
2 个三系杂交稻保持系航天诱变效应的研究
2013-08-29 09:30:26刘永柱许立超张建国陈志强
华南农业大学学报 2013年3期
刘永柱 ,许立超 ,郭 涛 ,王 慧 ,张建国,陈志强
(1 华南农业大学,国家植物航天育种工程技术研究中心,广东 广州 510642;2 广东省农业科学院 水稻研究所,广东 广州 510640)
水稻航天育种技术(Rice space flight breeding)是将水稻干种子搭载返回式航天器(卫星)经过空间诱变作用产生变异,在地面选择有益变异培育新种质、新品种的育种方法.实践证明,航天诱变育种技术在水稻优异新种质(基因)创制和培育农作物新品种上已发挥其独特的优势和作用,尤其能创造出一些其他方法和途径难以获得的罕见突变体,是农作物遗传改良的新途径[1-18].
水稻杂种优势利用为我国水稻生产做出了巨大贡献,水稻雄性不育系是杂种优势利用的基础,传统的不育系选育方法主要是利用杂交、回交等手段,难以打破不良性状之间的紧密连锁,造成恢复系和不育系亲缘关系太近,遗传背景单一[19-23].
目前,利用航天诱变技术开展三系杂交稻保持系和不育系选育和改良研究还较少.欧阳乐军等[24]对4 个水稻保持系进行航天搭载,结果表明保持系诱变后代突变株对不育系的不育性保持能力发生了变异,测交后代花粉育性和自交结实率都发生了不同程度的变化.王晓玲等[25]选择了一批水稻保持系进行航天诱变,经过4 代定向复选,回交转育,从Ⅱ-32B 诱变后代中鉴定出8 对BC4 代不育系,从科丰B诱变后代中转育出5 对不育系.
本研究以2 个三系杂交水稻保持系HT-B 和Y华农B 干种子为材料,2006年通过“实践八号”农业育种卫星进行搭载,返回地面后,连续多代种植跟踪调查,考察空间环境对水稻三系保持系的诱变效应,同时对高代突变体进行测交,观察测交后代花粉育性及自交结实率变化,考察保持系品种经航天诱变后对相应不育系育性保持能力的变异特性,旨在通过航天诱变,改良保持系的不良特性,并结合农艺、经济性状和品质性状的考察,选育出新的优良三系杂交稻保持系.
1 材料与方法
1.1 材料
航天诱变的水稻Oryza sativa 三系保持系HT-B和Y 华农B 干种子及其SP1、SP2、SP3代,从SP4代中选出不同类型的突变株,测交不育系HT-A 和Y华农A.
1.2 方法
1.2.1 诱变处理 取HT-B 和Y 华农B 干种子搭载“实践八号”农业育种卫星进行航天诱变处理,一部分种子留地面作对照(CK).卫星近地点高度为187 km,远地点高度为469 km,轨道倾角63°,舱内温度为7.21~20.72℃,空间重离子注量率为4.44 个·cm-2·d-1,种子所受较低LET 空间辐射的平均剂量为4.79 mGy,运行355 h,航程超过900 万km[26].
1.2.2 田间试验 经航天诱变回收的水稻种子及地面对照种子,于2007年3月—2008年12月在华南农业大学农学院试验农场进行连续4 代的田间试验.田间试验的种植、选种、留种方法参照农作物空间诱变育种规范原则[27]2-3执行,SP4代根据突变性状考察结果,选取突变株分别与HT-A、Y 华农A 进行测交,考察杂种花粉育性及套袋自交结实率.
随机选取诱变SP1代回收种子和对照种子,3 次重复进行标准发芽试验[28],SP1代发芽率、芽长、苗高、结实率的生理损伤分别以对照为100%进行换算,生理损伤=(对照-处理)/对照×100%[29].
突变体农艺、经济性状考察方法参照“水稻品种资源基本情况与主要形态农艺性状观察调查项目及规范”[30].SP2代将株高、有效穗数、穗长、结实率、千粒质量、谷粒长宽比及剑叶长宽在“对照平均数±3倍标准差”范围以外的植株和生育期明显相差5 d以上的植株定为突变株[27]34-36.育性观察参考朱英国等[31]的方法,育性划分标准参照范树国等[32]的方法.按国标法测定SP2和SP3代单株与株系的直链淀粉含量[33].
1.2.3 数据处理方法 试验数据采用DPS 6.55 版统计软件进行分析.直链淀粉含量的差异显著性测验采用t 测验[34].利用相关系数估算直链淀粉含量遗传力.
2 结果与分析
2.1 SP1代的生理损伤
经航天搭载处理的2 个保持系品种SP1代种子的发芽率、芽长、苗高和结实率均表现出一定程度的生理损伤,性状间损伤程度差异较大,HT-B 芽长损伤率最大,达40.11%,HT-B 的SP1代发芽率、芽长、苗高和结实率的生理损伤都大于Y 华农B(表1),表明不同基因型三系水稻保持系品种可能对空间环境的敏感性存在一定的差异.2 个材料SP1代群体田间表现整齐,在主要农艺经济性状上与对照没有明显变异(表2).
2.2 SP2代的性状变异
2.2.1 农艺、经济性状变异情况 SP2代根据群体大小,收获10%~20%单株,田间调查株高,室内考种.t 测验分析结果表明,2 个材料SP2代群体的主要农艺、经济状性均发生了明显的正向和负向变异(表3).与对照比较,HT-B 诱变SP2代除株高外,其他农艺经济性状均呈现负向变异,其中总粒数和穗数分别达到差异显著和极显著水平;Y 华农B 诱变SP2代总粒数和千粒质量增加,分别达到显著和极显著水平,有效穗数减少,达显著水平,说明通过诱变,对改良Y 华农B 的穗粒性状是有效的.
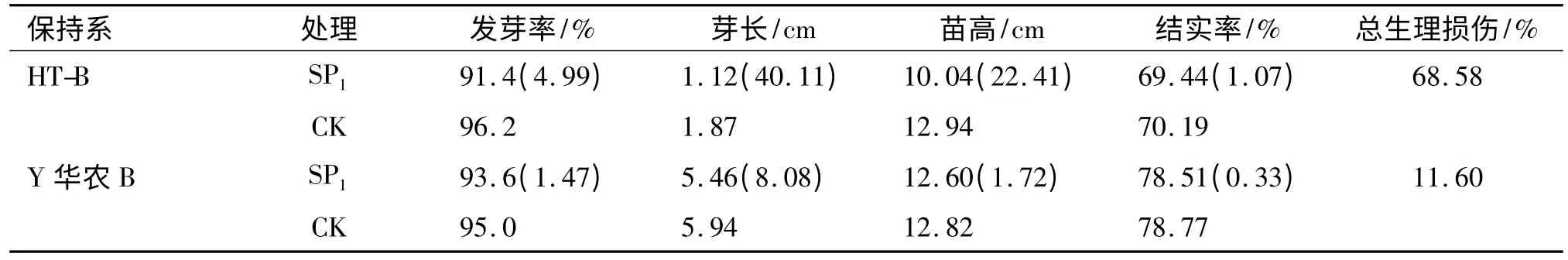
表1 诱变SP1代的生理损伤效应1)Tab.1 The physiological damage effects of space mutation of SP1mutation progeny
表2 诱变SP1代主要农艺、性状特征值Tab.2 The main agronomic and economic traits of SP1mutation progeny ±SE
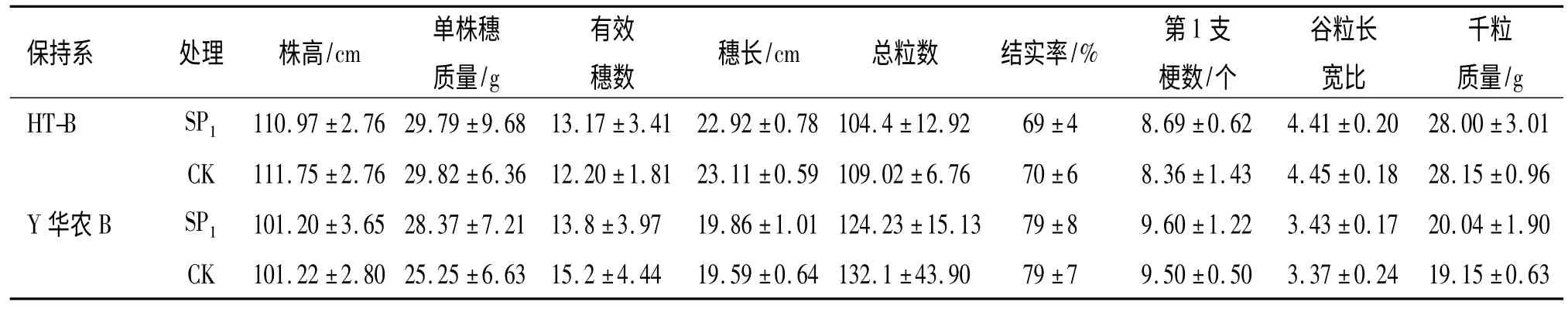
表2 诱变SP1代主要农艺、性状特征值Tab.2 The main agronomic and economic traits of SP1mutation progeny ±SE
表3 诱变SP2代主要农艺、经济性状1)Tab.3 The main agronomic and economic traits of SP2mutation progeny ±SE
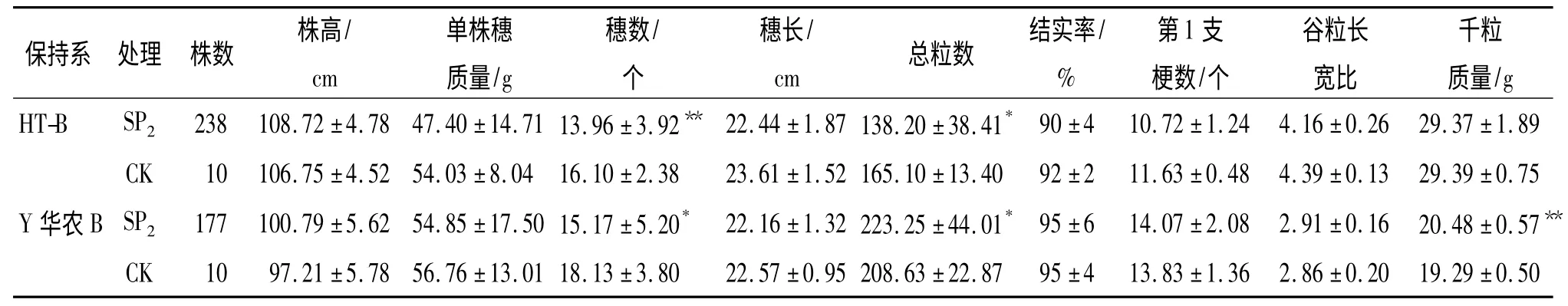
表3 诱变SP2代主要农艺、经济性状1)Tab.3 The main agronomic and economic traits of SP2mutation progeny ±SE
1)“*”、“**”分别表示相同保持系SP2代与CK 差异达0.05、0.01 显著水平(t 检验).
依据突变体鉴定标准,获得突变单株类型和数目见表4.由表4 可见,2 个保持系中分别获得26 和13 株突变体单株.HT-B 在谷粒长宽比性状上突变个体最多,而Y 华农B 出现突变个体最多的性状是千粒质量.研究还表明,2 个三系水稻保持系诱变SP2代群体的结实率与原种均无明显差异,在单株穗质量性状上均未出现突变单株(表3).HT-B 的总突变率为13.87%,高于Y 华农B的9.04%(表4).说明空间诱变能够有效地诱导农作物发生农艺性状的变异,且不同基因型品种以及同一品种的不同性状之间对空间条件的敏感性不同.

表4 诱变SP2代突变体农艺性状突变类型和突变率Tab.4 The mutant types and frequency of SP2mutation progeny
2.2.2 直链淀粉含量变异 结合农艺经济性状考察结果,分别从HT-B、Y 华农B 诱变SP2代中选取73 和96 株农艺、经济性状优良的单株测定直链淀粉质量分数(包括SP2代中筛选出的突变体单株,下同),结果如表5 所示.与对照相比,2 个保持系诱变SP2代株系的直链淀粉质量分数都发生了强烈的光谱分离,变幅极广,变异系数(CV)大.HT-B 诱变群体直链淀粉质量分数(w)大部分集中分布在低值区间(w 为9%~20%),变异幅度为1.66%~30.92%,出现了胚乳外观半透明的糯性突变体单株[w(直链淀粉)=1.66%],Y 华农B 诱变群体直链淀粉质量分数主要集中在高值区间(w≥25%),变异幅度为16.75%~38.03%.按照突变体选择标准,从HT-B 中获得24 个突变体单株,其中16 株w(直链淀粉)高于对照,8 株低于对照.从Y 华农B 中获得49个突变体单株,其中24 株w(直链淀粉)高于对照,25 株低于对照,并且出现了直链淀粉质量分数高的极端突变体[w(直链淀粉)=38.03%].说明通过空间诱变能够有效地改良杂交稻保持系直链淀粉质量分数性状,并且不同品种诱变效果不同.
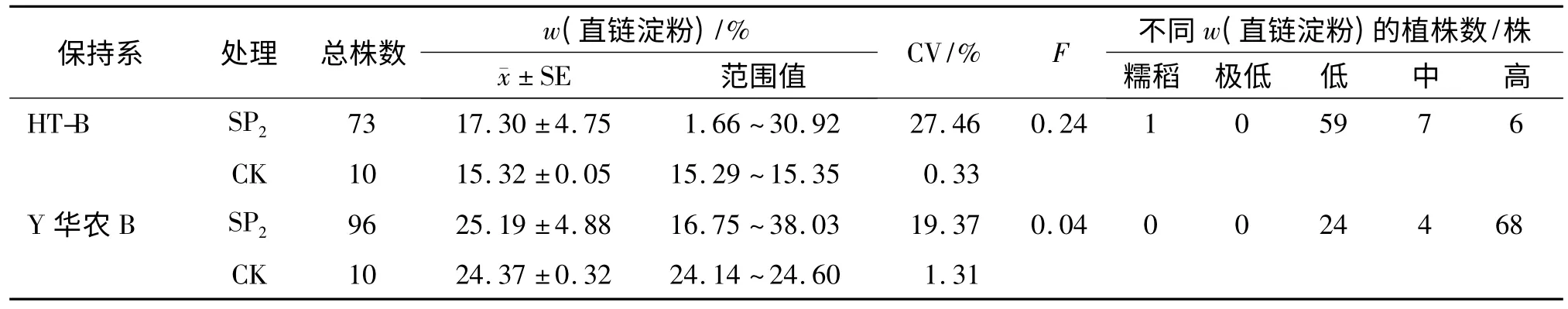
表5 诱变SP2代直链淀粉质量分数变异情况1)Tab.5 Variations of amylose content of SP2mutation Progeny
2.3 SP3代的性状表现
2.3.1 农艺、经济性状变异 经t 测验,部分SP3代突变株系农艺、经济性状表现见表6.从表6 可以看出,与原种相比,HT-B 的SP3代73 个株系中,有38个株系在株高、单株穗质量、穗长、结实率、谷粒长宽比和千粒质量上呈现分离现象,其中HT-B-14 等18个株系表现为单株穗质量变异,HT-B-4 等17 个株系表现为千粒质量变异,HT-B-22 等10 个株系表现为谷粒长宽比变异,HT-B-17 等8 个株系表现为株高变异,HT-B-17 等5 个株系表现为穗长变异,HT-B-49表现为结实率变异,HT-B-27 等17 个株系表现为至少2 个或者2 个以上的性状发生变异;而第一枝梗数、总粒数、有效穗数3 个性状变异不明显.96 个Y华农B 的SP3代株系中,有14 个株系出现变异,单株穗质量、株高、谷粒长宽比、千粒质量、有效穗数和穗长变异株系分别为8、4、3、2 和1 个,4 个株系表现为多个性状上的变异,而第1 枝梗数、总粒数、结实率3 个性状变异不明显,说明在SP2代表现不分离的单株在SP3代继续出现了分离.2 个材料从SP2代选择的突变体单株(分别为26 和13 株)农艺、经济性状变异在SP3代表现出了同样的变异趋势.
2.3.2 直链淀粉含量变异 为了减少环境对品质性状的影响,采用了相关系数估算SP2代单株与SP3代株系间直链淀粉含量的相关遗传力.结果表明,HT-B、Y 华农B 诱变SP3代与SP2代之间直链淀粉含量遗传力分别达到极显著(r=0.73**)和显著(r=0.24*)相关水平,SP3代株系直链淀粉质量分数与SP2代单株表现为相应的高低变化关系.经t 测验,HT-B 的SP3代73 个株系中,有51 个株系直链淀粉质量分数出现了变异,其中,18 个株系高于对照,w(直链淀粉)的变异幅度为22.32%~31.48%,24株变异达到显著差异,其中1 株达到极显著差异.96个Y 华农B 的SP3株系中,有4 个株系高于对照,变异范围为32.56%~34.63%.6 株表现为差异显著,其中2 株为差异极显著(部分突变体直链淀粉质量分数见表6),说明经过航天诱变,可以有效地改良水稻保持系直链淀粉含量性状.
2.4 航天诱变三系水稻保持系诱变后代恢保关系的变异
结合SP2代和SP3代农艺、经济性状和直链淀粉含量变异情况,从HT-B 和Y 华农B 诱变SP4代中分别选择了49 和70 个突变株系与其相应的不育系测交,考察测交杂种F1代花粉育性及套袋自交结实率的变化.测交F1代的可育花粉率以及套袋自交结实率值经反正弦转换后做方差分析,结果见表7.从表7 可以看出,突变株测交F1代株系间的可育花粉率及套袋自交结实率差异均达极显著水平.说明经航天诱变后,不同突变株对相应不育系的测交F1代育性恢保能力发生了较大改变,而且不同品种的不同突变株变化也不尽相同.
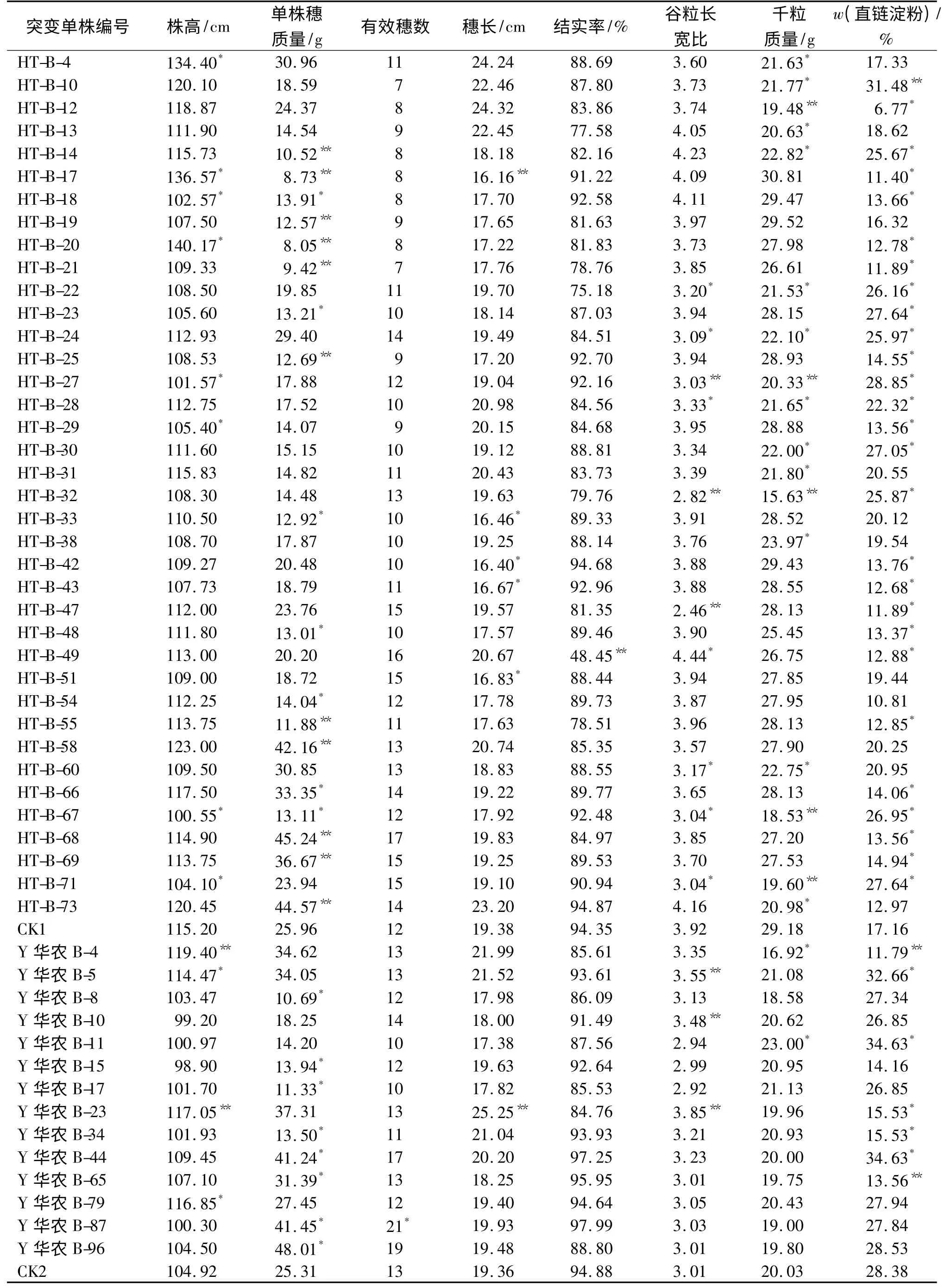
表6 诱变SP3代突变体农艺、经济性状和直链淀粉质量分数情况1)Tab.6 The main agronomic and economic traits and amylose contents of SP3mutation progeny
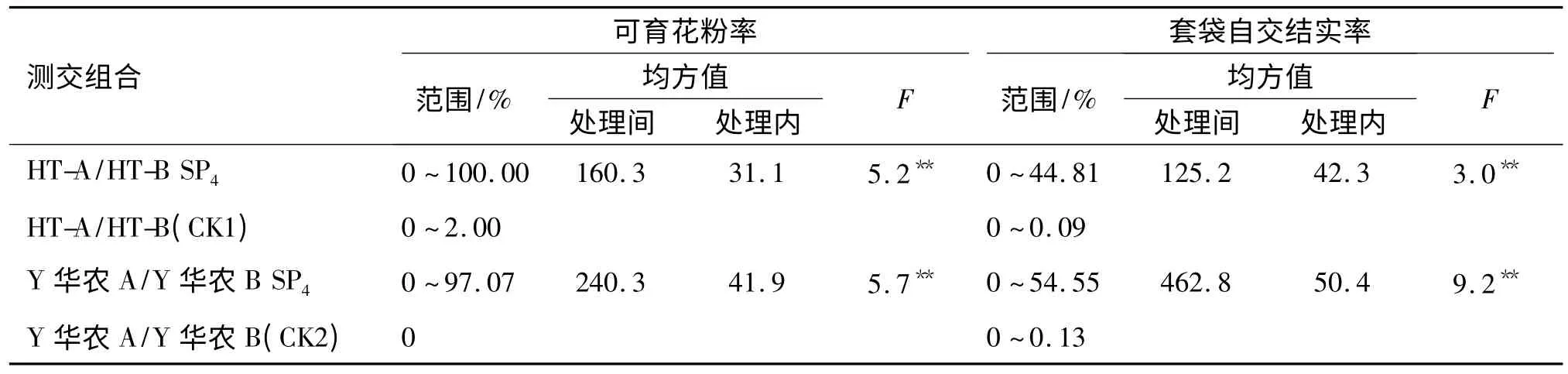
表7 突变株测交F1可育花粉率和套袋自交结实率的方差分析1)Tab.7 Variance analysis of pollen fertility rate and the seed setting rate of test cross F1of mutation progeny
HT-B 与对应不育系HT-A 测交F1可育花粉率变幅为0~100%,套袋自交结实率变幅为0~44.81%,其中HT-B-23 突变株系测交F1代花粉可育率为100%,表现出对不育系完全恢复的特性,HT-B-61 单株套袋自交结实率最高(图1a).49 个突变株系测交F1中,有14 个表现为正常可育,6 个表现为半不育,3个表现为高度不育,26 个表现为全不育.Y 华农B 与对应不育系测交F1代花粉可育率变幅为0~97.07%,套袋自交结实率变幅为0~54.55%,其中3 个株系测交F1代可育花粉率达到90%以上,Y 华农B-20 突变株测交F1代花粉可育率最高,套袋自交结实率也达到30.6%(图1b).70 个突变株系测交后代中,有18 个表现为正常可育,6 个表现为低不育,5 个表现为半不育,3 个表现为高不育,38 个表现为全不育.
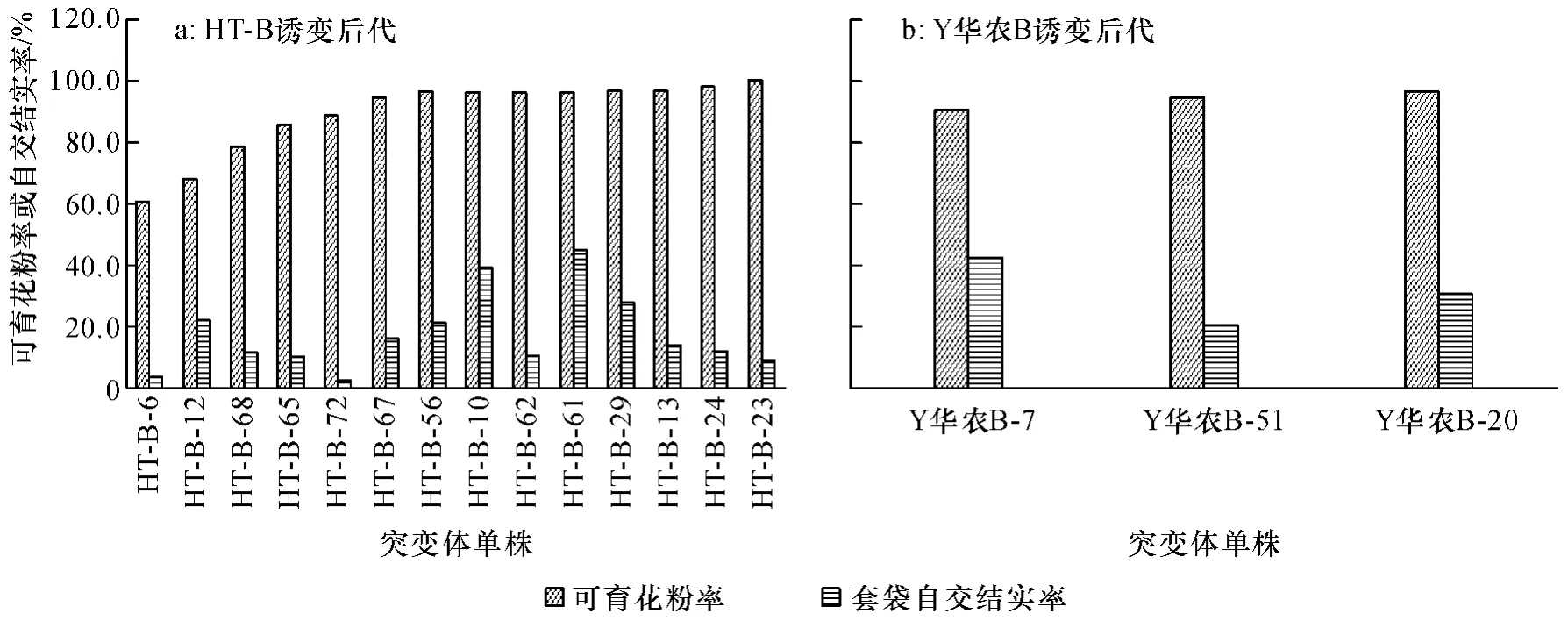
图1 HT-B 和Y 华农B 诱变后代突变体测交F1代花粉育性和套袋自交结实率变化Fig.1 The pollen fertility and seed-setting rate changes of test cross F1generation of HT-B and Y Huanong B mutation progeny
2.5 突变体新种质筛选
经过连续4 代跟踪选择,结合农艺经济性状、品质性状以及测交后代恢保关系的变化,HT-B 诱变SP4代26 个测交F1代表现为全不育突变体中,直链淀粉含量低于对照并且达到差异显著水平的有9株,其中HT-B-9、HT-B-17 和HT-B-70 的直链淀粉质量分数分别为5.3%、11.4%和11.4%,为低直链淀粉含量突变体,而其他农艺经济性状变异不显著,可作为选育低直链淀粉含量不育系加以利用.Y 华农B 诱变后代38 个测交F1代表现为全不育的突变体中,Y 华农B-4、Y 华农B-23、Y 华农B-50 和Y 华农B-65 的直链淀粉质量分数分别为11.79%、15.53%、14.16%和13.56%,明显低于对照,这对于改良Y 华农A 直链淀粉含量偏高具有重要的利用价值.在HT-B 诱变后代测交F1花粉育性表现为完全可育(>85%)突变体中,有HT-B-23、HT-B-29、HT-B-34、HT-B-56、HT-B-61、HT-B-62 和HT-B-72 共7 个突变体直链淀粉含量明显低于对照,且达到差异显著水平,可以为直链淀粉含量恢复系选育加以利用.同时,在2 个材料中也筛选出多个高直链淀粉含量突变体,这对于选育特殊用途稻米杂交稻新品种同样具有较好的应用价值.
3 讨论与结论
本研究结果表明,通过航天搭载2 个水稻保持系材料,SP1代在性状表型上不分离,而种子的发芽率、芽长、苗高和结实率均表现出一定程度的生理损伤,性状间损伤程度差异较大,不同材料不同性状之间的损伤程度不尽相同.SP2代在各农艺经济性状上发生了较大的分离,呈现明显的正负向变异.前人研究表明,诱变SP2代在株高、千粒质量、结实率、谷粒长宽比、单株穗质量和有效穗数变异效果明显[35].本研究发现,2 个材料在诱变SP2、SP3代出现了较为丰富的分离类型,在株高、穗数、千粒质量、谷粒长宽比、单株穗质量、有效穗数等性状上都发生了明显的变异,从中可筛选出目标性状较好的突变株系.但结实率在2 个世代中都变异不明显,这和前人研究结果有所不同,预示着空间环境因素对不同材料的诱变效果不一样.
直链淀粉含量(w)是稻米品质中最重要的理化指标,目前,影响我国籼稻,尤其是杂交水稻稻米品质的因素之一就是直链淀粉含量过高[36-38].鲍正发等[39]的研究指出,籼稻品种9311 经空间诱变后,SP2代单株直链淀粉含量出现了高、低、糯3 种类型的突变.本研究2 个材料经空间诱变后,SP2代单株出现了丰富的直链淀粉变异,变异幅度大,与对照相比达到显著和极显著差异的比例分别达到了32.88%和51.04%,在HT-B 诱变突变单株中,出现了直链淀粉含量极低(w 为1.66%)的个体,表现为糯稻性质,而且SP3代品质性状变异趋势与SP2代具有较强的对应关系.同时,在2 个材料诱变后代中也出现了直链淀粉含量极高的突变个体,这为稻米特殊用途材料的选育提供了选择基础.
当前三系不育系的选育大都采用多代回交选育的办法,耗时太长,且受恢保关系的限制.本研究发现,突变体测交后代花粉育性和套袋自交结实率与对照差异明显.筛选出了多个农艺经济性状变异不显著,而直链淀粉含量降低且保持了对不育系的育性保持能力的突变体,如HT-B-9、HT-B-17、HT-B-70和Y 华农B-4、Y 华农B-23、Y 华农B-50、Y 华农B-65 等,这对于选育低直链淀粉含量不育系具有重要的利用价值.研究表明经航天诱变后,可以有效地诱导三系杂交稻保持系发生多个性状变异,结合农艺经济性状和品质性状以及测交F1代花粉育性表现,可以获得优良的保持系新种质,从而改良原亲本不育系的某些性状,培育新的保持系和不育系,缩短育种年限.
[1]郑家团,谢华安,王乌齐,等.水稻航天诱变育种研究进展与应用前景[J].分子植物育种,2003,1(3):367-371.
[2]陈志强,郭涛,刘永柱,等.水稻航天育种研究进展与展望[J].华南农业大学学报,2009,30(1):1-5.
[3]刘录祥,郭会君,赵林姝,等.我国作物航天育种20年的基本成就与展望[J].核农学报,2007,21(6):589-592,601.
[4]陈芳远,卢升安,易小平.空间诱变创造克服籼粳杂种半不育性新种质和新恢源研究[J].激光生物学报,2000,9(2):119-124.
[5]王慧,张建国,陈志强.航天育种优良水稻品种华航一号[J].中国稻米,2003(6):18.
[6]严文潮,徐建龙,俞法明,等.不同早籼基因型水稻的空间诱变效应研究[J].核农学报,2004,18(3):174-178.
[7]郭涛,蔡金洋,王慧,等.水稻空间诱变SP2代品质性状变异分析[J].华南农业大学学报,2007,28(1):6-9.
[8]王慧,张书涛,郭涛,等.籼型矮秆突变体CHA-2 的矮生性状遗传分析及基因初步定位[J].分子植物育种,2006,4(S2):1-6.
[9]王慧,陈志强,杨祁云,等.水稻空间诱变突变品系主要农艺经济性状及稻瘟病抗性变异[J].华南农业大学学报,2004,25(4):1-5.
[10]王俊敏,魏力军,骆荣挺,等.航天技术在水稻诱变育种中的应用研究[J].核农学报,2004,18(4):252-256.
[11]梅曼彤.空间诱变研究的进展[J].空间科学学报,1996,16(增刊):148-152.
[12]王慧,陈志强,张建国.水稻卫星搭载突变性状考察和品系选育[J].华南农业大学学报,2003,24(4):5-8.
[13]张广彬,于良斌.水稻空间诱变育种研究[J].黑龙江农业科学,1997(4):1-4.
[14]沈桂芳,孙永成,钦天均.中国航天育种搭载试验综述[G]∥信乃诠.航天育种高层论坛论文选编.北京:中国高科技产业研究会,2005:1-6.
[15]吴殿星,舒庆尧,夏英武,等.空间技术诱发的水稻变异及育种价值[J].核农学报,1999,13(3):175-178.
[16]余红兵,周峰,姚涓,等.高空气球搭载水稻种子后代变异的研究[J].核农学报,2004,18(4):276-279.
[17]王呈祥,白志良,王良群.航天育种:我国农业科技革命的新路[J].山西农业科学,2003,31(3):92-96.
[18]余红兵,周峰,姚涓,等.高空气球搭载诱导水稻后代形态变异研究[J].江苏农业科学,2005(1):21-22.
[19]张瑞祥,廖家槐,张红林,等.中国水稻杂种优势利用现状及对策[J].江西农业大学学报,1998,20(2):223-226.
[20]段世华,毛加宁,朱英国.利用RAPD 分子标记对我国杂交水稻主要恢复系的DNA 的多态性研究[J].武汉大学学报:理学版,2001,47(4):508-512.
[21]程式华,闵绍楷.中国水稻品种:现状与展望[J].中国稻米,2000(1):13-16.
[22]王三良,许可.我国籼型杂交水稻育种现状,问题与对策[J].杂交水稻,1996(3):1-4.
[23]陆贤军,钟永模.四川杂交水稻系谱分析[J].西南农业学报,1995,8(水稻专辑):60-67.
[24]欧阳乐军,郭建夫.三系水稻保持系航天育种的研究[J].安徽农业科学,2008,36(27):11855-11858.
[25]王晓玲,余传元,雷建国,等.航天诱变改良水稻不育系研究新进展[C]∥2011年航天工程育种论坛论文集.北京:中国空间技术研究院,2011:82.
[26]吕兑财,黄增信,赵亚丽,等.实践八号育种卫星搭载植物种子的空间辐射剂量分析[J].核农学报,2008,22(1):5-8.
[27]国家高技术航天领域专家委员会.空间诱变育种:农作物育种的新途径[M].北京:国家高技术航天领域专家委员会,2000.
[28]申宗坦.作物育种学实验[M].北京:中国农业出版社,1998:1-4.
[29]严文潮,徐建龙,俞法明,等.不同早籼基因型水稻的空间诱变效应研究[J].核农学报,2004,18(3):174-178.
[30]应存山.中国稻种资源[M].北京:中国农业科技出版社,1993:135-147,541-549.
[31]朱英国,梅启明,涂金洪.不同类型水稻雄性不育系的恢保关系研究[J].遗传,1984,6(5):14-16.
[32]范树国,梁承邺.水稻体细胞无性系R-1、R-2 代中的雄性育性变异观察[J].遗传学报,1995,22(4):293-301.
[33]莫惠栋.我国稻米品质的改良[J].中国农业科学,1993,26(4):8-14.
[34]盖钧镒.试验统计方法[M].北京:中国农业出版社,2000:75-98,193-209.
[35]吴德志,刘永柱,郭涛,等.实践八号育种卫星搭载籼稻的诱变效应研究[J].核农学报2010,24(2):209-213.
[36]黄华康,张卫清,林强.籼型杂交稻米质现状及其遗传改良[J].福建农林大学学报:自然科学版,2002,31(2):155-159.
[37]陈建伟,吴东辉,陈文丰,等.广东省新育成水稻品种米质分析[J].广东农业科学,2004(5):19-20.
[38]韦汉西,邹优柱,何少美,等.高直链淀粉含量早籼组合优Ⅰ315 的选育与应用[J].广西农业生物科学,2007,26(3):240-242.
[39]鲍正发,段智英,赵海军,等.空间诱变引起水稻9311的品质变异[J].核农学报,2004,18(4):272-275.
猜你喜欢
园艺与种苗(2021年10期)2021-11-28 23:31:51
吉林畜牧兽医(2021年6期)2021-08-06 04:16:32
食品工业(2020年8期)2020-08-25 08:23:28
中国粮油学报(2019年4期)2019-07-12 09:06:32
中国种业(2018年8期)2018-08-28 06:40:46
中国塑料(2016年2期)2016-06-15 20:29:57
大学生(2015年1期)2015-05-30 10:48:04
种子科技(2015年8期)2015-05-24 16:14:56
安徽农业科学(2015年13期)2015-01-13 06:20:26
食品科学(2013年22期)2013-03-11 18:29:45
