
Environment controls on Mesoproterozoic thrombolite morphogenesis: A case study from the North China Platform
2013-08-22 07:44TangDongjieShiXiaoyingJiangGanqingPeiYunpengZhangWenhao
Journal of Palaeogeography 2013年3期
Tang Dongjie , Shi Xiaoying , , Jiang Ganqing, Pei Yunpeng, Zhang Wenhao
1. State Key Laboratory of Biological and Environmental Geology, China University of Geosciences (Beijing),Beijing 100083, China
2. School of Earth Science and Resources, China University of Geosciences (Beijing), Beijing 100083, China
3. Department of Geoscience, University of Nevada, Las Vegas, NV 89154-4010, USA
Abstract Thrombolites are widespread in the Mesoproterozoic Wumishan Formation in the North China Platform. This study shows that they mainly concentrated in subtidal carbonate facies with relatively low hydrodynamic conditions, rather than in intertidal zone as suggested previously. From the deep lower to the shallow upper subtidal facies, the thrombolites show evident changes in morphology from dominantly domal to tabular forms, likely suggestive of environmental controls on their morphogenesis and distribution. As the most important component in thrombolites, mesoclots typically consist of organic-rich micritic nuclei and organic-poor fibrous aragonite rims. Mesoclots may vary considerably in their morphology,but a type of specifically shaped mesoclots tends to concentrate predominantly in a particular group of thrombolites. The proportion of the fibrous aragonite rims in mesoclots decrease as the depositional environments become shallower, likely suggesting that the environmental changes also have controls on the internal fabrics of thrombolites. Putative filamentous bacterial colonies are well preserved in some aragonite fans in the matrix between mesoclots,invoking rapid precipitation and aragonite-supersaturated conditions in the ambient waters. It seems that a suboxic to anoxic environment, highly alkaline seawater and relatively low hydrodynamic conditions were among the important factors that facilitated the development and preservation of thrombolites in the Mesoproterozoic epeiric sea on the North China Platform.
Key words mesoclots, fibrous aragonite, bacterial sulfate reduction, organo-mineralization, carbonate supersaturation
1 lntroduction*
Thrombolites are a kind of bio-sedimentary structure formed through interactions of microbes and sedimentation, and constitute a major group of microbialites(Kennard and James, 1986; Aitken and Narbonne, 1989;Riding, 1999, 2000, 2006), in addition to stromatolites,oncolites and leiolites (Ridinget al., 1991; Riding, 1999,2002; Altermann, 2008). They are often associated with stromatolites in the geological record, but differ from the latter markedly in their clotted rather than stratified internal fabrics (e.g., Aitken, 1967; Kennard and James, 1986;Burne and Moore, 1987; Kennardet al., 1989; Ridinget al., 1991; Riding, 2002, 2006, 2008).
Chronologically, thrombolites abound in carbonate successions of the Late Neoproterozoic to Early Ordovician times, and often occur as a major component in build-ups(e.g., Aitken and Narbonne, 1989; Kennard, 1989, 1994;Feldman and Mckenzie, 1998; McCormick and Grotzinger,2001; Grotzinger and James, 2000; Adamset al., 2005;Hicks and Rowland, 2009; Ravioloet al., 2010). Since the Early Ordovician, thrombolites declined sharply due to the diversification of metazoans (Walter and Heys, 1985; Sheehan and Harris, 2004; Riding, 2006) and became restricted largely to highly stressed environments or the aftermaths of mass extinctions. For instance, thrombolites have been well documented in strata of the latest Ordovician to earliest Silurian age (e.g., Sheehan, 2001; Sheehan and Harris, 2004),latest Devonian (e.g., Whalenet al., 2002), Early Triassic(Grotzinger and Knoll, 1995; Ezakiet al., 2003, 2008, 2012;Kershawet al., 2007, 2011; Yanget al., 2011) and in the Miocene interval of the Messinian Salinity Event (Feldmann and McKenzie, 1997). During the Mesozoic, thrombolites seem to have had a resurgence, especially in the Late Jurassic, when they often constituted a prominent portion of reefs(Olivieret al., 2003; Manciniet al., 2004; Helm and Schulke, 2006; Matyszkiewiczet al., 2006), and were able to form noticeable oil reservoirs in marine settings (e.g., Leinfelder and Schmid, 2000; Manciniet al., 2006). It is worth noting that thrombolites are rare in strata prior to the Late Neoproterozoic, with only a few examples known from the Paleoproterozoic of Canada, which may represent the earliest records of thrombolites, although they were commonly regarded as inorganic in origin (Kah and Grotzinger, 1992).To date, thrombolites are virtually unknown from the Mesoproterozoic successions (Grotzinger and James, 2000; Riding, 2000, 2008; Harwood and Sumner, 2011), except for those from the North China Platform (Tanget al., 2013a).
Ecologically, modern thrombolites are known from a variety of environments, such as the restricted hypersaline embayment at Shark Bay, Australia (e.g., Riding, 2000;Jahnert and Collins, 2011, 2012), an open subtidal setting in the Bahamas (e.g., Feldmann and McKenzie, 1998; Planavsky and Ginsburg, 2009; Myshrallet al., 2010; Mobberleyet al., 2012), freshwaters (e.g., Ferriset al., 1997;Lavalet al., 2000; Gischleret al., 2008, 2011), as well as hypersaline (e.g., Puckettet al., 2011) and alkaline lakes(e.g., Kempe and Kazmierczak, 1993; Arpet al., 2003),with a few examples also reported from hot spring settings(e.g., Campbellet al., 2008). In comparison with stromatolites, however, far less attention has been paid to thrombolites, partly due to the presence of many transitional forms between the two that have obscured a conclusive separation, and partly due to the relative restriction of thrombolites in both temporal-spatial distribution and abundance.In many cases, therefore, thrombolites have been studied under the umbrella term of microbialites (Riding, 2000,2006; Sheehan and Harris, 2004; Kershawet al., 2011).
As to the genesis of thrombolites, opinions are controversial. A common idea is that thrombolites developed since the Late Neoproterozoic are biogenic, with their mesoclots (Shapiro, 2000) probably derived from the calcification of microbial colonies in sediments (Kennard and James, 1986; Kennard, 1989; Myshrallet al., 2010).However, earlier thrombolites from the Paleoproterozoic,such as those from the Rocknest Formation, Canada, were thought to have resulted mainly from abiotic processes(Kah and Grotzinger, 1992; Grotzinger and James, 2000),due to the high proportion of fibrous aragonite cement in their interior, which was commonly interpreted as a kind of sea-floor precipitation (Kah and Grotzinger, 1992). In regard to the morphogenesis of thrombolites, opinions are also divergent. For instance, some researchers suggest that they may have resulted from the destruction of stromatolites either through diagenetic alteration or metazoan bioturbation (e.g., Hofmann, 1973; Walter and Heys,1985), or from a high proportion of microbial eukaryotes involved (e.g., Feldmann and McKenzie, 1998) and therefore reflect different microbial metabolisms (Myshrallet al., 2010; Harwood and Sumner, 2011). Whereas some others propose that they may have resulted from taphonomic factors (Turneret al., 2000; Planavsky and Ginsburg, 2009). Nevertheless, most researchers agree that thrombolites represent a distinct type of microbialites and are generated by microbial communities and their interactions with the environments where they have lived (e.g.,Kennard, 1989; Shapiro, 2000; Riding, 2002, 2006; Tanget al., 2013a).
Thrombolites from the Mesoproterozoic Wumishan Formation (ca 1.5-1.45 Ga) of the North China Platform(NCP) (Figs. 1A, 1B) have been recently studied by Tanget al. (2013a), with a focus on their biogenicity and organomineralization, mainly from the angle of ultra-fabrics and constituent organominerals. In this paper, based on their macro- to microscopic texture variations and distribution in relationship to depositional environments, we primarily focus on mesoclot genesis, possible environmental controls on their morphogenesis, as well as their palaeoecologic constraints.
2 Geological setting
The NCP witnessed a tectonic evolution from the breaking-up of the Columbia supercontinent to the assembly of the Rodinia supercontinent. During Proterozoic times, it received a sedimentary succession of up to ~10 km in maximum thickness in the central part of the platform (Chenet al., 1999; Wanget al., 2000; Chenget al., 2009; Tanget al., 2013a). The Mesoproterozoic (1.6-1.0 Ga) rocks in the central platform comprise two groups, with the Jixian Group, characterized primarily by carbonates, in the lower part and the Qingbaikou Group, dominated by siliciclastics, in the upper part (Fig. 1A). The Jixian Group is bounded disconformably both at base and top, and includes five formations that constitute two large sedimentary cycles (Fig. 1A). The lower cycle consists of the Gaoyuzhuang Formation of dark-grey carbonates in the lower part and the Yangzhuang Formation of light dolostone alternating with purplish siltstone in the upper part, showing an overall tendency of shallowing-upward from subtidal to supratidal facies. The upper cycle comprises three formations (Fig. 1A). The Wumishan Formation, one of the most widespread lithostratigraphic units in the Mesoproterozoic of the NCP, is dominated by peritidal carbonates, locally rich in thin chert bands, and contains a variety of micro-bialites. The overlying Hongshuizhuang Formation in the middle is characterized by dark shale, with some thin siltstone interbeds. The Tieling Formation at the top consists primarily of light carbonates with well developed columnar stromatolites that often form dune-shaped build-ups.Three formations are recognized in the Qingbaikou Group(Fig. 1A). The Xiamaling Formation is dominated by dark shale, the Changlongshan Formation is characterized by quartz sandstone, and the Jingeryu Formation consists mainly of carbonates. Three significant disconformities,respectively, at the base of the Xiamaling Formation and the Changlongshan Formation, and on top of the Jingeryu Formation (Fig. 1A), are widely recognizable across the platform, representing long intervals of non-deposition.

Fig. 1 Proterozoic succession and simplified palaeogeographic map of the study area. A-Revised Proterozoic succession for the NCP with key age constraints (ages adopted from Lu and Li, 1991; Gao et al., 2007, 2008a, 2009; Qiao et al., 2007; Su et al., 2008, 2010;Li et al., 2010); B-Location of the study area; C-Simplified palaeogeographic map of the study area (after Qiao and Gao, 2007) and location of the study sections.
In recent years, a number of zircon SHRIMP U-Pb datings (Fig. 1A) have been obtained from the Mesoproterozoic succession of the NCP. The Lower Xiamaling Formation is dated as 1368±12 Ma (Gaoet al., 2007,2008b) and 1372±18 Ma (Suet al., 2010), the Middle Tieling Formation as 1437±21 Ma (Suet al., 2010), the Upper Gaoyuzhuang Formation as 1560 Ma (Liet al., 2010),and the Upper Dahongyu Formation as 1625 Ma (Lu and Li, 1991; Gaoet al., 2008a). Therefore, ages for the top and basal boundaries of the Gaoyuzhuang Formation can be reasonably estimated at 1.53 Ga and 1.60 Ga, respectively; and the basal boundary of the Xiamaling Formation is set at 1.40 Ga (Qiaoet al., 2007; Gaoet al., 2009). To date, no direct dating has been obtained from the Wumishan Formation, according to known ages and stratigraphic constraints, however, this formation can be approximately inferred to have been deposited between 1.50 Ga and 1.45 Ga (Gaoet al., 2009; Tanget al., 2013a), belonging to the Calymmian System of the Early Mesoproterozoic (Ogget al., 2008; ICS, 2012).
The Wumishan Formation is about 3340 m thick in Jixian County, Tianjin, where it has conformable contacts with the overlying and underlying strata. In the Yesanpo area, Hebei Province (Fig. 1C), the formation is reduced to 1800 m in thickness, underlain disconformably by the Gaoyuzhuang Formation, but overlain conformably by the Xiamaling Formation. In the Huailai area, Hebei Province(Fig. 1C), it further decreases to 1400 m, and has disconformable contacts with both overlying and underlying strata. Lithologically, sedimentation of the Wumishan Formation remains relatively constant across the platform, and is commonly dominated by dolostones of peritidal facies with diversified microbialites, including thrombolites, biolaminites, stromatolites, dendrolites, and various microbially induced sedimentary structures (MISS; Shiet al., 2008;Shi and Jiang, 2011). In the study area (Fig. 1B, 1C), the Wumishan Formation comprises four members. The basal member is dominated by oolite-bearing, stromatolitic and micritic dolostones. The lower member consists mainly of thrombolitic and laminated dolostones. The middle member is largely composed of thick-bedded thrombolitic and layered stromatolitic dolostones, with some domal microbialites. The upper member is typically made up of micritic and finely laminated dolostones, with occasional thin layers of thrombolitic dolostone in its lower part.
3 Sedimentary facies and thrombolite distribution
In the study area (Fig. 1B, 1C), the Wumishan Formation comprises hundreds of upward-shallowing parasequences that further stack into depositional cycles of various orders. In general, four types of parasequences can be distinguished according to their sedimentary features and internal architectures. Each of the parasequences is primarily made up of 2-3 kinds of Basic Depositional Units(BDUs, Unit a-c; Fig. 2; Table 1). Although these BDUs may vary considerably in thickness and relative proportion in a parasequence, depending on their particular depositional environments in places, their architectures and stacking patterns in most parasequences are similar (Tanget al., 2011, 2013a). The characteristics of the four parasequence types and their constituent units can be summarized as follows (Table 1; Fig. 2).
Type-I parasequences often occur in the upper part of a depositional sequence, and may have been deposited in upper subtidal to supratidal environments (Fig. 2).Type-I parasequences commonly consist of three BDUs(I-a, I-b and I-c). The basal unit (I-a) is characterized by thinly layered thrombolites of 0.1-0.8 m thick (Fig. 3A),occasionally with fine intraclasts or ooids. Mesoclots in the thrombolites are commonly small, elliptical to irregularly patchy in shape. In places, Unit I-a may change into dolorudite (Figs. 3B, 3C) or oolitic dolostone (Fig. 3D),often with herringbone cross-bedding (Fig. 3C), indicating a highly dynamic environment of upper subtidal to lower intertidal (Zhouet al., 2006; Tanget al., 2011). It is worth noting that mesoclots and intraclasts rarely coexist in unit I-a, and thrombolites contain only a minor amount of fine sand, whereas dolorudites contain virtually no mesoclots. The middle unit (I-b) is dominated by micritic dolostone and rich in silicified microbial mats that are often expressed as chert bands in vertical section (Fig. 3E). Trough cross-bedding (Fig. 3F) and mat-protected ripple marks (a kind of MISS) are common in this unit, suggestive of a low intertidal environment. The upper unit (I-c) is commonly composed of micritic, or occasionally argillaceous dolostone, often with mud-cracks,halite pseudomorphs (Fig. 3G), siliceous crusts and mat shrinkage cracks with curled margins (a kind of MISS;Fig. 3H; Shi and Jiang, 2011), suggestive of a supratidal environment with strong evaporation (Tanget al., 2011).

Table 1 Characteristics of the four major parasequence types of the Wumishan Formation and their constituent depositional units in the study area
Type-II parasequences also consist of three BDUs (II-a,II-b and II-c), and primarily developed in upper subtidal to lower supratidal environments, with the basal unit (II-a)slightly deeper than that (I-a) of the type-I parasequence(Fig. 2B). In a type-II parasequence, Unit II-a is mainly made up of thick-bedded thrombolites (1-2 m thick).Mesoclots in the thrombolite often exist as curved strips or ribbons, producing a distinctive fingerprint-like pattern of the thrombolite on weathered surfaces (Fig. 4A), or occur as tongue-shaped to irregular patches. The general lack of cross-bedding and swash marks and occasional presence of intraclasts (Fig. 4B) in this unit likely indicate a subtidal environment of low-energy that may be occassionally influenced by strong waves. Unit II-b is dominated by micritic dolomite rich in microbial mats and chert bands, bearing some fine intraclasts, while unit II-c consists primarily of interbedded light micritic dolostone (Fig. 4C). Unit II-b is interpreted as mainly an intertidal deposit, and the unit II-c as a supratidal deposit.
Type-III parasequences usually comprise two BDUs(III-a and III-b), and commonly lacks the supratidal unit.Compared with those in the type-II parasequence, mesoclots in Unit III-a often appear as irregular patches (Fig.4D, 4E). Furthermore, Unit III-b is richer in microbial mats, often bearing small domal stromatolites and flatpebble-like intraclasts (Fig. 4F). These features likely indicate that the type-III parasequence was mainly formed in lower subtidal to lower intertidal environments, deeper than the type-II parasequence (Fig. 2B).
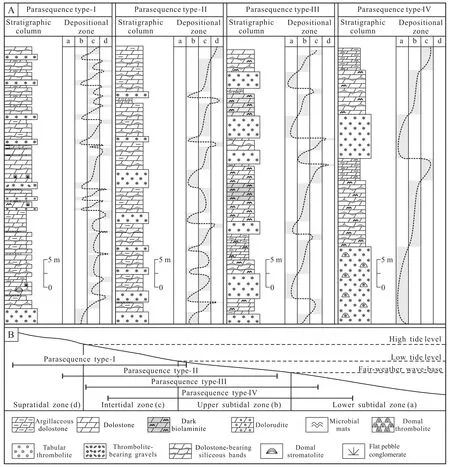
Fig. 2 Major parasequence types in the Wumishan Formation and their suggested distribution model. A-Common parasequence types; B-Suggested distribution of the four types of parasequences: type-I from uppermost subtidal to supratidal facies (low part of the fourth Member); type-II from upper subtidal to supratidal facies (top of the third Member); type-III from shallow lower subtidal to intertidal facies (middle part of the third Member); and type-IV from deep subtidal to lower intertidal facies (low part of the third Member).
Type-IV parasequences consist of two BDUs (IV-a and IV-b) (Fig. 2A), and may have been deposited in the lower subtidal to intertidal zone, deeper than the type-III parasequence (Fig. 2B). In the type-IV parasequence, Unit IV-a is commonly dominated by massive or domal thrombolites in which mesoclots are often vertically arranged,more or less in rows (Figs. 5A, 5B) or in branches (e.g., in dendroid forms) (Fig. 5C). Occasionally, variable thrombolite build-ups containing abundant infilled voids can be observed in this unit (Figs. 5D, 5E). Overall, Unit IV-a has the greatest thickness among the basal units in each of the four types of parasequences, and a single bed may attain a maximum thickness of 12 m. No swash or any other dis-turbed structures or intraclasts have been observed in this unit, suggesting that Unit IV-a was most likely formed in the quiet and deeper environment of the low subtidal zone.Unit IV-b consists primarily of micritic dolostone, with abundant silicified microbial mat layers and some domal stromatolites, possibly indicating upper subtidal to lower intertidal environments.

Fig. 3 Major sedimentary facies in the parasequence type-I from the Wumishan Formation. A-Domal thrombolites in Unit I-a;B-Dolorudite in Unit I-a; C-Dolorudite with herringbone cross-bedding in Unit I-a; D-Purplish oolitic dolostone in Unit I-a; EDolomicrite composed of alternating silicified mats (dark) and carbonate precipitate layers (light) in Unit I-b; F-Trough cross-bedding in Unit I-b; G-Halite pseudomorphs in Unit I-c; H-Silicified mat shrinkage cracks with curled margins in Unit I-c. Fig. D, G, and H are in plan view, others are perpendicular to bedding.

Fig. 4 Common sedimentary facies in parasequences type-II and type-III of the Wumishan Formation. A-Thrombolite with ribbon- or stripe-like mesoclots showing fingerprint-patterns in Unit II-a (view perpendicular to bedding); B-Flat-pebble-like intraclast in thrombolite (view perpendicular to bedding); C-Light gray dolomicrite in Unit II-c; D-Tabular thrombolite with irregular blocky mesoclots in Unit III-a (plan view); E-Tabular thrombolite with irregular blocky and patchy mesoclots in Unit III-a; F-Imbricated silicified mat chips in Unit III-b.
Overall, from type-I to type-IV, the thickness and proportion of Unit-a in each of the parasequence types increases, while those of Unit-c are gradually reduced and become completely absent in the type-IV parasequence(Fig. 2A). These variations likely indicate a gradual deep-ening of the environment in which the parasequences were deposited, and probably also imply that distribution of the thrombolites and, to some extent, their morphogenesis,were largely controlled by environmental factors related to water-depth.

Fig. 5 Major facies in the parasequence type-IV of the Wumishan Formation. A-Thrombolite with branches formed by vertically lined mesoclots; B-Enlarged view of the boxed area in Fig. A, showing branches formed by vertically arranged mesoclots; C-Mesoclots arranged in a dendroid pattern in a domal thrombolite; D-Dendroid columns in a large thrombolite reef; E-Thrombolite with irregularly branching mesoclots and infilled voids in a microbial reef.
Based on sedimentary characteristics and regular changes in the proportions of BDUs in various parasequence types, a facies distribution model for these parasequence types is suggested (Fig. 6). It is worth noting that domal thrombolites primarily occur in lower subtidal environments, tabular thrombolites with blocky mesoclots are mainly concentrated in lower upper subtidal environments, while thrombolites with fingerprint-like fabrics are more often developed in shallow upper subtidal environ-ments. Dolorudite and oolitic dolostone may occur in the upper subtidal to lower intertidal, where herringbone and trough cross-bedding are often observable. Laminated dolostone rich in mat layers and chert bands is commonly developed in the intertidal facies. In contrast, argillaceous micritic dolostone is primarily seen in the supratidal facies, often associated with mat shrinkage structures, halite pseudomorphs, and other evidence of subaerial exposure,indicative of frequent exposure and strong evaporation.
4 Morphogenesis and microtextures of the thrombolites
Thrombolites are abundant in the Wumishan Formation,and primarily exist in the lower units of all parasequence types, largely confined to the subtidal zone. They vary considerably in morphology, but can be broadly grouped as domal and tabular forms. Highly domal thrombolites only occur in the Unit-a of the type-IV parasequence (Fig.5), providing evidence of a deeper subtidal setting (Fig. 6),whereas tabular forms widely exist in Unit-a of all parasequence types (Fig. 4A, 4B, 4D, 4E), suggesting a broader and shallower subtidal setting (Fig. 6).
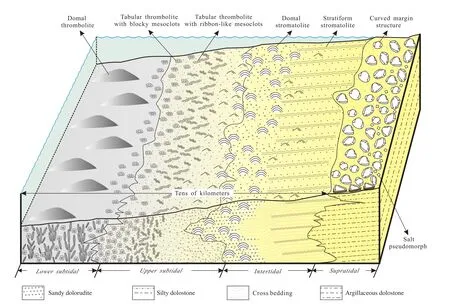
Fig. 6 Onshore-offshore distribution model suggested for the various types of thrombolites. Domal thrombolites with vertically arranged or dendroid mesoclots are primarily developed in the lower subtidal zone. Tabular thrombolites with irregular blocky mesoclots mainly occur in the lower upper subtidal zone; tabular thrombolites with ribbon-like mesoclots, which commonly result in fingerprint patterns on thrombolite surfaces, dominate the middle part of the upper subtidal zone; ooids, sands, and cross-bedding commonly appear in the shallower upper subtidal to lower intertidal zones. Domal stromatolites are mainly distributed in the lower intertidal zone,while stratiform stromatolites dominate the upper intertidal zone. Microbial mat shrinkage structures and salt pseudomorphs are primarily distributed in the supratidal zone.
Three basic components recognized in the thrombolites(Fig. 7) have been discussed by Tanget al. (2013a), we will not restate them herein. Our observation of variably shaped thrombolites in the Wumishan Formation reveals four types of mesoclots with different shapes, which are the most important component of the thrombolites: (1) irregular blocks (Figs. 4D, 4E, 7); (2) curved strips or ribbons (Figs. 4A, 7); (3) irregular columns (Fig. 5A, 5B); and(4) branching columns (Fig. 5C). The blocky mesoclots commonly vary from 0.1 cm to 4 cm in size. Some of them are isolated and surrounded by micritic fillings or sparitic dolomite; some others clump together (Figs. 4D, 4E, 7).The ribbon-like mesoclots are generally 1-2 mm wide and several millimeters to a few centimeters long (Figs. 4A, 7).The irregular column-like mesoclots are usually vertically arranged, but highly variable in size (commonly 5 cm × 1 cm) (Fig. 5A, 5B). The branching column-like mesoclots,often subvertical or at a high-angle to the bedding plane,are generally 5 cm long and 2 cm wide in the middle parts of the columns, and 0.5-1 cm long and 0.2 cm wide in the case of the branches (Fig. 5C).
Each of the mesoclot types tends to concentrate preferentially in a particularly shaped thrombolite. For instance,blocky mesoclots often constitute medium- to thick-bedded tabular thrombolites with irregular internal structures (Figs.4D, 4E, 7), curved ribbon-like mesoclots produce thickbedded thrombolites with fingerprint-like textures in their interior (Fig. 4A), irregular column mesoclots mainly occur in lower domal thrombolites (Fig. 5B), and branching mesoclots show up primarily in massive domal thrombolites(Fig. 5C). In large thrombolite build-ups, variously shaped mesoclots may co-exist and be entangled (Fig. 5D, 5E).
5 Micro-fabrics of the thrombolites

Fig. 7 Polished slab showing internal fabrics and three major components in thrombolites. a-Variably shaped dark mesoclots; b-Gray micrite as inter-framework fillings surrounding mesoclots; c-Voids filled with light sparitic cement. Sample from the Second Member of the Wumishan Formation, Yesanpo, Hebei Province.
Micro-fabrics of the thrombolites have been described by Tanget al. (2013a). Here, we only give a brief sum-mary of their major characteristics and supplement with some newly observed results. Microscopically, mesoclots primarily consist of (1) organic-rich micritic nuclei, and(2) outer rims of fibrous aragonite (Fig. 8A, 8D), which have been variably dolomitized. In some of the nuclei, tiny pyrite grains (Fig. 8E, 8G) are present, and often associated with organic relics, especially in silicified samples(Fig. 8F, 8G). Most of the outer rims appear as small radial fans or isopachous layers of fibrous aragonite (Fig.8A, 8B), and some of them may also develop multi-layer structures formed by alternating organic-rich micritic layers and sparitic cement (Fig. 8E), providing evidence of multi-stage growth of the outer rims.
Organic remains are commonly present in mesoclots.In some silicified thrombolite samples, putative bacterial filaments and closely associated relics of extracellular polymeric substances (EPS) (Fig. 9A-9D), which are partially replaced by nanoglobules (Fig. 9C, 9D) and other organic remains (Fig. 9E), are clearly recognizable in the nuclei. The EPS relics often appear as translucent films covering tiny crystals (Fig. 9B-9D). In some well-preserved samples, abundant spheroids, generally 10-15 μm in diameter, are observed, comprising numerous coccoids about 1 μm in diameter (Fig. 9F). The coccoids, closely associated with EPS relics (Fig. 9G), likely derive from mineralized bacteria cells, and the spheroids are possibly silicified bacteria colonies (Tanget al., 2013a). Partially degraded filaments, ~1.5 μm in diameter and up to 8 μm in length, also have been observed in the nuclei of mesoclots(Fig. 9H). The filaments are likely mineralized filamentous bacteria relics, similar to those found in some Mesoproterozoic microdigitate stromatolites (Tanget al., 2013b).
The inter-framework fillings within thrombolites comprise mainly micritic to silt-sized dolomite (Figs. 8A, 8B,8F, 10A), with a few fine quartz and some pyrite grains(Fig. 10A, 10B). Occasionally, some small fibrous aragonite (pseudomorph) fans also exist in the matrix of thrombolites. They are commonly 2000 μm high, and tend to connect horizontally to form isopachous layers (Fig. 10C).Crystal fibers in the fans have feathery features and irregular extinctions, suggesting that they were altered from aragonite precursors (Loucks and Folk, 1976; Mazzullo,1980; Sandberg, 1985; Perytet al., 1990; Sumner and Grotzinger, 2000), which were likely originally deposited as sea-floor precipitates.
In some samples, putative bacterial colonies are observable in the fibrous aragonite fans (Fig. 10D). They often appear as clumps, 20-30 μm in size, with putative bacterial filaments about 1 μm in diameter that are entangled together. Needle-like crystal clusters also exist in the matrix (Fig. 10E). The needles seem to have nucleated on organic-rich clumps, then grew radially outward into clusters with spear-like terminations.
6 Discussion
6.1 Origin of micritic nuclei and fibrous aragonite rims in mesoclots
The general lack of intraclasts and terrigenous particles in mesoclots likely indicates that the mesoclots primarily derived from carbonate precipitation induced by bio-chemical processes, rather than from trapping and binding of sediment by microbial activity. The sharp contact between nuclei and rims, and the distinct constituents in each, are indicative of different formation mechanisms. Particularly,the nuclei of mesoclots were most likely derived from the organo-mineralization of microbial cells or their colonies and EPS secreted by them, while rims encasing the nuclei were possibly the result of carbonate precipitation facilitated by interactions between microbes and the surrounding microenvironment (Tanget al., 2013a).
Mesoclots without aragonite rims in thrombolites were once suggested to have formed byin situmineralization of microbial communities (Kennard and James, 1986), but no direct evidence has been provided. In the present study,most of the observed mesoclots are surrounded by fibrous aragonite that has been neomorphosed without recognizable microbial fossils (Fig. 8A, 8B, 8D), while in some earlier silicified mesoclots, which commonly lack fibrous aragonite outer rims, EPS relics and pyrite grains associated with organic remains are observable in the organic-rich nuclei (Fig. 8F, 8G). It is possible that silicification prior to significant decomposition of organic matter enabled the microbial fossils to be preserved in nuclei (Fig. 9F). These phenomena likely support the hypothesis that mesoclots were formed byin situmineralization of microbial communities and imply that the decomposition of organic matter in the nuclei was an important factor in inducing the precipitation of the fibrous aragonite that surrounds the nuclei.
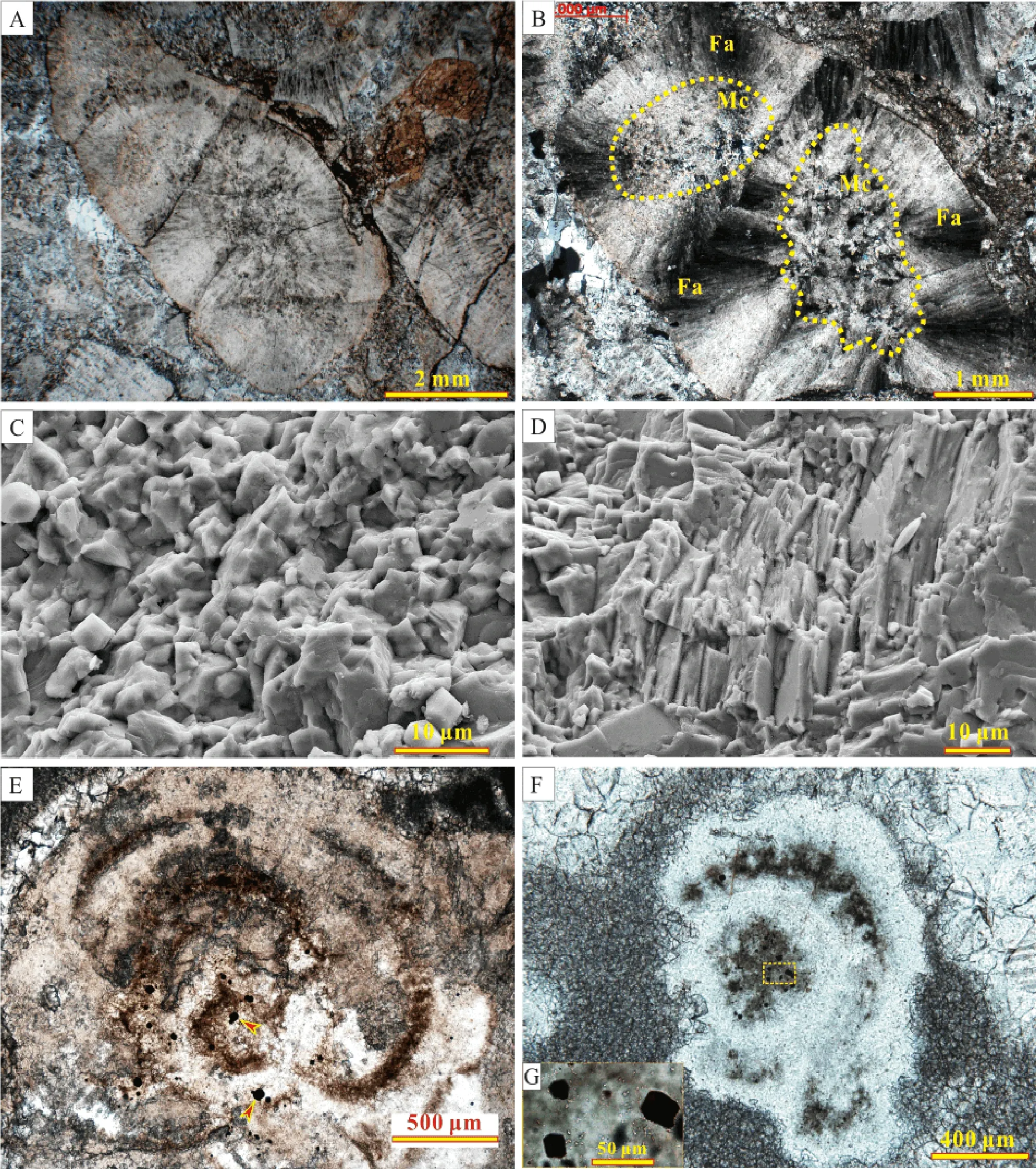
Fig. 8 Micro-textures of mesoclots in the thrombolites. A-Lobate mesoclot composed of an organic-rich micritic nucleus and aragonite outer rim; B-Mesoclot with two micritic nuclei (Mc) and an outer rim formed by fibrous aragonite fans (Fa); C-Micritic particles in the nucleus of a mesoclot; D-Fibrous aragonite pseudomorphs in the outer rim of a mesoclot; E-A mesoclot with multi-layered rim and tiny pyrite grains (denoted by arrows); F-A silicified mesoclot; G-Enlarged view of the boxed area in Fig. F, showing pyrite grains in an organic-rich nucleus. A and E-G in plane-polarized light; B in cross-polarized light; C-D observed using field emission scanning electron microscopy (FESEM).

Fig. 9 Mesoclots and putative bacterial fossils in silicified thrombolites. A-A mesoclot with a silicified micritic nucleus; B-Enlarged view of the boxed area in A, showing putative bacterial filaments (arrows) and closely associated EPS relics as irregular translucent films (arrows) on crystal surfaces; C-FESEM view of the area denoted by arrows in B (rightmost of the three arrows), showing filmlike EPS and closely associated nanoglobules (Ng); D-Another enlarged view of the area denoted with arrows in B (rightmost of the three arrows), showing putative filamentous bacteria (FB), which are partly covered by film-like EPS relics and partially replaced by nanoglobules (Ng); E-Organic remains in the nucleus of a silicified mesoclot; F-Putative bacterial colonies in a silicified mesoclot;G-A fossilized coccoid observed in the putative bacterial colonies shown in F; H-A partially degraded filament found in the nucleus of a mesoclot. A in cross-polarized light; B and E-F in plane polarized light; C-D and G-H, FESEM photos.

Fig. 10 Some other features associated with mesoclots. A-Micritic matrix between mesoclots, mixed with a few fine quartz and pyrite grains; B-A close-up of pyrite grains in the matrix; C-Dolomitized aragonite fans in the matrix within the thrombolite; D-A putative filamentous bacterial colony in a fibrous aragonite fan; the filaments are entangled with small dark clumps; E-Crystal clusters in a void within thrombolites, growing radially outward from organic-rich clumps.
Fibrous aragonites, similar to those developed in the Wumishan thrombolites, are commonly interpreted as sea-floor precipitates and are common in the subtidal carbonate facies of Neoarchean to Lower Paleoproterozoic successions, but were reduced greatly in abundance in the Meso- to Neoproterozoic, and are almost completely absent in Phanerozoic carbonates. Such an obvious decline with time is commonly interpreted as the result of secular changes in seawater chemistry, especially in calcium carbonate saturation (Grotzinger and Kasting, 1993; Sumner and Grotzinger, 1996; Knoll and Semikhatov, 1998; Grotzinger and James, 2000). Three possible factors conducive to fibrous aragonite precipitation have been proposed(Bartleyet al., 2000): (1) an arid environment with strong evaporation, which will facilitate the elevation of calcium carbonate saturation in seawater, (2) decomposition of microbial mats, which can remove the inhibiting factors for carbonate precipitation, and (3) an elevated concentration of HCO3-(largely produced by anaerobic decomposition of organic matter), which will increase water alkalinity and provide material for carbonate precipitation. However,these conditions may also exist in the Phanerozoic marine environments, and aragonite layers and rims encasing mesoclots have rarely been recorded in thrombolites of that period. In the Wumishan Formation, it has been noticed that (1) supratidal carbonates with halite pseudomorphs are devoid of fibrous aragonite, while subtidal thrombolites contain abundant fibrous aragonite, indicating that evaporation may not be a critical factor for aragonite precipitation; (2) most fibrous aragonite primarily initiated from organic-rich nuclei (Fig. 8A, 8B, 8E) or on degraded microbial mats (Fig. 10C), suggesting a close relationship between fibrous aragonite precipitation and decomposition of organic matter; (3) the carbonate saturation under which the thrombolites developed was much higher than that in the modern oceans, implying that the supersaturated carbonate in the microenvironment was likely the result of the anaerobic decomposition of organic matter.
In general, the thrombolites in the Wumishan Forma-tion are distinct from those documented from the Cambrian to Ordovician successions (Pratt and James, 1982;Kennard, 1989; Turneret al., 2000) in having a much higher proportion of aragonite cement (i.e., fibrous aragonite rims) within frameworks. They share similarity with those from the Rocknest Formation (ca 1.9 Ga), Canada,which are also characterized by abundant fibrous aragonite (Kah and Grotzinger, 1992). We suspect that these Proterozoic thrombolites, by having similar microscopic structures and mineral components, are probably close in origin and mineralization mechanism. Although microbial fossils can hardly be identified from the Paleoproterozoic thrombolites due to strong diagenetic alteration, their possible biotic origin and comparable microbial communities to those elsewhere in the Mesoproterozoic would be expected.
6.2 Growth rate and potential source of fibrous aragonite
Fibrous aragonite layers in the thrombolites are commonly composed of crystal fans that can grow as high as 2000 μm (Fig. 10C). The large size of the crystal fans requires a very high crystal growth rate, a very low terrigenous input rate, or a combination of both (Sumner, 2002).If the crystal growth rate was comparable to the estimated rate of 1 mm/40 yr for aragonite cement precipitation in modern reefs (Grammeret al., 1993, 1996), it would take 80 yr for a 2000 μm thick aragonite crystal fan to be formed in the thrombolites. However, it might be unrealistic to deduce no obvious sediment influx that would interrupt aragonite crystal growth for such a long time (Sumner, 2002). Rather, it is more likely that the growth rate of the aragonite crystals in the Mesoproterozoic thrombolites was much faster than that in modern reefs.
The preservation of the filamentous bacteria also provides evidence for a high aragonite growth rate in the thrombolites. In some of the samples, filamentous bacterial colonies of ~30 μm in diameter appear to have been preserved with their original morphology in the fibrous aragonite layers (Fig. 10D). This likely suggests that they were mineralized much earlier before significant decomposition could have taken place, possibly in less than 30 days (Bartleyet al., 2000). Taking this estimate as a reference, a growth rate of 1 μm per day would be expected for the fibrous aragonite layers (30 μm thick on average), so that the bacteria could be completely buried before degradation. If this is true, then the growth rate of aragonite layers in the thrombolites would be approximately 13.6 times higher than in modern reefs (Grammeret al., 1993, 1996).Such rapid precipitation may imply that the saturation with respect to aragonite of the Mesoproterozoic seawater was higher than that of the modern oceans (Grotzinger, 1989;Grotzinger and Kasting, 1993), like that of “Aragonite Seas” as suggested by Hardie (1996, 2003).
If aragonite fan height could be used as a reliable indication of aragonite saturation in seawater, then the development of aragonite fans as high as 1 m in Archean subtidal settings (Sumner and Grotzinger, 2000) would require a much higher aragonite saturation than that in the Mesoproterozoic. Thus, the thrombolites in the Wumishan Formation may have developed in an environment in which aragonite saturation was intermediate between that of the Archean and Phanerozoic.
A high precipitation rate of fibrous aragonite requires a persistent high carbonate saturation of the seawater(Grotzinger, 1989; Grotzinger and Kasting, 1993). In the Wumishan Formation, although degraded microbes and EPS in the nuclei of mesoclots likely initiated carbonate nucleation and precipitation of fibrous aragonite in the outer rims, they alone could hardly sustain the continuous growth of the fibrous aragonite rims. Other material sources might be required for the rapid accretion of fibrous aragonite rims. Organic matter from microbes in ambient water could be a potential complementary source (Woodset al., 1999).
In modern oceans, carbonate precipitation can be facilitated by various microbial groups and their metabolisms (Dupraz and Visscher, 2005; Visscher and Stolz,2005), among which BSR (bacterial sulfate reduction,CH2O+SO42-→HCO3-+HS-+H2O) is commonly regarded as one of the important processes (Visscheret al., 2000;Duprazet al., 2004; Dechoet al., 2005; Dupraz and Visscher, 2005; Baumgartneret al., 2006; Braissantet al.,2007). It has been suggested that the Mesoproterozoic ocean, similar to the modern Black Sea, was permanently stratified, and lower in sulfate concentration (Anbar and Knoll, 2002; Rouxelet al., 2005), with a moderately oxic surface layer, but an anoxic and ferrous deep layer (Canfield, 1998; Anbar and Knoll, 2002), and a possible euxinic wedge in between the two in coastal areas (Liet al.,2010; Planavskyet al., 2011). The chemocline in the ocean was likely around a depth of 25 m (Brockset al, 2005;Tanget al., 2011), and seawater sulfate concentration was about 0.5-2.5 mM (Kahet al., 2004; Brockset al., 2005).According to these estimations, most thrombolites in the Wumishan Formation likely developed in a suboxic to anoxic environment, consistent with an independent study of redox sensitive elements of the formation (Tanget al.,2011), where sulfate-reducing bacterial activity might have prevailed. The presence of acicular gypsum crystals in the matrix of thrombolites (Fig. 10E) likely indicates a moderate sulfate supply to the environment. Therefore,we suspect that suboxic to anoxic conditions and an active BSR process were probably among the major causes that maintained aragonite supersaturation in subtidal environments in the NCP during the Mesoproterozoic.
6.3 Possible environmental controls on the throm‑bolites
In the Wumishan Formation, most thrombolites occur in the subtidal facies (Figs. 2A, 6), providing evidence that water depth is an important control on the development of thrombolites. As accommodation space, hydrodynamics, detrital influx, sulfate concentration, and redox conditions are all closely related to water depth, they may, as important environmental factors, affect the activities and metabolisms of microbial groups, and thus exert influence on the development of thrombolites in various ways.
In terms of dominance, from the lower subtidal to upper subtidal facies, the thrombolites change from domal to tabular forms. This clear tendency suggests that accommodation variations indeed affected the morphology of thrombolites (Kershawet al., 2011). Although the thrombolites occurred in both quiet and agitated environments,the morphology of dominant mesoclots in them varies considerably, likely in response to hydrodynamic conditions. In the intertidal carbonate facies, due to the strong disturbance by currents and waves, tabular and lower dune-shaped stromatolites dominate instead of thrombolites, while in the subtidal facies thrombolites prevail. This likely indicates that although variable and perhaps not directly related to particular groups of microbial builders, a relatively deeper and quieter setting is more favorable for the formation of thrombolites (Jahnert and Collins, 2011,2012). Terrigenous influx apparently restrains the accretion of fibrous aragonite, while the latter is a critical requirement for the preservation of mesoclots. This may also partly explain why thrombolites are rare in siliciclastic sediments.
Salinity and redox conditions may not control the presence of thrombolites directly. They will, however, affect microbial groups and their metabolisms, and thus influence the microstructures and possibly also mineral components in the mesoclots indirectly. The general lack of intraclasts and wide presence of microbial remains in the nuclei of mesoclots likely indicate that, instead of trapping and binding of sediment particles by microbial communities,which is well-documented in modern stromatolites from the Bahamas and Shark Bay (e.g., Feldmann and McKenzie, 1998; Macintyreet al., 2000; Riding, 2000; Reidet al., 2003), organo-mineralization and chemical carbonate precipitation induced by microbial activities are the major mechanisms for the formation of mesoclots.
Fibrous aragonite is rare in the oxic intertidal facies, but often exists in anoxic voids trapped in carbonates, suggesting that redox was probably one of the major controls for its precipitation (Tanget al., 2011). As sulfate-reducing bacteria communities, therefore active BSR, prevail in anoxic to suboxic conditions (Sasset al., 2002, 2006),and bicarbonate (HCO3-) produced through BSR plays an important role in increasing alkalinity in the microenvironments (Braissantet al., 2007; Duprazet al., 2009;Gallagheret al., 2012), the redox state of seawater will inevitably exert an impact on the development of fibrous aragonite layers. As an indispensable material source for the precipitation of fibrous aragonite, bicarbonate largely derived from BSR by the consumption of organic matter and sulfate, a relatively high microbial biomass in anoxic to suboxic environments (Grotzinger and Knoll, 1999),and sufficient sulfate supply in subtidal settings were likely the prerequisites. Therefore, it seems that an anoxic to suboxic background, shallow chemocline (Brockset al,2005; Tanget al., 2011), abundant microbial communities,and sufficient sulfate in the shallow Mesoproterozoic epeiric sea on the NCP provided favorable conditions for the precipitation of fibrous aragonite layers and the development of thrombolites.
7 Conclusions
Abundant thrombolites were recognized in the Mesoproterozoic Wumishan Formation (ca 1.50-1.45 Ga)of the North China Platform. Our study shows that they were mainly distributed in subtidal carbonate facies. From lower subtidal to lower intertidal facies, in response to decreasing accommodation and increasing hydrodynamics,thrombolites change morphologically from dominantly domal to tabular forms.
The nuclei of mesoclots in thrombolites may have resulted fromin situreplacement mineralization of microbial communities that likely induced by anoxic decomposition of organic matter via BSR. Thus, suboxic to anoxic conditions seem to be the important requirement for the formation of mesoclots, and less agitated environments in the subtidal zone are more conducive for the preservation of thrombolites. The fibrous outer rims surrounding nuclei
in mesoclots probably derived from carbonate precipitation facilitated by interactions between microbes and their microenvironments. Well-developed sub-millimetric aragonite fans and multi-layered micro-textures in the outer rims may provide evidence of rapid and multi-phase precipitation in an aragonite supersaturated micro-environment, which possibly resulted from geochemical changes caused by BSR in the ambient waters. As rapid carbonate precipitation and formation of fibrous aragonite rims surrounding mesoclot nuclei are critical for the preservation of mesoclots in the Wumishan thrombolites, it appears that suboxic to anoxic conditions, moderate sulfate supply, and therefore active BSR in subtidal zone are likely more favorable for the formation of thrombolites. Additionally,low detrital influx is also a basic requirement for aragonite development, and therefore for the rapid formation of mesoclots.
Acknowledgements
The study was supported by the Ministry of Science and Technology (No. 2011CB808806), and the National Natural Science Foundation (No. 41272039 and No. 40921062)of China. We are grateful to Prof. Feng Zengzhao, Franz T. Fürsich, Spencer G. Lucas and Wang Yongbiao for their constructive suggestions and enlightening comments,which improved the paper greatly. Thanks are given to Zhao Guisheng, Wu Jinjian and Li Danqiu for their assistance during field work.
 Journal of Palaeogeography2013年3期
Journal of Palaeogeography2013年3期
- Journal of Palaeogeography的其它文章
- Palaeogeographical zonation of gypsum facies:Middle Miocene Badenian of Central Paratethys(Carpathian Foredeep in Europe)
- Lithofacies and sedimentary characteristics of the Silurian Longmaxi Shale in the southeastern Sichuan Basin, China
- Lithofacies palaeogeography and biostratigraphy of the lowermost horizons of the Middle Triassic Hallstatt Limestones (Argolis Peninsula, Greece)
- Artemisia pollen-indicated steppe distribution in southern China during the Last Glacial Maximum
- Palinspastic reconstruction and geological evolution of Jurassic basins in Mongolia and neighboring China
- 1st International Palaeogeography Conference September 14–17, 2013 Beijing, China