叶树6种定类变量进行分析,判断在黄泥河自然保护区狍冬季对这些卧息地因子选择的差异性[18-19]。
利用单样本Kolmogorov-Smimov Test对海拔、坡度、隐蔽水平、郁闭度、食物丰富度、灌丛盖度、树胸径、人为干扰距离和雪深这9种数量型变量的原始数据进行正态分布检验,对数据符合正态分布(P>0.05)的郁闭度、食物丰富度和树胸径3类变量利用两独立样本t检验进行比较;对不遵从正态分布(P<0.05)的其他变量先进行反正弦正态性转换,由于转换后仍然不遵从正态分布,故采用非参数估计中Mann-Whitey U检验对这些数量型卧息地因子选择的差异性进行比较。
为了保证黄泥河自然保护区冬季狍卧息地各变量之间的独立性,将上述两步数据分析中存在显著性差异的坡向、坡位、林型、裸岩、针叶树、海拔、隐蔽水平、郁闭度、灌丛盖度、食物丰富度和雪深11个变量采用Spearman法进行相关性分析。相关分析表明坡位与海拔、林型之间的相关系数绝对值均大于0.6。根据野外调查,海拔和林型在狍卧息地选择中的影响较为明显,因此在进一步逻辑斯蒂回归分析前剔除坡位这个变量。

表1 定类变量编码Table1 Categorical variables coding
将样方利用与否作为因变量(对照样方=“0”,利用样方=“1”),将坡向、林型、裸岩、针叶树、海拔、隐蔽水平、郁闭度、灌丛盖度、食物丰富度和雪深10种存在显著性差异的卧息地因子作为自变量,采用Backward conditional法(条件向后逐步剔除法)进行二分逻辑斯蒂回归分析,以筛选出影响黄泥河自然保护区冬季狍卧息地选择的主要生态因子。因为自变量中存在坡向、林型、裸岩和针叶树4个定类变量,不能直接进行逻辑斯蒂回归分析,需对这些定类变量设置亚变量[20]。坡向和林型均含有3种质因素,故设置2个亚变量,分别为坡向(1)和坡向(2)以及林型(1)和林型(2)。裸岩和针叶树为二分定类变量,均设置1个亚变量,分别为裸岩(1)和针叶树(1)。在设置亚变量时,均以最后一类质因素为参考类,亚变量编码如表1。
以上所有的数据处理全部在Excel 2003和SPSS 18.0 for Windows软件包中进行。
3 结果
3.1 定类变量的Baileys法分析
采用Baileys置信区间法对狍卧息地选择中的6种名词型变量进行分析,结果显示狍喜欢在处于中坡位阳坡的针阔混交林中卧息,避开在阴坡和有裸岩的地区卧息,对倒木的选择不显著。狍在卧息时对针叶树表现出明显偏好,但同时又避开选择纯针叶林(表2)。
3.2 数量型变量的比较
对黄泥河自然保护区狍冬季卧息地利用样方和对照样方的数量型变量进行比较(表3),可以发现利用样方的主要特征表现为海拔低,雪被厚度浅,有较高的食物丰富度,灌丛盖度、郁闭度和隐蔽水平,但是在利用样方与对照样方的比较中,树胸径、坡度和人为干扰距离3类卧息地因子差异不显著。
3.3 狍冬季卧息地选择的逻辑斯蒂回归分析
在本次逻辑斯蒂回归分析中,最终进入方程有明显统计学意义的变量为坡向、林型、裸岩、针叶树、海拔、隐蔽水平、食物丰富度和雪深。最终模型的拟合优度检验各参数分别为:-2log(likelihood)=35.337,Nagelkerke R2=0.924,说明模型对数据的拟合优度很高。

表2 狍对每种生境类型的利用和选择Table2 Selection and utilization of each habitat types by roe deer in Huangnihe nature reserve

表3 狍数量型卧息地生态因子的比较(Mean±SD)Table3 Comparison of numerical ecological factors on bed-site selection by roe deer in Huangnihe nature reserve
由表4可知,食物丰富度、针叶树、雪深、裸岩和海拔是影响黄泥河自然保护区冬季狍卧息地选择的主导因子,林型、坡向和隐蔽水平是次要因子。在只含有常数项的起始模型中,Wald统计量的显著性水平P=0.006<0.05,所以常数项参数有效。因此狍对卧息地选择的最终逻辑斯蒂回归模型为:Z=32.628+11.675×坡向(1)+9.741×坡向(2)-5.486×林型(1)-7.933×林型(2)-7.496×裸岩(1)-9.906×针叶树(1)-0.043×海拔+0.170×隐蔽水平+0.220×食物丰富度-0.429×雪深,选择利用概率P(z)=ez/1+ez,模型整体正确预测率为96.1%。
4 讨论
影响野生动物卧息地选择的因素复杂多变,这与动物的本身特性、栖息地特征、食物分布、隐蔽场所、竞争和捕食等因素息息相关[21-22]。在黄泥河自然保护区,冬季狍对卧息地表现出明显的选择性。在保护区,狍在卧息时喜欢海拔中等,位于阳坡中坡位上食物丰富度、灌丛盖度、郁闭度和隐蔽级都较高、雪被较浅的针阔混交林生境,尤其喜欢在针阔混交林中的针叶树下卧息,避开选择阴坡、裸岩和典型针叶林生境。
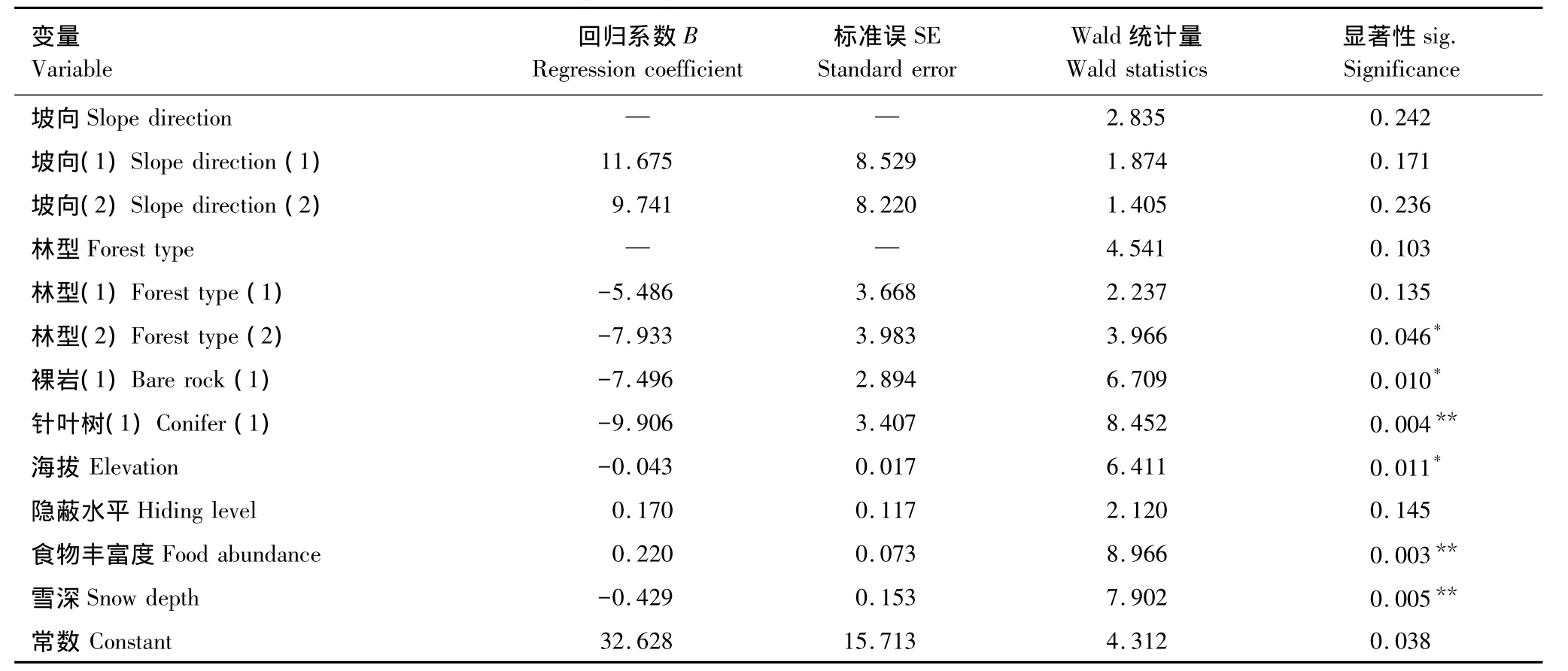
表4 狍卧息地选择逻辑斯蒂回归分析Table4 Logistic regressions for bed-site selection by roe deer
针叶树(不包括落叶松)这一单一生态因子对狍冬季卧息地选择的影响在国内外未见报道。在黄泥河自然保护区,针叶树是影响狍卧息地选择的重要生境因子(表2、表4),这可能与天然保温覆盖物有关。植被结构可以对微生境产生影响,乔木和灌木等天然保温覆盖物在狍卧息地选择中具有重要作用[7,23-24]。在冬季,针叶树树冠可以改变雪的降落方向,在调节温度、辐射和降雪方面要比落叶之后的阔叶树更为明显,这对于体表面积较大的狍减少能量散失、维持能量收支平衡具有重要作用。而且,狍在卧息时有一种独特行为是将卧迹处的积雪刨走,这种行为被认为是使卧息地更加舒适和保存能量的一种策略[3,25],在黄泥河自然保护区冬季野外调查时也发现了狍的这一行为。显然,在雪被较深的地区刨除积雪明显要比在雪被浅的地区刨除积雪消耗更多的能量,不利于在严酷的冬季维持能量收支平衡,这可能正是狍选择雪被较浅的地区卧息的原因(表3、表4)。而冬季针叶树的树冠能够改变雪的降落方向,阻碍一部分积雪落到针叶树下,使得相同海拔地区针叶树下的雪被厚度明显比阔叶树下和空旷地带的雪被厚度浅,从而降低了狍在刨除积雪时的能量消耗。狍在针叶树下卧息正是对冬季严酷气候和环境的一种适应。
在黄泥河自然保护区冬季,针叶树虽然是影响狍卧息地选择的重要因素,但是由表2可知狍在卧息地选择时会避开典型的针叶林,偏好选择针阔混交林,这与Mysterud和Østbye对挪威南部狍卧息地选择的研究[9]以及滕丽微等人对小兴安岭南部低山丘陵区狍卧息地选择的研究[3]得出的狍喜欢在针叶林下卧息的结果有明显不同。造成这种差异的原因一方面可能是由不同地区间的环境、气候差异造成的;另一方面也可能是由于这些研究中没有把针阔混交林中的针叶树作为一个独立生态因子对其单独考虑引起的。在黄泥河自然保护区冬季,狍喜欢在针叶树下卧息,但又同时避开典型针叶林的现象说明针叶树并不是影响狍卧息地选择的唯一因素,而是由针叶树和其他多个因素共同决定的,同时也说明针阔混交林中针叶树下的小气候更能满足狍的卧息需求。
在黄泥河自然保护区冬季狍喜欢在阳坡中坡位的针阔混交林中卧息(表2),这与国内外的许多研究结果相似[3,9]。完达山地区和小兴安岭地区与黄泥河自然保护区同处于动物地理区划中的东北区,大尺度下的气候环境相似,冬季寒冷,雪被较厚,食物匮乏,这可能是造成研究结果相似的原因。海拔是影响狍卧息地选择的重要生态因子(表3、表4),而中坡位海拔一般相对较低、雪被较浅,阳坡又背风向阳,环境条件相对温和,这样的卧息地组合显然对狍具有较高的吸引力。此外,针阔混交林不但有狍卧息时需要的针叶树,而且针阔混交林中林下地面植被较多,这些植被的嫩枝嫩芽能够满足狍的食物需求,避免了狍在取食地和卧息地之间长距离迁移造成的能量耗损,符合以最小的能量损耗获取最多能量的最佳取食原则。
卧息地选择是有蹄类动物的一种贮存能量策略[1]。在黄泥河自然保护区冬季,食物丰富度对狍卧息地选择的影响(表3、表4)恰好能说明这一点,丰富的食物是狍顺利度过冬天的关键。捕食压力对野生动物的卧息地选择有重要影响,动物可以通过选择卧息地生境降低被捕食风险[1,21]。在本次研究中,虽然灌丛盖度最终未落入逻辑斯蒂回归模型中,隐蔽水平对模型的影响也不显著,但是从表3中可以看出,在黄泥河自然保护区冬季,狍在卧息时喜欢选择隐蔽水平和灌丛盖度均较高的生境,这与滕丽微等人对小兴安岭南部狍卧息地选择的研究结果一致[3]。灌丛可以为狍提供隐蔽物,隐蔽水平较高的卧息地能够降低狍被捕食风险。而且,灌丛盖度能够在一定程度上降低风速,改变卧息环境小气候,进一步弥补狍在保温覆盖物上选择的不足,这些对狍顺利度过寒冬都至关重要,明显提高了对狍卧息地选择时的吸引力。
致谢:感谢吉林省林业厅以及黄泥河自然保护区保护处对野外调查工作提供的支持。
[1] Liu Z S,Cao L R,Zhai H,Hu T H,Wang X M.Winter habitat selection by Red deer(Cervus elaphus alxaicus)in Helan Mountain,China.Zoological Research,2004,25(5):403-409.
[2] Luo Y,Zhang M M,Liu Z S,Li Z G,Hu T H,Zhai H.Winter and spring habitat selection of Red deer(Cervus elaphus alxaicus)in the Helan Mountain,China.Acta Ecologica Sinica,2009,29(5):2757-2763.
[3] Teng L W,Liu Z S,Zhang E D,Ma J Z.Winter bed-site selection of Capreolus capreolus in low mountain areas of southern Xiaoxing'anling Mountains.Chinese Journal of Ecology,2007,26(2):213-218.
[4] Mautz W W.Sledding on a bushy hillside:the fat cycle in deer.Wildlife Society Bulletin,1978,6(2):88-90.
[5] Parker K L,Robbbins C T.Thermo regulation in mule deer and elk.Canadian Journal of Zoology,1984,62(7):1409-1422.
[6] Schmitz O J.Thermal constraints and optimization of winter feeding and habitat choice in white-tailed deer.Ecography,1991,14(2):104-111.
[7] Teng L W,Liu ZS,Zhang E D,Ma JZ.Winter bedding site selection by the roe deer(Capreoluscapreolus)in Sanjiang National Nature Reserve,Heilongjiang Province,China.Zoological Research,2006,27(4):403-410.
[8] Gao Z S,Ma JZ.Ecological value of wild deer and the influence factors.Journal of Economic Animal,2004,8(1):13-15.
[9] Mysterud A,Østbye E.Bed-site selection by European roe deer(Capreolus capreolus)in southern Norway during winter.Canadian Journal of Zoology,1995,73(5):924-932.
[10] Zhang Z C.Studies on Current Status of Amur Tiger's Prey Population Resources and Habitat use of Amur T-iger in Wandan Mountains of Heilongjiang Province[D].Harbin:Northern Forestry University,2006.
[11] Bu Z J,Wang SZ,Lang H Q,Guo K Q.Vegetation vertical zone spectrum and its features on southern slope of Laobai Mountain in Huangnihe Nature reserve.Journal of Mountain Science,2003,21(1):80-84.
[12] Zhou L L.Habitat Selection of Musk Deer(Moschus moschiferus)in Tonghe Forest Area during Summer and Winter in Lesser Khingan Mountains[D].Harbin:Northern Forestry University,2005.
[13] Hu Z J,Wang Y,Xue W J,Jiang H R,Xu H F.Studies on habitat selection by moschus berezovskii in winter in Zibaishan Nature Reserve.Journal of Henan University:Natural Science,2006,36(1):70-74.
[14] Zhang J N.Frequently-Used Experimental Method and Technology of Ecology.Beijing:Chemical Industry Press,2007:125-126.
[15] Borkowski J,Ukalska J.Winter habitat use by red and roe deer in pine-dominated forest.Forest Ecology and Management,2008,255(3/4):468-475.
[16] Song Y,Song G H,Zhang W.Study on the diet nature of capreolus capreolus in winter in Heilongjiang Fenglin Nature Reserve.Forestry Science and Technology,2001,26(6):31-34.
[17] Storms D,Said S,Fritz H,Hamann JL,Saint-Andrieux C,Klein F.Influence of hurricane Lothar on red and roe deer winter diets in the Northern Vosges,France.Forest Ecology and Management,2006,237(1/3):164-169.
[18] Thomas D L,Taylor E J.Study designs and tests for comparing resource use and availabilityⅡ.Journal of Wildlife Management,2006,70(2):324-336.
[19] Rong K,Zong C,Ma J Z.A method for analysis of habitat selection data:bailey's interval.Zoological Research,2009,30(2):215-220.
[20] Zhang W T.Statistical Analysis Tutorials of Tool Spss 11.0(advanced level).Beijing:Beijing Publishing House of Electronics Industry,2002:91-100.
[21] Houtman R,Dill L M.The influence of predation risk on diet selectivity:a theoretical analysis.Evolutionary Ecology,1998,12(3):251-262.
[22] Chu H J,Jiang Z G,Jiang F,Ge Y,Tao Y S,Li B.Summer and winter bed-site selection by goitred gazelle(Gazella subgutturosa sairensis).Zoological Research,2009,30(3):311-318.
[23] Armstong E,Euler D,Racey G.Winter bed-site selection by white-tailed deer in central Ontario.Journal of Wildlife Management,1983,47(3):880-884.
[24] Lang B K,Gates J E.Selection of sites for winter night beds by white-tailed deer in a hemlock-northern hardwood forest.American Midland Naturalist,1985,113(2):245-254.
[25] Sergeant G A,Eberhardt L E,Peek JM.Thermoregulation by mule deer(Odocoileus hemionus)in arid rangelands of southcentral Washington.Journal of Mammalogy,1994,75(2):536-544.
参考文献:
[1] 刘振生,曹丽荣,翟昊,胡天华,王小明.贺兰山区马鹿对冬季生境的选择性.动物学研究,2004,25(5):403-409.
[2] 骆颖,张明明,刘振生,李志刚,胡天华,翟昊.贺兰山马鹿冬春季生境的选择.生态学报,2009,29(5):2757-2763.
[3] 滕丽微,刘振生,张恩迪,马建章.小兴安岭南部低山丘陵地区狍冬季卧息地选择.生态学杂志,2007,26(2):213-218.
[7] 滕丽微,刘振生,张恩迪,马建章.黑龙江省三江自然保护区狍对冬季卧息地的选择.动物学研究,2006,27(4):403-410.
[8] 高智晟,马建章.野生鹿类的生态价值及其影响因素.经济动物学报,2004,8(1):13-15.
[10] 张常智.黑龙江省完达山地区东北虎猎物种群现状及东北虎生境利用研究[D].哈尔滨:东北林业大学,2006.
[11] 卜兆君,王升忠,郎慧卿,郭克勤.黄泥河自然保护区老白山南坡植被垂直带谱及其特点.山地学报,2003,21(1):80-84.
[12] 周玲玲.小兴安岭通河林区原麝(Moschus moschiferus)夏冬季生境选择研究[D].哈尔滨:东北林业大学,2005.
[13] 胡忠军,王淯,薛文杰,姜海瑞,徐宏发.紫柏山自然保护区林麝冬季生境选择.河南大学学报:自然科学版,2006,36(1):70-74.
[14] 章家恩.生态学常用实验研究方法与技术.北京:化学工业出版社,2007:125-126.
[16] 宋影,宋国华,张伟.黑龙江省丰林自然保护区狍冬季食性的研究.林业科技,2001,26(6):31-34.
[19] 戎可,宗诚,马建章.Bailey's方法在生境选择研究中的应用.动物学研究,2009,30(2):215-220.
[20] 张文彤.世界优秀统计工具SPSS11.0统计分析教程(高级篇).北京:北京希望电子工业出版社,2002:91-100.
[22] 初红军,蒋志刚,蒋峰,葛炎,陶永善,李斌.鹅喉羚夏季和冬季卧息地选择.动物学研究,2009,30(3):311-318.
猜你喜欢
黑龙江工程学院学报(2020年5期)2020-10-21 05:37:10
测绘学报(2019年11期)2019-11-20 01:31:42
水土保持研究(2019年6期)2019-10-19 03:33:32
现代农业研究(2017年2期)2017-04-01 12:29:22
科学与财富(2016年26期)2016-12-01 22:14:22
土壤与作物(2015年3期)2015-12-08 00:46:55
防护林科技(2015年5期)2015-06-10 11:42:52
海南热带海洋学院学报(2015年5期)2015-03-14 09:51:07
现代农业科技(2014年13期)2014-09-18 13:20:07
植物营养与肥料学报(2012年3期)2012-10-26 02:54:52
