
不同轮作制度下施肥对冬小麦田间杂草群落 及小麦生长的影响
2013-07-13 07:44程传鹏万开元陶勇汤雷雷潘俊峰陈防
生态环境学报 2013年3期
程传鹏,万开元,陶勇,汤雷雷,潘俊峰,3,陈防*
1. 中国科学院武汉植物园 中国科学院水生植物与流域生态重点实验室, 湖北 武汉 430074;2. 中国科学院大学,北京 100049;
3. 安徽农业大学资源与环境学院,安徽 合肥 230036;4. 国际植物营养研究所武汉办事处,湖北 武汉 430074
农田杂草作为农田生态系统的重要组成部分,可以与作物竞争多种资源(光照、土壤养分、水分等),是导致作物减产的重要因素之一[1-3]。另外,有研究报道也指出,杂草在促进田间养分循环,维持土壤生物多样性,减少土壤流失、酸化等方面具有重要作用[4-5]。因此,我们在进行田间杂草控制的同时,应考虑如何通过保持一定的田间杂草生物多样性来利用杂草的生态保护功能。
施肥作为农业生产中重要的养分管理措施可以影响土壤养分状况,改变作物与杂草之间的竞争关系,显著影响农田杂草群落组成,是潜在控制杂草的有效农田管理措施[6-7]。关于长期施肥方式对农田杂草群落和作物生长的影响研究已开展较多,结果表明长期不同施肥方式对田间杂草密度、物种组成、生物多样性及作物产量等都有显著影响[8-10]。但至今关于不同施肥方式对杂草群落的影响,国内外仍没有一个统一的结论。Kandasamy等[6]研究认为,平衡施肥处理(无论是无机肥配施还是施加有机肥)和未施肥处理时杂草干物质量较低,而N、P配施时杂草干物质质量较高。但Blackshaw等[9]研究却发现未施肥处理时杂草密度和干物质质量最低,而杂草密度和干物质质量较高的为撒施无机肥或堆肥的处理。另外,不同研究中对杂草发生及其生物多样性影响起主要作用的养分元素的报道差异也较大。Bittman等[11]在加拿大北部平原的研究表明,N对牧场杂草物种组成影响较大。Andreasen等[12]研究却发现土壤中P含量是影响大多数杂草发生的主要因素。赵锋等[13]的研究也表明,施磷是导致红壤稻田田间杂草总密度变化的原因之一。
以上结果之间有共同之处,同时也存在差异,其差异可能是由各研究者所在位点其他田间生产条件的不同引起的,尤其是作物轮作制度及土壤条件的差异,这可能会在一定程度上改变施肥对杂草群落的影响,而这其中轮作制度差异的影响可能更大[14-16]。轮作可以改变土壤营养水平[17-18]、土壤酶含量和活性[19]、土壤微生物和动物的种群与数量[3],这些条件的差异可以直接或间接地影响杂草群落和作物产量。不同轮作制度下作物属性的较大差异也是轮作影响杂草群落的重要原因,例如,特殊的生长习性和不同的资源竞争模式[3,20]。作物快速萌发和出苗、较快的叶面积和冠层发展、高叶面积指数和较长持续时间、较大植株高度等都有助于增加作物对杂草的竞争力[21],因为作物冠层覆盖可以通过减少光照、降低土壤表面温度、物理阻止杂草生长来抑制杂草。另外,还有不同轮作制度下化感作用[22]对杂草生长及杂草种子库影响[23-24]的差异的报道。
因此,我们认为不同轮作制度下这些条件的差异会在一定程度上改变不同轮作制度中不同施肥模式对杂草群落的影响。我们的研究选择3种轮作制度下的冬小麦田为研究对象,研究不同轮作制度下冬小麦田杂草群落与冬小麦生长变化规律,以期阐明不同轮作制度下施肥对田间杂草群落及作物生长的影响差异,为合理选择轮作制度及不同轮作制度下合理施肥模式提供理论依据。
1 材料与方法
1.1 研究地区概况和试验设计
本研究的长期田间肥效试验定位点分别位于中国安徽省蒙城县(33°13´38˝N, 116°36´58˝E和33°14´1˝N, 116°31´43˝E)和湖北省武汉市(30°29´23˝N,114°19´13˝E),3个长期定位点分布位置如图1所示。
1.1.1 安徽蒙城试验点
2个长期定位田间肥效试验点分别位于安徽省蒙城县马店村和七里许村,分别为冬小麦-大豆轮作和冬小麦-夏玉米轮作。试验点地处皖北平原中部,属暖温带半湿润季风气候;年平均气温14.8 ℃、降水量872.4 mm;土壤类型为砂浆黑土。
冬小麦-大豆轮作田间试验自1994年开始进行,前5年为冬小麦-夏玉米轮作,后改为冬小麦-大豆轮作至今。该处土壤N和P素缺乏,尤其是P素缺乏严重,K素相对较充足。实验处理为:(1)CK,对照;(2)PK;(3)NP;(4)NK;(5)NPK;(6)N1/2PK,施半量P肥和全量N、K肥。每年N、P和K肥仅在冬小麦播种前以底肥一次性施入,施肥量分别为N 187.5 kg·hm-2、P2O590 kg·hm-2和K2O 135 kg·hm-2,大豆种植期间不施肥。试验处理小区面积20 m2,3次重复,随机区组排列。采用除草剂对田间杂草进行控制,冬小麦季杂草三叶期每公顷喷施120 g苯磺隆(Tribenuron-methyl)1次,大豆播后苗前每公顷喷施1500 g乙草胺(Acetochlor)1次。

图1 长期田间肥效试验位点图 Fig. 1 Locations of the long-term field fertilization trials
冬小麦-夏玉米轮作试验自2000年夏玉米季开始。该处土壤N、P和K素都缺乏,尤其是P素缺乏严重。实验处理为:(1)NP,N180P90,N225P90;(2)NPK,N180P90K90,N225P90K120;(3)NP7/3K,N180P90K210,N225P90;(4)NP(7/3)K,N180P90,N225P90K210;(5)N2/3PK,N180P60K90,N225P60K120;(6)N3/2PK,N180P120K90,N225P120K120。每一处理中前面部分为冬小麦季施肥量,后面部分为夏玉米季施肥量,元素后面数字为N、P2O5和K2O施肥量(单位kg·hm-2)。试验处理小区面积40 m2,4次重复,随机区组排列。采用除草剂对田间杂草进行控制,冬小麦季杂草三叶期每公顷喷施120 g苯磺隆1次,玉米播后苗前每公顷喷施1500 g乙草胺1次。
1.1.2 湖北武汉试验点
该长期田间肥效试验点设在湖北省农科院南湖试验站,为冬小麦-中稻轮作。试验点处在北亚热带向中亚热带过渡型的地理气候带,年平均气温16.7 ℃,年降雨量1300 mm左右;土壤类型为黄棕壤发育的黄泥田;地形为垄岗平原。
该试验于1981年中稻季开始,水旱轮作。该处土壤有效N较充足,P素不足,K素也不丰富。试验共设9个处理,本研究只选取其中的5个处理进行观察:(1)CK,对照;(2)N;(3)NP;(4)NPK;(5)OM+NPK,施常量有机肥、N、P和K肥。无机肥料用量为每年全年施用N 150 kg·hm-2,P2O575 kg·hm-2,K2O 150 kg·hm-2。有机肥料为鲜猪粪,堆置田头一周后施用,施用量为11 250 kg·hm-2。无机肥料的施用量中,稻占60%,小麦占40%;有机肥料的施用量中,稻、小麦各占一半。肥料的施用方式:在水稻和小麦上P和K肥、有机肥料均采用移栽播种前一次全层基施,氮肥在水稻上基肥40%、分蘖肥40%、穗肥20%;小麦基肥50%、腊肥25%、拔节肥25%。试验处理小区面积40 m2,3次重复,随机区组排列。采用除草剂对田间杂草进行控制,冬小麦苗期每公顷喷施150 g苯磺隆1次,中稻苗期每公顷喷施1600 g苄嘧·丙草胺(Pretilachlor)1次。
1.2 田间调查
于2012年冬小麦成熟期进行田间杂草调查,每小区随机设置3个50 cm×50 cm样方,记录所有杂草物种数、每种数量,并将地上所有杂草植株取出,置烘箱内70 ℃烘干后称量,测定其地上生物量。同时,使用照度计(型号T-1H)分别准确测量地表和冬小麦冠层顶部的光照强度,计算光照透过率。在冬小麦收获时测产和进行室内考种,测定冬小麦地上生物量和产量。
1.3 数据处理
研究数据使用Excel 2003、Arcgis 9.3和SigmaPlot 10.0进行处理、绘图,并使用SPSS 17.0进行数据统计分析,测定各处理间的差异显著性。
杂草密度为单位面积杂草株数。相对密度以每处理中某给定杂草密度除以该处理所有杂草的总密度来计算。通过计算不同的生物多样性指标来评估杂草群落物种多样性。物种丰富度S为每处理平均杂草物种数。杂草群落的香农物种多样性指数H’(Shannon-Wiener Index),辛普森指数D(Simpson Index)和均匀度指数E(Evenness Index)按如下方程计算:
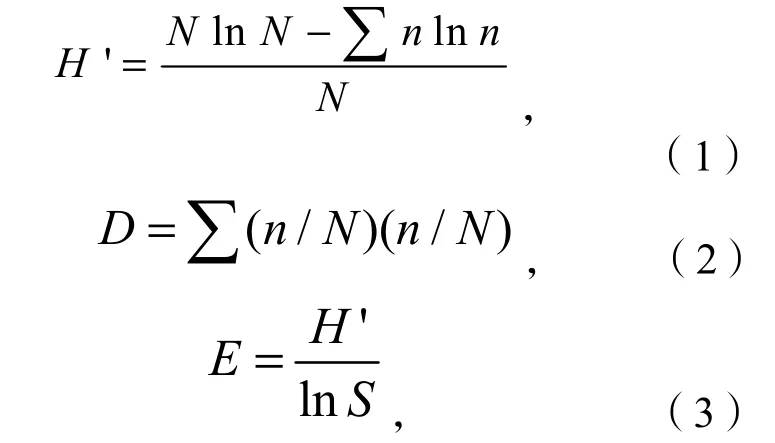
式中,N表示处理内所有杂草物种个体的总数目,n表示处理内每种杂草物种个体的数目。田间光照透过率(%)用来表示杂草可获得的光照强度的大小,以地表光照强度与冠层上方光照强度的比值来表示。
2 结果
2.1 杂草密度和地上生物量
3种轮作制度的冬小麦田中共观察到杂草16科33种,其中冬小麦-大豆轮作中有12科17种;冬小麦-中稻轮作中有9科15种(表1);冬小麦-夏玉米轮作中有6科10种。在冬小麦-大豆轮作中刺儿菜Cirsium segetum、鹅观草Roegneria kamoji、小飞蓬Conyza canadensis、野老鹳草Geranium carolinianum、大巢菜Vicia sativa、波斯婆婆纳Veronica persica 6种杂草发生较多,占田间杂草的90%。冬小麦-中稻轮作中发生较多的杂草物种有水蓼Polygonum hydropiper、菵草Beckmannia syzigachne、鳢肠Eclipta prostrata、斑种草Bothriospermum chinense、牛筋草Gramineae 5种,其比例也达到了90%(表1)。而在冬小麦-夏玉米轮作中猪殃殃Galium aparine占到了绝对优势,其密度比例高达95%。
在冬小麦-大豆轮作中,PK处理的杂草总密度最高(335.11株·m-2),除NK处理外,显著高于其他各个处理。施N和P肥的处理杂草总密度相对较低,其中以NP处理杂草总密度最低(29.33株·m-2)。杂草地上生物量趋势与杂草总密度基本一致,差别较大的是NP和NPK处理,NPK处理杂草地上生物量最小(9.57 g·m-2)。在冬小麦-中稻轮作中,OM+NPK处理的杂草总密度最低(88.44株·m-2);NP处理的总杂草密度最高(449.33株·m-2)。其他各处理的杂草总密度差异不显著;而且杂草地上生物量趋势与杂草总密度趋势基本相同,最小的为OM+NPK处理,只有1.95 g·m-2(表1)。在冬小麦—夏玉米轮作中杂草总密度都较低,最低的NPK处理为22.00株·m-2,最高的NP7/3K处理也只有48.00株·m-2;而且杂草地上生物量趋势与总密度趋势基本相同,最低的NPK处理为13.43 g·m-2,最高的NP7/3K处理为29.07 g·m-2(表2)。
以上结果表明,在不同轮作制度下,施肥模式对冬小麦田杂草密度和杂草地上生物量的影响是基本一致的。平衡施用N、P、K肥或者NPK肥配施有机肥均可以显著降低田间杂草密度和地上生物量,但是不同轮作制度下施肥模式对杂草密度和杂草地上生物量影响的程度有较大差异(表1、2)。杂草密度和地上生物量相对值对比表明,在平衡施肥条件下,冬小麦-中稻轮作中施肥对杂草密度和地上生物量的影响小于冬小麦-大豆轮作中施肥的影响,并且除NP处理外,冬小麦-中稻轮作与冬小麦-大豆轮作相比其他施肥处理对杂草密度和地上生物量的影响与平衡施肥处理相比差异程度也较小。冬小麦-夏玉米轮作中施肥对杂草密度和地上生物量的影响略小于冬小麦-大豆轮作冬小麦田中施肥的影响。综上所述,3种轮作制度改变冬小麦田中施肥对杂草密度和地上生物量影响程度的顺序为:冬小麦-中稻轮作>冬小麦-夏玉米轮作>冬小麦-大豆轮作。

表1 不同轮作制度下施肥对冬小麦田间杂草相对密度、总密度和地上生物量的影响 Table 1 Weed species relative density, total density and aboveground biomass in different treatments under different rotations
2.2 优势杂草种类和杂草群落生物多样性
长期不同施肥处理不仅会改变田间杂草总密度,也改变各种杂草在群落中所占的相对比例(表1)。在冬小麦-大豆轮作中,优势杂草种类较多的为PK和N1/2PK处理,都有4种。其中PK处理优势杂草为鹅观草、小飞蓬、大巢菜和波斯婆婆纳;N1/2PK处理为刺儿菜、小飞蓬、野老鹳草和荠菜Capsella bursa-pastoris。优势杂草种类较少的为CK和NK处理,都只有2种,而且都是刺儿菜和鹅观草。其余2个处理(NP和NPK处理)都有刺儿菜、小飞蓬和野老鹳草3种优势杂草。在冬小麦-中稻轮作中,OM+NPK处理优势杂草种类最多,高达4种,分别为水蓼、菵草、牛筋草和空心莲子草Alternanthera philoxeroides。其次为N处理和NPK处理,都有3种,分别为水蓼、菵草、斑种草和水蓼、菵草、鳢肠。优势杂草种类最少的为CK和NP处理,都只有2种,都是水蓼和菵草。由此可见,在2种轮作制度中,平衡施用N、P、K肥或者NPK肥配施有机肥处理的优势杂草种数较多,且种类更加多样,而在CK处理中优势杂草种类很少且种类单一,一般为总田间杂草群落优势物种。而且2种轮作制度改变施肥对优势杂草种类数影响的程度基本相同。冬小麦-夏玉米轮作中各施肥处理优势杂草种类只有猪殃殃1种,这可能是因为该轮作中施肥处理差异较少。
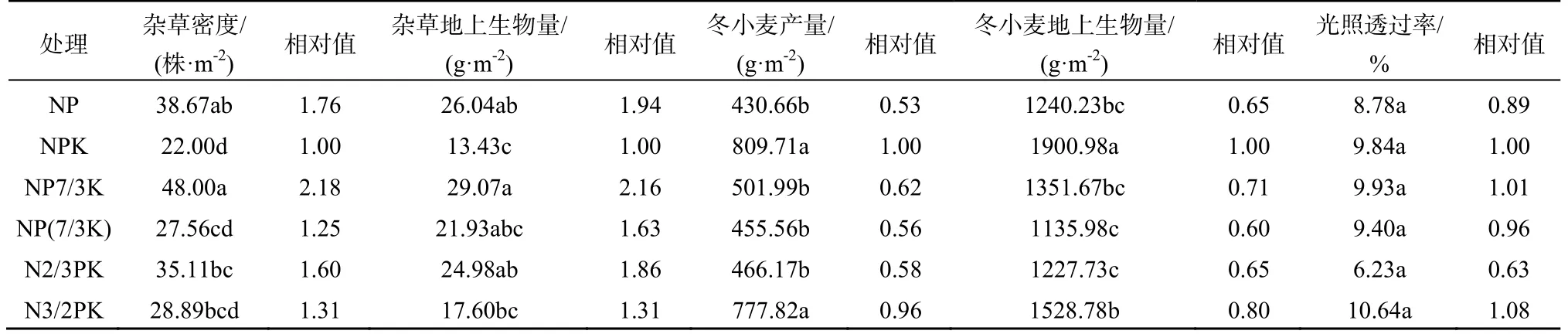
表2 冬小麦-夏玉米轮作制度下施肥对冬小麦田间杂草密度、地上生物量、冬小麦产量、地上生物量和光照透过率的影响 Table 2 Weed species density, aboveground biomass, winter wheat yield, aboveground biomass and light transmittance in different treatments under winter wheat – summer maize rotation
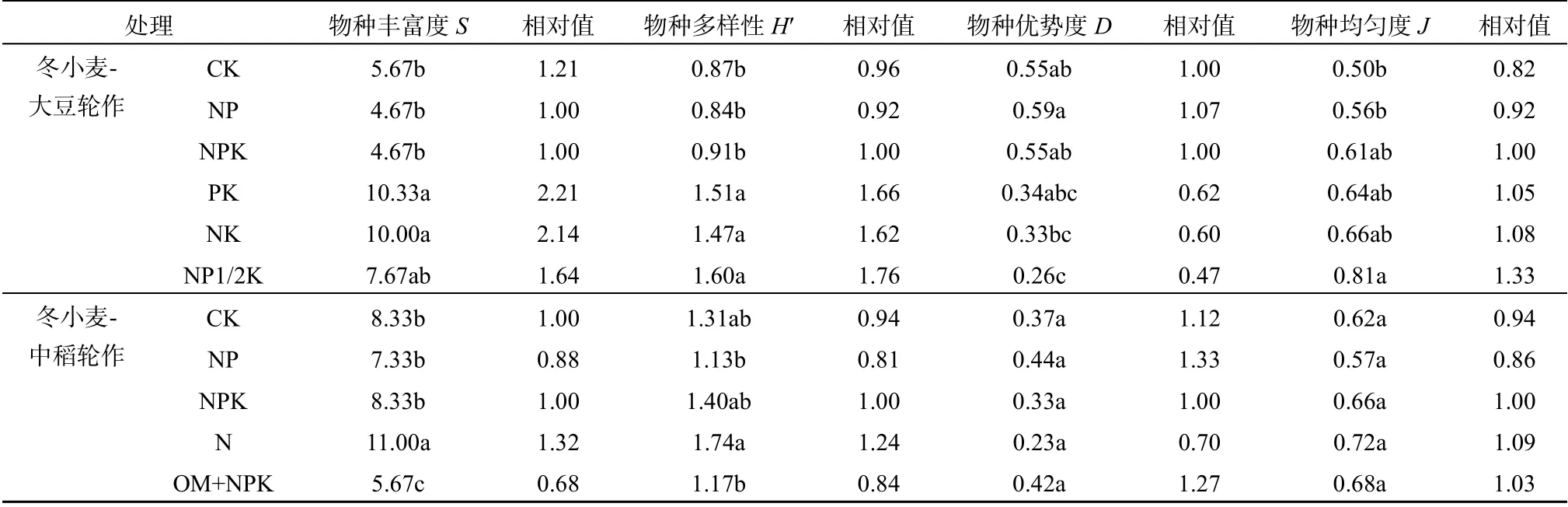
表3 冬小麦-大豆轮作和冬小麦-中稻轮作制度下冬小麦田间杂草群落物种丰富度S、 物种多样性指数H'、均匀度指数J和辛普森优势度指数D Table 3 Weed species richness (S), Shannon-Wiener Index (H'), Evenness Index (, J) and Simpson Index (D) of weed communities in different treatments under winter wheat – soybean rotation and winter wheat - mid-season rice rotation
长期施肥处理对田间杂草总密度和分布的影响,可能改变各处理田间杂草群落的生物多样性。在冬小麦-夏玉米轮作制度中,田间杂草以猪殃殃占绝对优势,而其他杂草种类在各处理中的分布较随意,未计算其田间杂草生物多样性指数。另外2种轮作制度下,各施肥处理田间杂草生物多样性指数明显不同(表3)。在冬小麦-大豆轮作中杂草群落物种丰富度以PK和NK处理的最高,达10种,除N1/2PK处理(7~8种),显著高于其他各处理(4~6种)。物种多样性指数和均匀度指数均以N1/2PK处理最高,相反,其优势度指数较低,而CK和NP处理具有较高的杂草群落物种优势度。在冬小麦-中稻轮作中杂草群落物种丰富度最高的为N处理,达11种,显著高于其他各处理(5~9种);其次为CK和NPK处理(8~9种),最低的OM+NPK处理只有5~6种。物种多样性指数较高的为N、NPK和CK处理,显著高于其他各处理。均匀度指数较高的为N、NPK和OM+NPK处理,但各处理间差异不显著。优势度指数趋势与均匀度指数趋势也相反,但各处理间差异性并不显著。由此可见,在2种轮作的冬小麦田中,尽管平衡施用N、P、K肥的处理杂草群落物种丰富度并不最高,但其物种多样性指数和均匀度指数均较高,并且优势度指数也较低。不同轮作制度下杂草生物多样性指数相对值对比表明,2种轮作制度改变冬小麦田中施肥对杂草生物多样性影响的程度差别不大。

表4 冬小麦-大豆轮作和冬小麦-中稻轮作制度下施肥对冬小麦产量及地上生物量的影响 Table 4 Winter wheat yield and aboveground biomass in different treatments under winter wheat – soybean rotation and winter wheat – mid-season rice rotation
2.3 冬小麦产量及生长
施肥作为增加作物产量的一项主要措施,在增加冬小麦产量和地上生物量上具有重要作用。由表2、4可知,在3种轮作制度下,施肥均可以对冬小麦产量和地上生物量造成影响。而且在3种轮作制度中施肥对冬小麦产量和地上生物量影响趋势基本相同,只在冬小麦-夏玉米轮作中施肥对冬小麦地上生物量的影响大于产量。在冬小麦-大豆轮作中,冬小麦产量和地上生物量均以N1/2PK和NPK处理较高,CK和PK处理较低,NP和NK处理居中;在冬小麦-中稻轮作中,冬小麦产量和地上生物量均以OM+NPK和NP处理较高,其次为NPK处理,而CK和N处理较低;在冬小麦-夏玉米轮作中,冬小麦产量和地上生物量均以NPK处理最高,但其他处理产量和地上生物量差异较小,其地上生物量差异大于产量。以上结果表明,3种轮作制度下,平衡施用N、P、K肥或者NPK配施有机肥均可以提高冬小麦产量和地上生物量。不同轮作制度下冬小麦产量和地上生物量相对值对比表明,供试条件下,3种轮作制度改变施肥对冬小麦产量和地上生物量影响程度的顺序为:冬小麦-中稻轮作>冬小麦-大豆轮作>冬小麦-夏玉米轮作。
2.4 光照透过率

图2 冬小麦-大豆轮作和冬小麦-中稻轮作制度下施肥 对冬小麦田间光照透过率的影响 Fig. 2 Light transmittance of winter wheat field in different treatments under winter wheat – soybean (WS) rotation and winter wheat –mid-season rice (WR) rotation
施肥对作物生长的影响会进一步改变作物冠层透过的光照。图2显示了冬小麦-大豆和冬小麦-中稻2种轮作制度下施肥对冬小麦田地表光照透过率的影响。结果表明,2种轮作制度下施肥对冬小麦田地表光照透过率影响基本一致。在冬小麦-大豆轮作中,CK处理光照透过率显著高于其他各处理;施肥处理对冬小麦田光照透过率影响较大的为NPK、NP和N1/2PK处理,显著大于其他2个处理。在冬小麦-中稻轮作中,CK和N处理光照透过率都显著高于其他各处理,施肥处理对冬小麦田光照透过率影响最大的为OM+NPK处理,其光照透过率显著低于其他处理;NPK和NP处理对作物各冠层光照透过率的影响基本相同。总的来说,在2种轮作制度下,施加N肥和P肥的处理都可以降低冬小麦田光照透过率,并且平衡施用N、P、K肥或者NPK配施有机肥对冬小麦田光照透过率影响较大。由图2还可知,施肥对冬小麦田间光照透过率的影响在冬小麦-中稻轮作中小于在冬小麦-大豆轮作中的影响。对比位于同一地点的冬小麦-夏玉米轮作和冬小麦-大豆轮作发现,冬小麦-夏玉米轮作中施肥对冬小麦田间光照透过率的影响略低。综上所述,供试条件下3种轮作制度中,施肥对冬小麦田间光照透过率影响程度的顺序为:冬小麦-中稻轮作>冬小麦-夏玉米轮作>冬小麦-大豆轮作。这与3种轮作制度中施肥对杂草密度和地上生物量影响程度的顺序相同。
地表获得光辐射的差异会显著影响杂草的种子萌发和幼苗生长,进而影响田间杂草发生量。图3给出了冬小麦-大豆轮作和冬小麦-中稻轮作制度下杂草密度与田间光照透过率之间的相关性。研究发现,2种轮作制度下田间杂草密度都随着田间光照透过率的增加而增加;而且冬小麦-大豆轮作中田间光照透过率与杂草密度的相关性(R=0.840 6)比冬小麦-中稻轮作(R=0.790 6)中要大,其直线斜率也较大,但两种轮作制度中的高相关性都显示了田间光照透过率在影响杂草发生中的作用。由于冬小麦-夏玉米轮作的冬小麦田中小麦生长较好,对田间光照透过率影响都较大,导致冬小麦田间光照透过率都较小,基本都在10%以下,其田间杂草密度与田间光照透过率相关性不明显,故未作分析。

图3 冬小麦-大豆轮作和冬小麦-中稻轮作制度下杂草密度 与冬小麦田间光照透过率之间的相关性 Fig. 3 Relationship between weed density and light transmittance of winter wheat fields under winter wheat – soybean (WS) rotation and winter wheat – mid-season rice (WR) rotation
3 讨论
施肥作为一项重要的农田养分管理措施,可以显著影响土壤养分状况,并在一定程度上改变作物与农田杂草之间的相互关系,形成具有不同丰饶水平特点和杂草生物多样性的杂草群落[6-7]。本研究中,施肥对3种轮作制度中冬小麦田杂草密度和地上生物量的影响趋势是基本一致的,平衡施用N、P、K肥或者NPK肥配施有机肥均可以显著降低田间杂草总密度和地上生物量;因为在平衡施用N、P、K肥或者NPK肥配施有机肥时,适宜的养分条件能改变作物与杂草以及杂草与杂草之间的竞争关系,有利于作物生长,抑制杂草生长。而在N、P和K肥单施或者两两配施时,施肥对冬小麦田间杂草密度和地上生物量的影响差异较大,这可能与3种轮作条件下土壤养分含量特征有关。此结果与Kandasamy等[6]和Nie等[25]的研究结果一致。但3种轮作制度改变冬小麦田中施肥对杂草密度和地上生物量影响的程度是不同的,这可能与3种轮作制度下不同作物轮作对土壤水肥状况改变的差异[24]、轮作作物对杂草化感作用的不同[22]以及前季轮作作物在形态特征、生长特点上的差异[14,26]有关。这些条件下的差异会影响冬小麦与杂草的竞争关系,改变两者对田间养分的吸收量和吸收效率,进而改变田间杂草密度和地上生物量。另外,轮作中其他作物对杂草种子库影响的差异对此也有一定的影响[23,27]。施肥对3种轮作制度下冬小麦田杂草群落优势杂草种类和生物多样性的影响略有差异,在冬小麦-大豆轮作和冬小麦-中稻轮作制度下平衡施用NPK肥的处理田间杂草群落优势杂草种数较多,且优势度指数较低,说明在这两种轮作制度下平衡施用N、P、K肥可以在控制杂草密度的同时保持一个生物多样性较高的杂草群落。但在冬小麦-夏玉米轮作制度下,优势杂草种类只有1种,这可能是因为在该轮作制度下冬小麦田施肥处理对冬小麦生长促进作用较强,使得冬小麦比其他田间杂草更具竞争力,而猪殃殃由于其特殊的生长特性(具有蔓生攀援性等构型特点)可以正常生长,导致了田间杂草群落的单一化。另外,研究也表明轮作制度改变冬小麦田中施肥对优势杂草种类数和杂草生物多样性影响的程度差别不大。这可能是因为轮作只是改变了施肥对田间杂草生长发育的影响程度,并没有达到引起田间杂草物种消亡的程度。
施肥在促进作物生长和增加作物产量上的作用早已被人们所认识[1,11,17,28]。本研究结果表明,在3种轮作制度下,平衡施用N、P、K肥或者NPK肥配施有机肥均可以提高冬小麦产量和地上生物量。因为在平衡施肥条件下,冬小麦获得营养元素较充足,也更加均衡,而适宜的养分条件有利于冬小麦生长发育和果实成熟。但3种轮作制度改变施肥对冬小麦产量和地上生物量影响的程度并不相同。这可能与3种轮作制度下不同作物轮作对土壤水、气、肥等状况改变的差异有关。例如在冬小麦-中稻轮作制度下,水旱轮作改变了田间的水湿环境,前季作物中稻需水较多,降低了后续作物冬小麦田中土壤氧气浓度、微生物种类和数量、土壤酶活性等,减少了冬小麦对土壤养分元素的吸收利用[23-24,27]。在农田生态系统中,杂草除与作物竞争养分、水分等资源外,对光照的竞争也很剧烈[25-26,29-30]。因为杂草可获得R/FR(Red to far-red photo ratio)比率的改变对杂草种子萌发、植株定植以及生长发育和结实都有显著影响,这可以在很大程度上改变田间杂草群落及其生物多样性[12,31]。研究表明,不同光辐射R/FR 比率影响单位面积杂草植株数目[29]。在本研究中,冬小麦-大豆轮作和冬小麦-中稻轮作制度下田间杂草密度都随着田间光照透过率的增加而增加,并且杂草密度与冬小麦田间光照透过率之间的高相关系数也表明了杂草可获得光照对田间杂草发生的影响,这与前人的研究结果是一致的。另外,在3种轮作制度下,不同施肥处理都可显著影响冬小麦田光照透过率,并且3种轮作制度改变施肥对冬小麦田间光照透过率影响程度的顺序与3种轮作制度改变冬小麦田中施肥对杂草密度和地上生物量影响程度的顺序是一致的,这也在另一方面证明了田间光照透过率对田间杂草的显著影响。由此可见,施肥对冬小麦田间光照透过率的改变可能是施肥影响冬小麦田间杂草群落的主要途径之一。
因此,在不同轮作制度的冬小麦田中,平衡施用N、P和K肥,不仅可以促进冬小麦生长,保证产量,还可以维持一定的田间杂草生物多样性,发挥其在维持生态平衡中的作用。掌握不同轮作制度下施肥对杂草群落及作物生长影响的差异还有利于科学合理地选择轮作制度,并在不同轮作制度中合理施肥。本研究对指导轮作制度的合理选择和科学合理施肥,从而兼顾农业生产和保护农田生态环境两方面的需要具有重要意义,对于后续研究的进行也有很好的借鉴意义。
4 结论
3种轮作制度下,施肥对冬小麦田间杂草群落及小麦生长的影响趋势是基本一致的。平衡施用N、P、K肥或者NPK肥配施有机肥均可以在促进小麦生长抑制田间杂草群落的同时,保证一定的田间杂草生物多样性。但不同轮作制度对施肥在冬小麦田中的作用有一定的影响,这与不同轮作制度下土壤条件和田间光照的差异有关。
[1] OKAFOR L I, DATTA S K. Competition between upland rice and purple nutsedge for nitrogen, moisture and light[J]. Weed Science, 1976, 24: 43-46.
[2] BUHLER D D, LIEBMAM M, OBRYCKI J J. Theoretical and practical challenges to an IPM approach to weed management[J]. Weed Science, 2000, 48: 274-280.
[3] VOLLMANN J, WAGENTRISTL H, HARTL W. The effects of simulated weed pressure on early maturity soybeans[J]. European Journal of Agronomy, 2010, 32(4): 243-248.
[4] ALBRECHT H. Suitability of arable weeds as indicator organisms to evaluate species conservation effects of management in agricultural ecosystems[J]. Agriculture, Ecosystems 暗淡 Environment, 2003, 98: 201-211.
[5] FRIED G, PETIT S, DESSAINT F, et al. Arable weed decline in Northern France: crop edges as refugia for weed conservation? [J].Biological Conservation, 2009, 142: 238-243.
[6] KANDASAMY O S, BAYAN H C, SANTHY P, et al. Long-term effects of fertilizers application and three crop rotations on changes the weed species in the 68th cropping (after 26 years) in the 68th cropping(after 26 years) [J]. Acta Agronomica Hungarica , 2000, 48: 149-154.
[7] YIN L C, CAI Z C, ZHONG W H. Changes in weed community diversity of maize crops due to long-term fertilization[J]. Crop Protection, 2006, 25: 910-914.
[8] SANTOS B M, DUSKY J A, STALL W M, et al. Influence of common lambsquarters (Chenopodium album) densities and phosphorus fertilization on lettuce[J]. Crop Protection, 2004, 23: 173-176.
[9] BLACKSHAW R E, MOLNAR L J, LAMEY F J. Fertilizer, manure and compost effects on weed growth and competition with winter wheat in western Canada[J]. Crop Protection, 2005, 24: 971-980.
[10] DAVIS A S. Nitrogen fertilizer and crop residue effects on seed mortality and germination of eight annual weed species[J]. Weed Science, 2007, 55: 123-128.
[11] BITTMAN S, MCCARTNEY D H, WADDINGTON J, et al. Long-term effects of fertilizer on yield and species composition of contrasting pasture swards in the Aspen Parkland of the Northern Great Plains[J]. Canadian Journey of Plant Science, 1997, 77: 607-614.
[12] ANDREASEN, C, SKOVGAARD, I M. Crop and soil factors of importance for the distribution of plant species on arable fields in Denmark[J]. Agriculture, Ecosystems and Environment, 2009, 133: 61-67.
[13] 赵锋,董文军,芮雯奕,等.不同施肥模式对南方红壤稻田冬春杂草群落特征的影响[J]. 杂草科学, 2009 (1): 7-12.
[14] MURPH Y K M, DAWSON J C, JONES S S. Relationship among phenotypic growth traits, yield and weed suppression in spring wheat landraces and modern cultivars[J]. Field Crops Research, 2008, 105: 107-115.
[15] SOSNOSKIE L M, HERMS C P, CARDINA J, et al. Seedbank and emerged weed communities following adoption of glyphosate-resistant crops in a long-term tillage and rotation study[J]. Weed Science, 2009, 57: 261-270.
[16] RYAN M R, CURRAN W S, GRANTHAM A M, et al. Effects of seeding rate and poultry litter on weed suppression from a rolled cereal rye cover crop[J]. Weed Science, 2011, 59: 438-444.
[17] BLACKSHAW R E, BRANDT R N, HENRY J H, et al. Differential response of weed species to added nitrogen[J]. Weed Science, 2003, 51: 532-539.
[18] LUPWAYIA N Z, HANSONB K G, HARKERC K N, et al. Soil microbial biomass, functional diversity and enzyme activity in glyphosate-resistant wheat–canola rotations under low-disturbance direct seeding and conventional tillage[J]. Soil Biology and Biochemistry, 2007, 39(7): 1418-1427.
[19] 徐培智, 解开治, 陈建生, 等.一季中晚稻的稻菜轮作模式对土壤酶活性及可培养微生物群落的影响[J].植物营养与肥料学报, 2008, 14(5): 923-928.
[20] LIEBMAN M, DYCK E. Crop rotation and intercropping strategies for weed management[J]. Ecological Applications, 1993, 3(1): 92-122.
[21] MASON H E, NAVABI A, FRICK B L, et al. The weed-competitive ability of Canada western red spring wheat cultivars grown under organic management[J]. Crop Science, 2007, 47: 1167-1176.
[22] 王瑞龙, 张墨溪, 宋圆圆, 等.豆科牧草对4 种农田常见杂草和水稻化感作用的研究[J]. 生态环境学报,2010, 19(10): 2307-2312.
[23] CARDINA J, HERMS C P, DOOHAN D J. Crop rotation and tillage system effects on weed seedbanks[J]. Weed Science, 2002, 50: 448-460.
[24] 魏守辉,强胜,马波,等.不同作物轮作制度对土壤杂草种子库特征的影响[J]. 生态学杂志, 2005, 24(4): 385-389.
[25] NIE J, YIN L C, LIAO Y L, et al. Weed community composition after 26 years of fertilization of late rice[J]. Weed Science, 2009, 57: 256-260.
[26] DIDON U M E. Variation between barley cultivars in early response to weed competition[J]. Journal of Agronomy and Crop Science, 2002, 188: 176-18.
[27] 强胜,胡金良.江苏省棉区棉田杂草群落发生分布规律的数量分析[J]. 生态学报, 1999, 19(6): 810-816.
[28] POGGIO S L, SATORRE E H, DE LA FUENTE E B. Structure of weed communities occurring in pea and wheat crops in the Rolling Pampa (Argentina) [J]. Agriculture, Ecosystems and Environment, 2004, 103: 225-235.
[29] BALLARE C L, CASAL J J. Light signals perceived by crop and weed plants[J]. Field Crops Research, 2000, 67: 149-160.
[30] 李儒海, 强胜, 邱多生, 等. 长期不同施肥方式对稻油轮作制水稻田杂草群落的影响[J]. 生态学报, 2008, 28(7): 3236-3243.
[31] HASHEM A, RADOSEVICH S R, DICK R. Competition effects on yield, tissue nitrogen, and germination of winter wheat (Triticum aestivum) and Italian Ryegrass (Lolium multiflorum) [J]. Weed Technology, 2000, 14: 718-725.
猜你喜欢
中国机械工程(2022年8期)2022-05-09
今日农业(2021年11期)2021-11-27
中国机械工程(2021年8期)2021-05-07
音乐教育与创作(2019年8期)2019-05-16
中国化肥信息(2018年12期)2018-03-01
新农业(2017年24期)2018-01-30
植物保护(2017年1期)2017-02-13
中学生(2015年4期)2015-08-31
新疆农垦科技(2014年5期)2014-02-28
新疆农垦科技(2014年2期)2014-02-28
