
Artemisia pollen-indicated steppe distribution in southern China during the Last Glacial Maximum
2013-08-22 07:44:42LiuHongyanLiuKanWeiFangling
Journal of Palaeogeography 2013年3期
Liu Hongyan, Liu Kan, Wei Fangling
Department of Ecology and Ministry of Education Key Laboratory of Earth Surface Processes, College of Urban and Environmental Sciences, Peking University, Beijing 100871, China
Abstract The Last Glacial Maximum (LGM) was the coldest period during the previous 20,000 years. There have been different points of views on steppe distribution during the LGM period in southern China, partly due to the different interpretations of Artemisia occurrences.To make a reliable interpretation of the pollen fossil Artemisia, the modern distribution of Artemisia species and the relationship of pollen with climate and vegetation over a large spatial scale in China was thoroughly analyzed. Information about Artemisia species and pollen distributions used in this paper were collected from published works completed by other researchers as well as ourselves. The southern limit of steppe vegetation during the LGM period was interpreted from the published contour map of Artemisia pollen percentages during the LGM.Artemisia species in China are mostly distributed either in the horizontally distributed steppe regions or in the vertically distributed desert-steppe in the desert region, which indicates a cold and dry climate. The steppe is a distribution center of Artemisia pollen. Fractions of Artemisia in surface pollen assemblages are lower in both the desert and the temperate forest. Neither high Artemisia species cover nor high percentages of Artemisia pollen were found in the coast areas of China. Twenty-five percent of Artemisia pollen in sediments might indicate a local occurrence of steppe vegetation. Percentages of Artemisia pollen in the subtropical and tropical forest are less than 10%. A close relationship between Artemisia pollen and temperate steppe in China is demonstrated. The southern edge of the steppe vegetation during the LGM might be along the middle reach of the Yangtze River. Our results support the hypothesis that the isolated high fraction of Artemisia pollen along the northern edge of the South China Sea was transported from a large source area.
Key words pollen, steppe, Artemisia, Last Glacial Maximum, LGM, South China Sea
1 lntroduction*
The vegetation in low latitudes during the Last Glacial Maximum (18 ka BP) has perplexed Quaternary ecologists for a long time (Delcourt and Delcourt, 1991;Prentice and Jolly, 2000). Although it is quite commonly regarded that tropical vegetation had thoroughly disappeared during the LGM on the southern edge of mainland China, there are still different views regarding its vegetation patterns, particularly concerning the southward expansion of steppe vegetation. By simulation, Winkler and Wang (1993) pointed out that subtropical vegetation was sparse in coastal areas in southernmost China. However, some palynological workers argue that the current tropical vegetation region was partially occupied by the temperate steppe, or characterized by the occurrence of grassland dominated by Poaceae,Artemisiaand Chenopodiaceae, during the Last Glacial Maximum (LGM) (Liewet al., 1998; Sun and Li, 1999; Sunet al., 2000; Sun and Luo, 2004). Different interpretations of LGM vegetation patterns in southern China might be caused by local scale pollen data on one hand, and different understandings on the indication of pollen spectra, such as the occurrence of high fraction ofArtemisiapollen, on the other hand.
There are over 300Artemisiaspecies all over the world,mostly distributed in temperate Asia, Europe and North America (Editorial Committee of “Flora of China”, CAS,1991).Artemisiapollen is easily identified and quite frequently used during the reconstruction of palaeoclimate and palaeovegetation. It has been quite commonly linked to climatic deterioration during the glacial phases in Europe(Suc and Zagwijn, 1983), Asia (Yiet al., 2003a; 2003b),and North America (Thompsonet al., 1993). However, the different adaptation ofArtemisiaspecies to climatic factors introduces uncertainty in some localized palaeoclimatic and palaeovegetational reconstructions. The peak ofArtemisiapollen in the Mediterranean region, for instance,was demonstrated to be inconsistent with18O-indicated cold climate (Cheddadi and Rosignol-Strick, 1995). Some investigators also found that high percentages ofArtemisiapollen also occurred with high percentages of arboreal pollen which prefers a humid climate (Horowitz, 1989; Cheddadi and Rosignol-Strick, 1995).
To make a reliable interpretation of pollen fossilArtemisia, relationships between surface pollen spectra and climatic/vegetational patterns are essential. Based on the habitat of differentArtemisiaspecies in the eastern Mediterranean region, Subally and Quézel (2002) concluded thatArtemisiapollen is much more reliable in indicating humidity variations rather than temperature changes during glacial/interglacial cycles. Sunet al. (1996) argued that there are two types ofArtemisiaspecies in China, one prefers a cool-humid climate, while the other favors a warm and dry climate, derived from the pollen-climate response surface based on limited samples from northern China. The ratio ofArtemisiato Chenopodiaceae pollen is commonly considered to indicate humidity changes since it was first suggested in the Middle East by El-Moslimany (1990).However, some research also suggests thatArtemisiapollen can indicate humidity changes; on the other hand the high percentages of Chenopodiaceae pollen might be associated with human disturbances of vegetation (Liuet al.,2006).
The correlation ofArtemisiapollen to vegetation and climate factors is still unclear due to the lack of comprehensive analysis of surface pollen on a large spatial scale.The climatic zones of China include tropical to boreal climatic zones with various vegetations ranging from humid forest to semi-arid steppe and arid desert. In this paper, the distribution ofArtemisiaspecies and modern pollen with climate and vegetation zones was comprehensively analyzed based on published works in China, as well as the systematic work of the authors in a forest-steppe-desert transect in central Inner Mongolia and its surroundings.Distribution of steppe vegetation during the LGM period was then interpreted from the distribution of accessible pollen fossils (Members of China Quaternary Pollen Data Base, 2000).
2 Methods
Distribution ofArtemisiaspecies in China was collected from “Flora of China” (Chinese edition, Vol. 76-2); the 1:10,000,000 vegetation map of China was used to linkArtemisiaspecies to vegetation types. Information on the distribution ofArtemisiaspecies in relation to different types of vegetation was obtained from previous vegetation surveys in China described in the “Vegetation of China”(Editorial Committee of “Vegetation of China”, 1980) and some provincial vegetation publications, as well as field investigations by the authors in the temperate zone. The distribution ofArtemisiaspecies in each vegetation type was calculated as the mean of at least 5 plots. The mean annual temperature (MAT) and mean annual precipitation(MAP) for each vegetational type were obtained from digitized climate maps of China.
The authors collected 354 surface pollen samples from a deciduous broadleaved forest-steppe-desert transect in China (Liuet al., 1999, 2006). The publications on the distribution of surface pollen across horizontally and vertically distributed vegetation belts were also selected to show the distribution ofArtemisiapollen along with climatic factors and vegetational types.
3 Distribution of Artemisia species
3.1 Distribution of Artemisia species with vegeta‑tional types
There are 186 species and 44 variations of the genusArtemisiain China (Editorial Committee of “Flora of China”,CAS, 1991). No clear gradient was detected in terms of the density ofArtemisiaspecies in China.
Those vegetational types withArtemisiacover greater than 5% were selected from the vegetation map of China;they were mostly distributed in the northern part of China and in the Qinghai-Tibet Plateau of the southwestern China. The vegetational types included grass-dominated steppe,Artemisia-dominated desert and some temperate scrub. The only forest type wasPinus sylvestrisvar.mongolica-forest with a very limited distribution in the eastern part of the steppe region.
3.2 Distribution of Artemisia species with climatic factors
The mean cover ofArtemisiaspecies in all the plots of each vegetational type was plotted against mean annual precipitation (MAP) and mean annual temperature (MAT)(Fig. 1). AlthoughArtemisiaspecies adapt to a wide range of MAP and MAT, the peak appears at the MAP of 200-700 mm and the MAT of 2-8 ℃ approximately. We thus infer thatArtemisiaspecies indicate a cold and dry climate, which differs from the local investigation by Sunet al. (1996).
4 Artemisia in surface pollen spectra
4.1 Distribution of Artemisia pollen in steppe and desert zones
The steppe vegetation in Inner Mongolia of China and in southern Mongolia changes from meadow steppe in the southeastern region, through typical steppe and desert steppe, to desert in the northwestern region. It is characterized by both high abundance and the cover ofArtemisiaspecies. Based on the data from the deciduous broadleaved forest-steppe-desert transect in northern China, it is estimated that the mean fraction ofArtemisiapollen in the typical steppe is ~60%, in the meadow steppe it is 62%and in the desert steppe it is only 24.7%. At the neighboring forest fringe, it is as low as 20.8% (Fig. 2a, 2b; Liuet al., 1999, 2006). The mean fraction ofArtemisiain the desert region is ~25.5%, as calculated from the samples published by Liet al. (2005a).
Data from the neighboring Mongolia desert and steppe region were also used for comparison. As calculated from the supplementary data of surface pollen distribution in Mongolia by Guninet al. (1999), the mean percentage ofArtemisiapollen in the steppe region of Mongolia accounts for ~24%, higher than the 18% of the desert region.LowerArtemisiapercentages in the Mongolian steppe is probably related to lessArtemisiacover due to slighter grazing by livestock.
Community level of surface pollen assemblages has been comprehensively investigated in the steppe and the desert regions in northern China (Liet al., 2005a, 2005b).In less disturbed natural steppe dominated by grasses,such asStipa grandisandLeymus chinensis, the mean value ofArtemisiapollen accounts for 8%-13%. In the highly disturbed steppe, such asAgropyron-Cleistogenessteppe,Stipa kryloviisteppe, percentages ofArtemisiapollen approach 40%-55% due to the high percentages ofArtemisiacover of up to 40%-80%. The sandy scrub dominated byArtemisia ordosicahas a percentage ofArtemisiapollen of up to 55%. In theArtemisia-dominated loamy desert, the fraction ofArtemisiapollen is higher than that of Chenopodiaceae pollen with a percentage above 50% (Liet al., 2005b). On the Qinghai-Tibet Plateau,Artemisiais a common component of non-forest vegetation types (Yuet al., 2001).

Fig. 1 Percentage cover of Artemisia with respect to mean annual temperature (MAT) and mean annual precipitation (MAP).
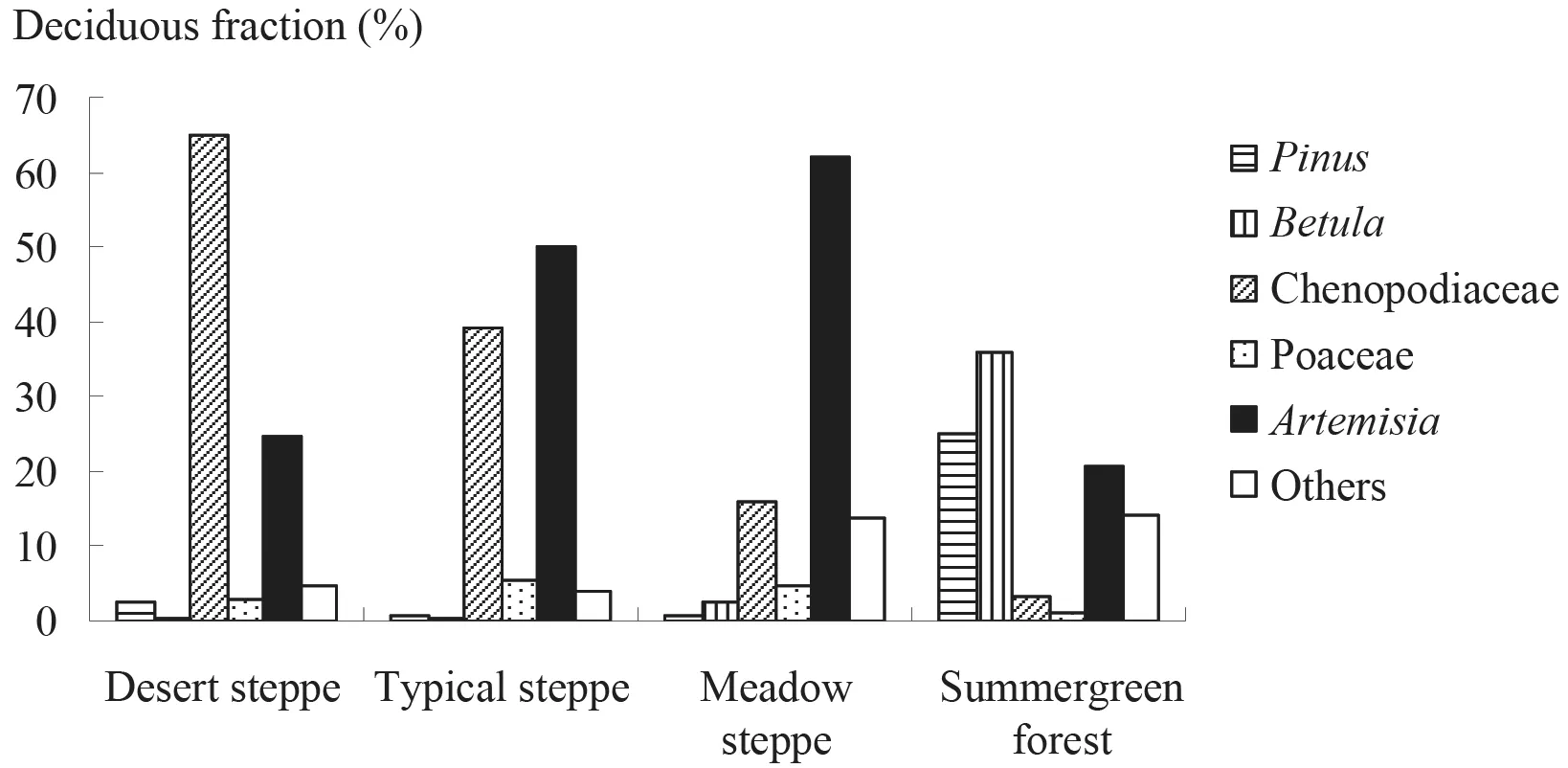
Fig. 2b Surface pollen spectra of the deciduous forest-steppe-desert transect.
The vegetation in mountains in the arid region changes from zonal desert vegetation at the foot of the mountains,through steppe desert and steppe, to coniferous forest and alpine meadow on the top. On the southern slope of the Tianshan Mountains (Fig. 2a), the forest belt is absent.The widely distributed steppe belt extends to 2800 m a.s.l.(a.s.1. means above sea level). The fraction ofArtemisiapollen is the highest in theArtemisia-dominated desertsteppe with ~55%. TheArtemisiapollen is deduced to come from the dominant speciesA. macrocephalain this belt (Yan, 1991; Xuet al., 1996). The vegetation on the northern slope is characterized by a forest belt above the steppe belt. The fraction ofArtemisiapollen in the desertsteppe zone with an elevation range of 700-1100 m a.s.l.reaches ~50% due to the widely distributedSerphidium(subgenus ofArtemisia) species, which might account for the high fraction ofArtemisiapollen (Yan, 1991; Ji,2005). The vegetation distribution on the northern slope of the Qilian Mountains (Fig. 3a) is quite similar to that on the northern slope of the Tianshan Mountains. Surface pollen investigations along elevations in its eastern and central part indicate that the highest percentage of theArtemisiapollen in the steppe-desert ranges from 1500 m to 1800 m a.s.l. at about 12%. In summary,unlike in the horizontally distributed vegetation, the highest percentage ofArtemisiaappears in the steppe-desert belt rather than in the steppe belt.
4.2 Distribution of Artemisia pollen in the tempera‑te forest zone
High percentages ofArtemisiapollen commonly appear in secondary scrubs in low mountainous areas in the middle of the temperate forest zone (Yu and Liu, 1997; Yanget al., 2004). High percentages ofArtemisia sacrorumin it may account for this phenomenon (Editorial Committee of“Vegetation of Hebei”, 1996).
Changbai Mountain (Fig. 2a) in the northern temperate forest region has a complete vertical vegetation spectrum from broadleaved forest at its foot, through mixed forest and coniferous forest, toBetula armandiidwarf forest and alpine tundra on its top. A decrease inArtemisiapollen from ~16% on the foot belt to 8% on the top belt was recalculated by Sunet al. (2003a). The vertical distribution of vegetation in the Xiaowutai Mountain(Fig. 2a) is very representative in the middle temperate forest region. Arboreal pollen dominates in all vegetation belts. The highest fraction ofArtemisiapollen occurs in the alpine meadow at 25%, possibly caused by high cover ofArtemisia tanacetifolia(Yu and Liu, 1997).Located in the southern temperate forest zone, the vegetation of Zhongtiao Mountain (Fig. 2a) is characterized by deciduous forest and mixed deciduous-coniferous forest. The fraction ofArtemisiapollen is lower than that on the Xiaowutai Mountain, with a maximum of 15% (Yao,1989).
A mean value ofArtemisiapollen in the temperate forest region is roughly estimated to be 10%-12% from the three mountains mentioned above, which preserve typical vertical vegetation spectra in the temperate forest zone.
4.3 Distribution of Artemisia pollen in the sub‑tropical and the tropical forest zones
Located at the northern edge of the subtropical forest region, the southern slope of the Qinling Mountains(Fig. 2a) is characterized by different types of forests up to 3500 m a.s.l.Artemisiapollen was found in almost all of the vegetation belts with a maximum percentage of 12.5%, which might be due toArtermisiaspecies in the herbaceous layer. Percentages ofArtemisiapollen of all the vertical vegetation belts are averaged to 8% (Zhaoet al., 1999). Occupied mainly by mixed evergreen-summergreen forest as well as mixed deciduous-coniferous forest, ZijinMountain (Fig. 2a) appears to have hosted very fewArtemisia, as even the maximum of its pollen percentage is no higher than 1% there (Yu and Han, 1995).Shennongjia Mountain (Fig. 2a) is the highest in the middle of the subtropical zone. Its vegetation changes from evergreen-deciduous forest to mixed deciduous-coniferous forests along with changes in elevation.Artemisiapollen is very limited in the deciduous summergreen forest and mixed deciduous-coniferous forest belts (Liuet al., 2001). Vegetation andArtemisiapollen distribution on Lushan Mountain (Fig. 2a) is quite similar to that on Shennongjia Mountain, with a maximum of up to 3.5%(Li, 1998). On Miao’er Mountain, only 2% ofArtemisiapollen was found under the evergreen broadleaved forest (Yao and Li, 1993). On Wuyi Mountain and Yulong Mountain (Fig. 2a), noArtemisiapollen was recorded across the vegetation belts at all (Li, 1998; Tonget al.,2003).
In tropical regions, such as in Hainan Island, noArtemisiapollen was found in all types of plant communities(Yu and Han, 1998).
4.4 Summary of Artemisia pollen as the indicators of climate and vegetation
The distribution ofArtemisiapollen can be summarized from the above review as: (1) High percentages ofArtemisiapollen are commonly associated with highArtemisiacover in plant communities. It is inferred thatArtemisiapollen mainly originated fromArtemisiaspecies in the temperate steppe, both horizontally and vertically distributed, and understoryArtemisiaspecies in the temperate forest region; (2) The massive dispersal ofArtemisiapollen is quite limited, particularly in forests. With the distribution center in the steppe region,percentages ofArtemisiapollen decline in both forests and deserts. Percentages ofArtemisiapollenin the steppe zone are higher than 25%,while in the forest-steppe transition, which is about 100 km wide, it is about 20%. In the temperate forest region,the fraction ofArtemisiapollen drops remarkably to about 10%.
We can roughly infer that over 25% ofArtemisiapollen indicates a local distribution of steppe. The fraction ofArtemisiapollen in the subtropical and tropical region is commonly less than 10%. The vertically distributed vegetation types in it do not alter the lowArtemisiapollen percentages.
5 lnterpretation of the southern limit of steppe during the LGM
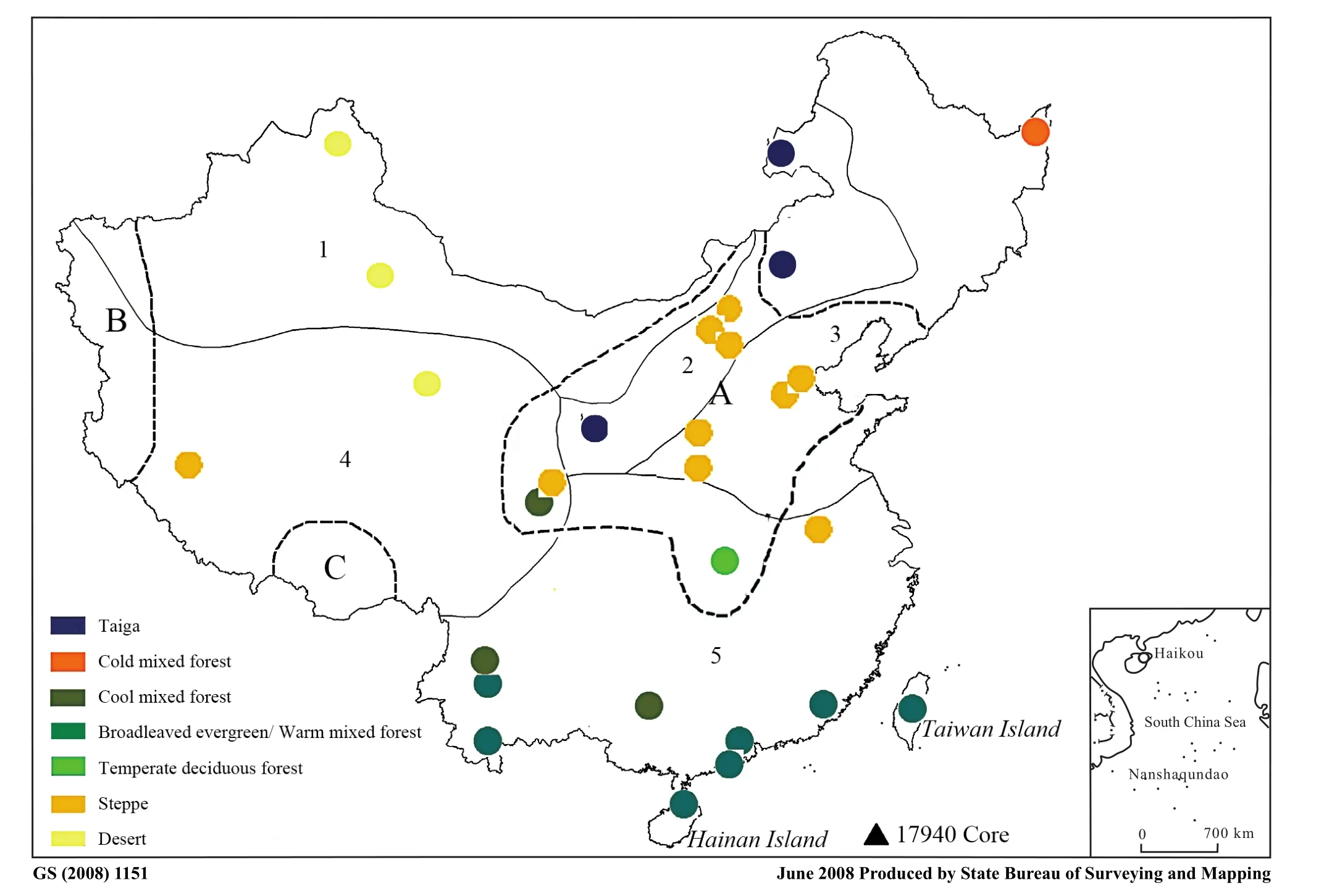
Fig. 3 Distribution map of steppe during the LGM by Biomisation method (from Yu et al., 2000) and indicated by Artemisia pollen distribution patterns. A, B and C denoted by dashed lines show the distribution of 25% Artemisia pollen during the LGM period with data from Members of China Quaternary Pollen Data Base (2000). 1-5 show the vegetation regions in Fig. 2a. The black triangle indicates the location of the 17940 core (Sun and Li, 1999; Sun et al., 2000).
The current steppe region and the southeastern and western edge of the Qinghai-Tibet Plateau was recognized as the distribution center ofArtemisiapollen from a contour map ofArtemisiapollen percentages during the LGM in China (Members of China Quaternary Pollen Database,2000), implying a similar vegetation gradient as at present(Fig. 3). By using the 25% value ofArtemisiapollen as a criterion to determine the boundary between temperate steppe and forest-steppe transition, the southernmost dis-tribution of the steppe was inferred to be along the middle reach of the Yangtze River (30˚N; Fig. 3), which is occupied by typical evergreen broadleaved forests. Both the modern summergreen broadleaved forest and mixed evergreen-summergreen broadleaved forest are interpreted to be occupied by steppe vegetation during the LGM. These results are approximately similar to the results of the Biomisation method (Yuet al., 2000). On the coastal area in the eastern part, however, the boundary extends much farther toward the north, implying the steppe had not reached the current southern temperate and subtropical/tropical coast even during the coldest LGM period, probably due to the humid climate along the coast during the LGM period.Our interpretation coincides well with the modern patterns ofArtemisiaspecies and steppe vegetation distributions.
6 Discussion
Although most of the pollen evidence supports subtropical forest or open forest vegetation in the southern edge of mainland China (e.g., Zheng, 1991; Winkler and Wang, 1993; Liu and Wang, 2004), some sites with high percentages of non-arboreal pollen, particularlyArtemisiapollen, during the LGM period, lead to a debate regarding regional vegetation patterns. Sun and Li (1999) deduced a steppe vegetation on the shelf at the northern edge of the current South China Sea based on greater than 40%Artemisiapollen in the 17940 core sampled there (Fig.3). They used the term “Artemisia-dominated grassland”instead of temperate steppe in their later works (Sun and Luo, 2001, 2004). Similar conclusions have also been drawn in Taiwan Island. Liewet al. (1998) found that assemblages of Gramineae,Artemisiaand Umbelliferae during the LGM are different from most of the rest of the last glacial deltaic sediments and deduced that a large part of the lowland area on the exposed shelf was grassland or barren during the LGM in central Taiwan. These works imply that the current tropical and subtropical vegetation in southern China were characterized by forests or woodlands in the mountains and grasslands in the lowlands.Liu and Wang (2004) suspected the woodland as zonal vegetation and grassland as azonal in specific landforms.Savanna vegetation characterized by high percentages of Poaceae pollen and arboreal pollen was also suggested based on pollen fossil spectra in the Leizhou Peninsula,north of Hainan Island (Zheng and Lei, 1999).
Although high percentages of Poaceae pollen were commonly recorded (Liewet al., 1998; Zheng and Lei,1999), the percentages ofArtemisiapollen exceeding 25%were only found along the northern edge of the eastern side of the South China Sea (the 17940 core; Sun and Li, 1999; Sunet al., 2000). In central Taiwan Island and Leizhou Peninsula, percentages ofArtemisiapollen is less than 10% (Liewet al., 1998; Zheng and Lei, 1999).From the distribution of modernArtemisiapollen in different vegetation regions, we further confirm that current tropical China was not occupied or even partly occupied by temperate steppes during the LGM. It is possible that savanna-like vegetation, which has no modern analog in China, did occur in southern China, as indicated by high percentages of Poaceae pollen, however it has no relation to the temperate steppe.
Based on the modern distribution ofArtemisiaspecies andArtemisiapollen, it is hard to interpret the very high percentage ofArtemisiain the current South China Sea.Liu and Wang (2004) suggested anArtemisia-dominated coastal meadow during the LGM on exposed shelves.However, both highArtemisiaspecies cover and high percentages ofArtemisiapollen were not found along the coast areas in China. We suspect that the highArtemisiapollen content might not have originated from the nearby exposed shelves. Surface pollen investigation in the South China Sea led to the conclusion that “pine pollen and fern spores, adapted to wind transportation and water flotation,come from the northeast through the straits borne on the winter monsoon and sea current. Their source areas should cover large regions, probably including south and southeast China” (Sunet al., 1999, 2003b); similar conditions might also have applied toArtemisiapollen. Another research based on pollen in air and water tends to support transportation ofArtemisiapollen to the South China Sea by wind rather than by water flow (Dai and Weng, 2011).Our analysis of the modern distribution ofArtemisiapollen and its relationship to vegetation supports the hypothesis that the isolated high fraction ofArtemisiapollen along the northern edge of the South China Sea was transported from a large source area.
In reality, there are uncertainties when relying on one pollen taxon to interpret palaeovegetation patterns. Due to the close link between high percentages ofArtemisiapollen and steppe vegetation, it is reliable for determining the ancient distribution of steppe vegetation on a large scale by using the distributionofArtemisiapollen.
7 Conclusions
Based on the distribution ofArtemisiaspecies and pollen in modern vegetation, a close relationship betweenAr-temisiapollen and temperate steppe in China is further confirmed.Artemisiaspecies in China are mostly distributed in the arid desert and the semi-arid steppe regions,indicating a cold and dry climate. The steppe is a distribution center ofArtemisiapollen. Fractions ofArtemisiain surface pollen spectra decline both in the desert and the temperate forest.
Twenty-five percent ofArtemisiapollen in the sediment might indicate a local occurrence of the steppe vegetation. The southern edge of the steppe vegetation during the LGM was along the middle reach of the Yangtze River.Our results suggest that the isolated high fraction ofArtemisiapollen along the northern edge of the South China Sea was transported from a large source area.
Acknowledgements
This paper was supported by National Natural Science Foundation of China Granted Projects (NSFC No.41071124).
 Journal of Palaeogeography2013年3期
Journal of Palaeogeography2013年3期
- Journal of Palaeogeography的其它文章
- Palinspastic reconstruction and geological evolution of Jurassic basins in Mongolia and neighboring China
- Environment controls on Mesoproterozoic thrombolite morphogenesis: A case study from the North China Platform
- Lithofacies palaeogeography and biostratigraphy of the lowermost horizons of the Middle Triassic Hallstatt Limestones (Argolis Peninsula, Greece)
- Lithofacies and sedimentary characteristics of the Silurian Longmaxi Shale in the southeastern Sichuan Basin, China
- Palaeogeographical zonation of gypsum facies:Middle Miocene Badenian of Central Paratethys(Carpathian Foredeep in Europe)
- General regulations about submitting manuscripts to Journal of Palaeogeography