
隐翅虫成虫取食、防御和生殖行为
2013-06-11 10:29:18曹丹丹嵇保中刘曙雯陈晓明王丽平赵正萍冀士琳
环境昆虫学报 2013年6期
曹丹丹,嵇保中*,刘曙雯,陈晓明,王丽平,赵正萍,冀士琳
(1.南京林业大学森林资源与环境学院,南京 210037;2.南京中山陵园管理局,南京 210014)
隐翅虫科Staphylinidae 隶属于昆虫纲Insecta鞘翅目 Coleoptera 多食亚目 Polyphaga。林奈(Linnaeus)1758年最早使用隐翅虫属Staphylinus概括了当时全部的已知种类。Paykull 1798年出版了《瑞典隐翅虫》 专著,随后德国学者Gravenhorst(1802)发表了“棕色的隐翅虫”并出版《隐翅虫》一书(1806),Erichson(1840)在其名著《隐翅虫的属与种》中对该类昆虫做了比较完善的分类。继Erichson 后,有关隐翅虫新属新种的报道层出不穷,以不同的语言形式出现于各种刊物上,隐翅虫其它方面的研究也逐步开展(郑发科,1992)。
Ashe(1987)、Alcock(1991,1994)、Betz(1996,1998)、Detter(1985,1986,1987,1991ab)、Newton and Thayer(1992)、Peschke(1978,1982,1983,1986,1987ab,1990)等学者在1990年代前后就隐翅虫捕食行为、化学防御及生殖行为做了较为广泛研究,对象主要包括前角隐翅虫亚科Aleocharinae、异形隐翅虫亚科Oxytelinae、隐翅虫亚科Staphylininae、及突眼隐翅虫亚科Steninae。Frank和Thomas(2011)对隐翅虫的主要习性做了全面概括。
我国地域辽阔,跨古北、东洋两个区系,隐翅虫种类十分丰富,有农田害虫的捕食性天敌,也有生态平衡调节者等不同的功能类群。我们研究松梢害虫防治的过程中,在微红梢斑螟Dioryctria rubella Hampson 蛀道发现一隐翅虫新种松梢隐翅虫Placusa pinearum Gao,Ji,Liu,sp.nov.(Gao et al.,2011),该种隐翅虫与微红梢斑螟幼虫跟随性较强(高江勇等,2010)。随后,我们又发现松材线虫病死松树皮下存在较丰富的隐翅虫资源,这些隐翅虫时间、空间生态位宽度值很大,与天牛、小蠹、象甲等钻蛀性害虫的时间、空间生态位重叠值及生态位相似性比例指数较高(待发表)。上述研究表明,以隐翅虫为载体,携带病原微生物,防治林木钻蛀性害虫具有较大的潜力。行为习性的研究是资源开发利用的前提,本文评述国内外隐翅虫成虫取食行为、防卫行为及生殖行为的研究进展,以作为相关研究的参考。
1 取食行为
隐翅虫有较广的生境及食性,喜潮湿的地方,以小型昆虫、植物花粉、菌类为食,也有一些种类寄生在动物体内。除此之外还有药用种类、卫生害虫,以及取食动植物有机残体、促进自然界物质循环、保持生态平衡的腐食性种类,与人类关系密切,是一类不可忽视的自然资源(张艳,2004)。
1.1 食性
现存的隐翅虫类群的食性大致总结如表1。总体来讲隐翅虫食性复杂不易确定,通过口器和消化道解剖结构进行的比较形态学研究认为:原始类型的隐翅虫可能是腐食性的,由腐食性的原发类型(四眼隐翅虫亚科Omaliinae、异形隐翅虫亚科Oxytelinae)向活泼游掠的高级类型(毒隐翅虫亚科Paederinae、隐翅虫亚科Staphylinidae)的缓慢进化造成隐翅虫科(Staphylinidae)内各亚科食性的演变(Crowson,1960;Frank and Thomas,2011)。表1 中所列的隐翅虫食性与分类系统各类群的关系所反映的也只是大概趋势,目前研究所涉及的种类相对较少,有些种类隐翅虫种类食谱范围较宽,为杂食性,如普拉隐翅虫属Platydracus(Staphylininae)的某些种类(许旺,2009),即使同属隐翅虫的食性也不尽相同,如佳隐翅虫属Gabrius(Staphylininae)大多为捕食性种类,但也有些为花粉食性等(齐楠,2009)。因此,不同类群隐翅虫的食性还需要不断补充完善。
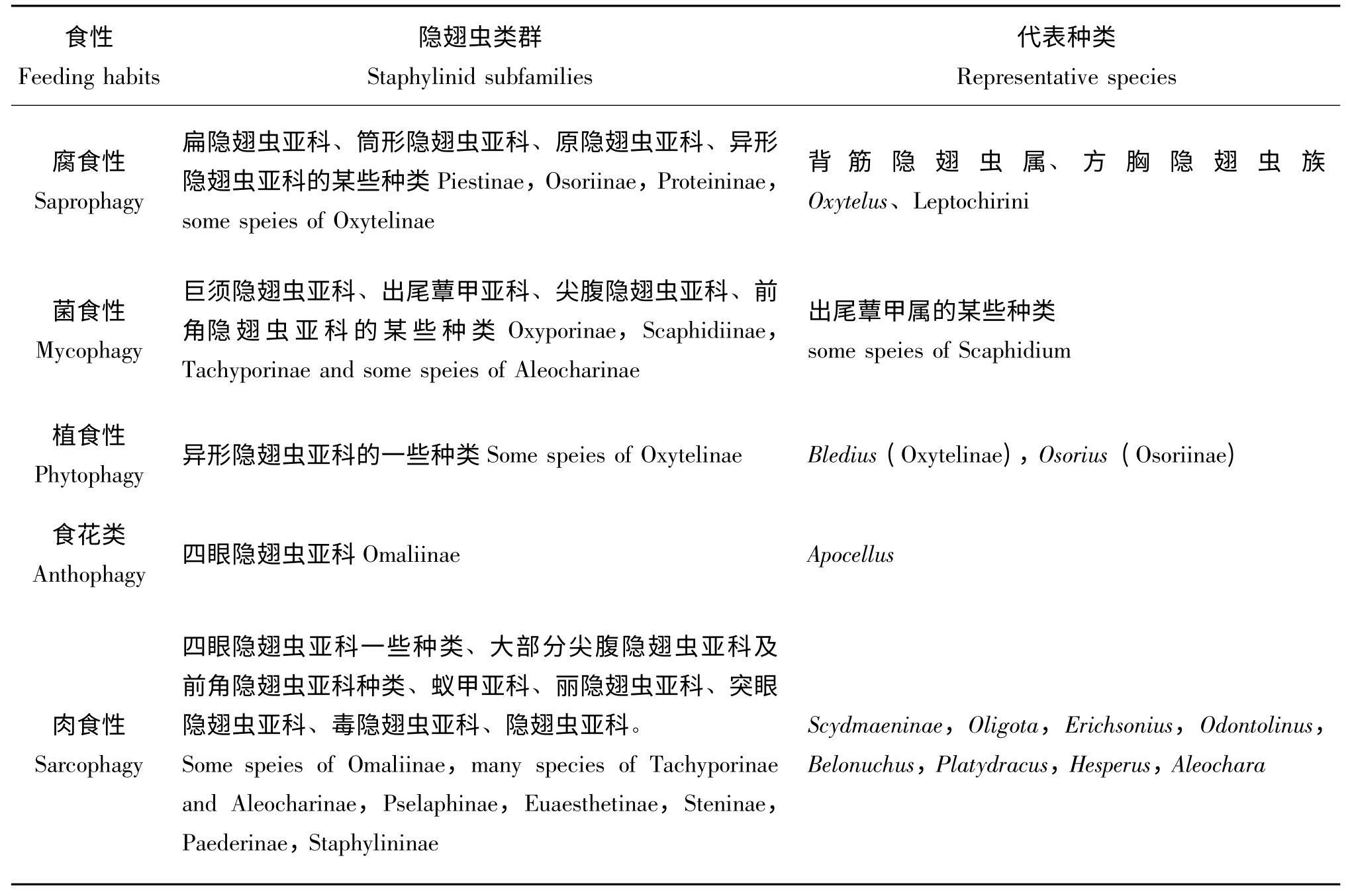
表1 隐翅虫食性Table 1 Feeding habits of staphylinid
1.2 捕食行为策略及取食方式
目前捕食性昆虫生态学方兴未艾,已成为现代生态学的一个重要内容。最优取食理论认为:对猎物密度变化信息的评估,是捕食者产生捕食策略的根本途径,或其与猎物相遇率的变化,或其肠内猎物含量的变化(戈峰,2008)。对于不同的捕食者,其策略形成的机理不同。有些捕食性的隐翅虫种类是农田害虫的重要生物控制因素,是一类应予以保护利用的天敌,如斑足突眼隐翅虫Stenus cicindela、黑胫突眼隐翅虫Stenus macies、黄足蚁形隐翅虫Paederus fuscipes 及脊胸小头隐翅虫Philonthus rutiliventris 等可捕食玉米螟、叶蝉、飞虱、蚜虫、蓟马等多种农业害虫。隐翅虫捕食行为的研究有助于制定有效的保护与利用天敌的方案。
1.2.1 捕食行为策略
隐翅虫常见的捕食策略有积极搜索方式、坐等捕食方式、搜索活动依猎物密度而变化的混合式等,如Ocypus olens 在觅食过程中有积极搜索行为(active searching behaviour)和埋伏等待行为(ambush-like behaviour)两种方式。热带雨林的Leistotrophus versicolor 会根据环境条件决定自己的捕食策略。在食物密度大的区域,个体落在脊椎动物的粪、腐肉等腐烂物质上,埋伏等待着接近腐肉的蝇(坐等捕食方式);但是当环境中没有腐烂物质时,成虫可停在树叶或岩石上,通过外翻肛门附近第8/9 背板腺体囊,分泌液滴于叶片或石头等基质上,引诱猎物(例如:小蝇)到可攻击范围内。这种行为属于种间化学欺骗引诱捕食,推断这些腺体分泌物与排泄物混合,含有信息素类似物(pheromone-mimics)或蝇类引诱物(lures of the flies)(Forsyth and Alcock,1990a)。
除化学引诱捕食外,有些隐翅虫种类与环境协同进化形成特殊的捕食器官。Stenus属成虫具有高度特化的下唇,在捕捉迅速逃跑猎物(例如:弹尾虫)的过程中,靠血淋巴的压力快速伸出,利用末端带有粘液的肉垫黏住猎物,并将其拖进上颚范围内取食(Betz,1996)。由于下唇捕食的长距离、高速度及瞬间固定猎物的能力,与用上颚捕食的种类相比,下唇能更成功的捕捉到跳虫(Betz,1998)。但是如果跳虫个体较大或者被鳞屑或刚毛覆盖,被捕对象不容易粘在下唇端部,则攻击成功率降低(Bauer and Pfeiffer,1991)。粘着固定作用由头壳内部的腺体产生并分泌到侧唇舌上的分泌物调节。这些粘腺(adhesive glands)由离散的腺体单元组成,每个单元包含3个细胞,其中两个细胞为分泌细胞,产生不同的分泌物,一个产生蛋白类物质,另一个产生类脂。所以,在侧唇舌的表面会发现两类分泌物。粘腺与一般的表皮腺体同源,但由于行使不同的功能导致结构上的分化(Kölsch,2000)。
1.2.2 取食方式
捕食性隐翅虫存在两种取食方式,一种为普通的撕碎-咀嚼,另一种为捕捉-口前消化(preoral digestion)的方式。梭毒隐翅虫 Paederus fuscipes 捕食蚜虫的行为包括搜寻、捕捉、嚼食、梳理、静止和排泄6个过程(李霞,2002)。Bonacci 等(2006)对Ocypus olens 的研究表明,上颚在捕捉、固定及撕碎食物方面发挥重要作用。当受到干扰的时候,O.olens 翘起腹部并张开上颚,在短暂追击碰到猎物时,用有力的上颚抓住猎物的前胸背板-腹部或者头部-前胸背板关节,猎物静止不动(假死反应),隐翅虫紧咬住前胸背板-腹部关节,旋转90°后用第一和第二对足固定食物,然后保持这种姿势至取食结束。与前述上颚撕咬、切碎猎物的种类相比,Philonthus decorus 等采用捕捉-口前消化(pre-oral digestion)的方式。P.decorus 的上颚发展成捕捉器官,在分开的上唇之间抓牢食物。下唇-咽下部(labium-hypopharynx)有浓密的刚毛覆盖,上唇中间有一条食物沟槽,食物进入口腔后部混入消化液后,返回到口腔前部由上颚重新咀嚼,在此过程中形成的消化产物由强有力的食窦泵吸入前肠,食物残渣则排出口腔外部(Evans,1963)。与P.decorus取食方式类似,Oligota kashmirica benefica 捕食时,将猎物六点黄蜘蛛Tetranychus urticae 揉搓成团,食物团在口腔后部与前肠分泌的消化液混合后,返回口腔前部由上颚再次咀嚼,该过程重复直到消化吸收完成。由于螨类个体小,咀嚼过程中体壁难以破碎,取食结束排出口腔的多为较完整的猎物体壳(Nield,1976;Shimoda et al.,1997)。
巨须隐翅虫亚科Oxyporinae、大眼隐翅虫亚科Steninae、丽隐翅虫亚科Euaesthetinae、毒隐翅虫亚科Paederinae和隐翅虫亚科Staphylininae 的一些种类均有捕捉-口前消化的取食方式。在此过程中,用于口前消化的消化液来源于前肠,部分消化的食物进入消化道进一步消化,消化道内容物多数为液体,另混合一些经初步消化的食物成分,所以采用解剖镜检一般难以辨别食物种类,进而判断食性,这可能也是许多隐翅虫食性研究的难点所在(Frank and Thomas,2011)。
2 防御行为
修长的身体,狭短的鞘翅、柔韧灵活的腹部使隐翅虫适应土壤、枯枝落叶、树皮及真菌缝隙中的自由活动,形成较强的扩展生存领域优势。但与腹部完全被鞘翅覆盖的甲虫相比,裸露的腹部更容易受到捕食者的攻击及各种有害生物的侵染。隐翅虫常见的防卫行为包括行为防卫(逃遁)、物理防卫(伪装、形态及颜色拟态)及化学防卫等,其中化学防卫由腹部防卫腺体及多种具有高度生物活性的防卫信息化合物完成,研究较为深入(Dettner,1987)。
2.1 防卫腺体
隐翅虫腹部末端常有一对刺状突起,被干扰时或受惊时,能快速奔跑并将腹部末端翘起释放分泌物,分泌物的产生和储存通常由腹部末端背侧面的某些腺体完成(Jenkins,1957;Happ and Happ,1973;Dettner,1993;Klinger and Maschwitz,1977;Kanehisa et al.,1984)。隐翅虫防卫腺体囊(gland reservoirs)的着生位置(具体见表2)及形态结构差别很大,所分泌的化学物质也有所不同(Huth and Dettner,1990)。当隐翅虫与其它社会性昆虫一起生活时(如一些螱客与蚁冢隐翅虫),来自捕食性天敌及环境中病原微生物侵害的生存压力减弱,则典型的防卫腺体不同程度退化或完全消失(Kistner,1979;Shower and Kistner,1977)。
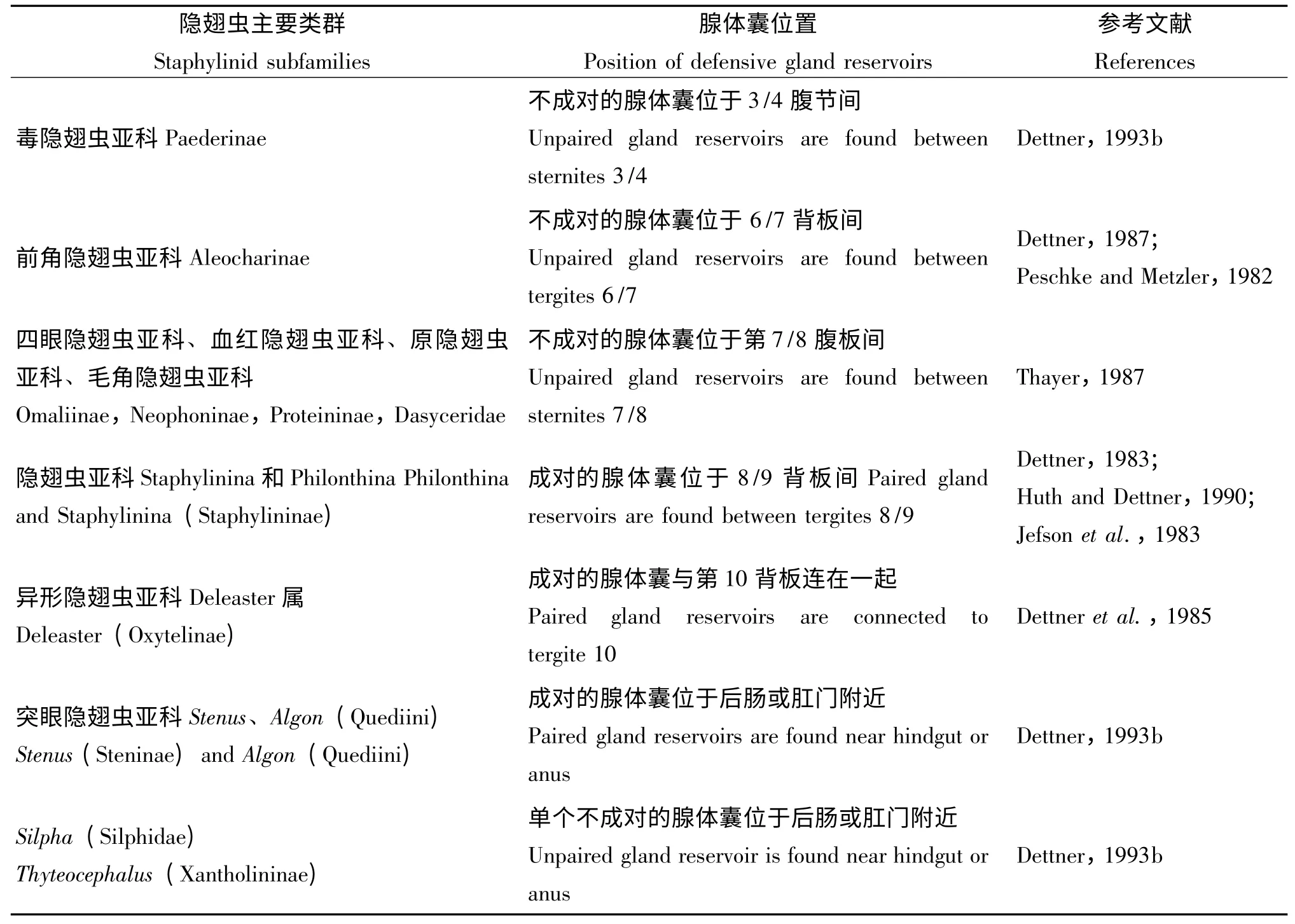
表2 隐翅虫防卫腺体囊位置Table 2 Position of defensive gland reservoirs
2.2 防卫信息化合物(defensive semiochemicals)
Abou-Donia 等(1971)发现Ocypus olens 防卫腺体分泌物与合成的琉蚁二醛一致,后经Fish和Pattenden(1975)鉴定该分泌物的主要挥发成分是4-甲基-3-己酮。Wheeler 等(1972)发现γ-丁位十二内酯是Bledius属防卫腺体系统的主要成分。此后,隐翅虫防卫信息化合物的研究快速发展。目前已有研究的隐翅虫不同类群的防卫性信息化合物鉴定情况统计如表1。醌类主要存在于前角隐翅虫亚科Aleocharinae、异形隐翅虫亚科Oxytelinae;环烯醚萜类主要存在于隐翅虫亚科Staphylininae、胸片隐翅虫亚科Xantholininae;酸类和alkenals 常见于四眼隐翅虫亚科Omaliinae、原隐翅虫亚科Proteininae 中;此外还有高度专化的物质:突眼隐翅虫亚科Steninae 臀板腺体中产生的哌啶衍生物stenusine、3-(2-甲基-1-丁烯基)嘧啶以及cicindeloine 可以迅速蔓延水面,帮助其逃离(Schildknecht et al.,1976;Dettner,1987;Schierling et al.,2012)。
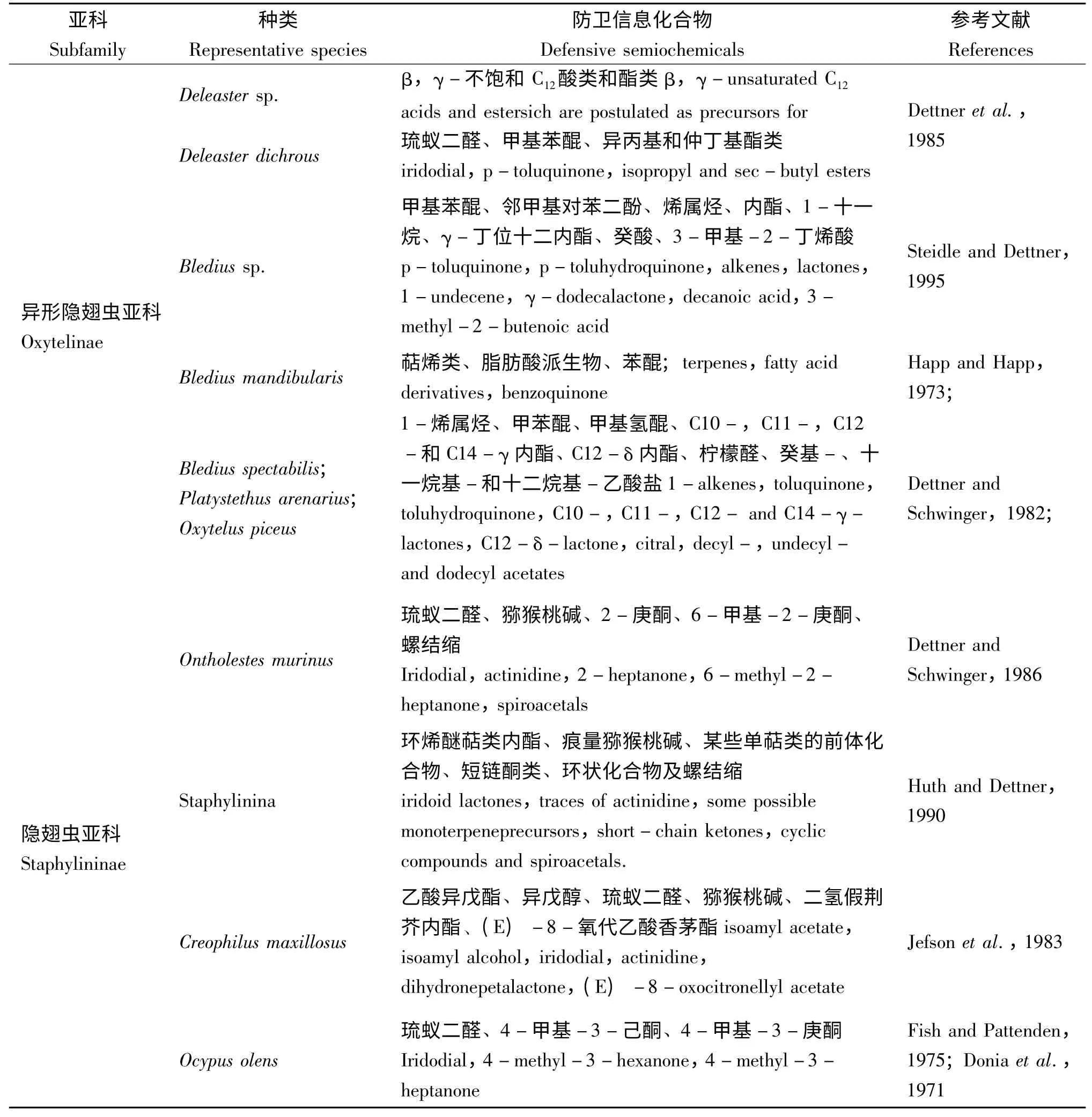
表3 隐翅虫主要代表种类的防卫分泌物Table 3 Defensive semiochemicals of Staphylinid representatives
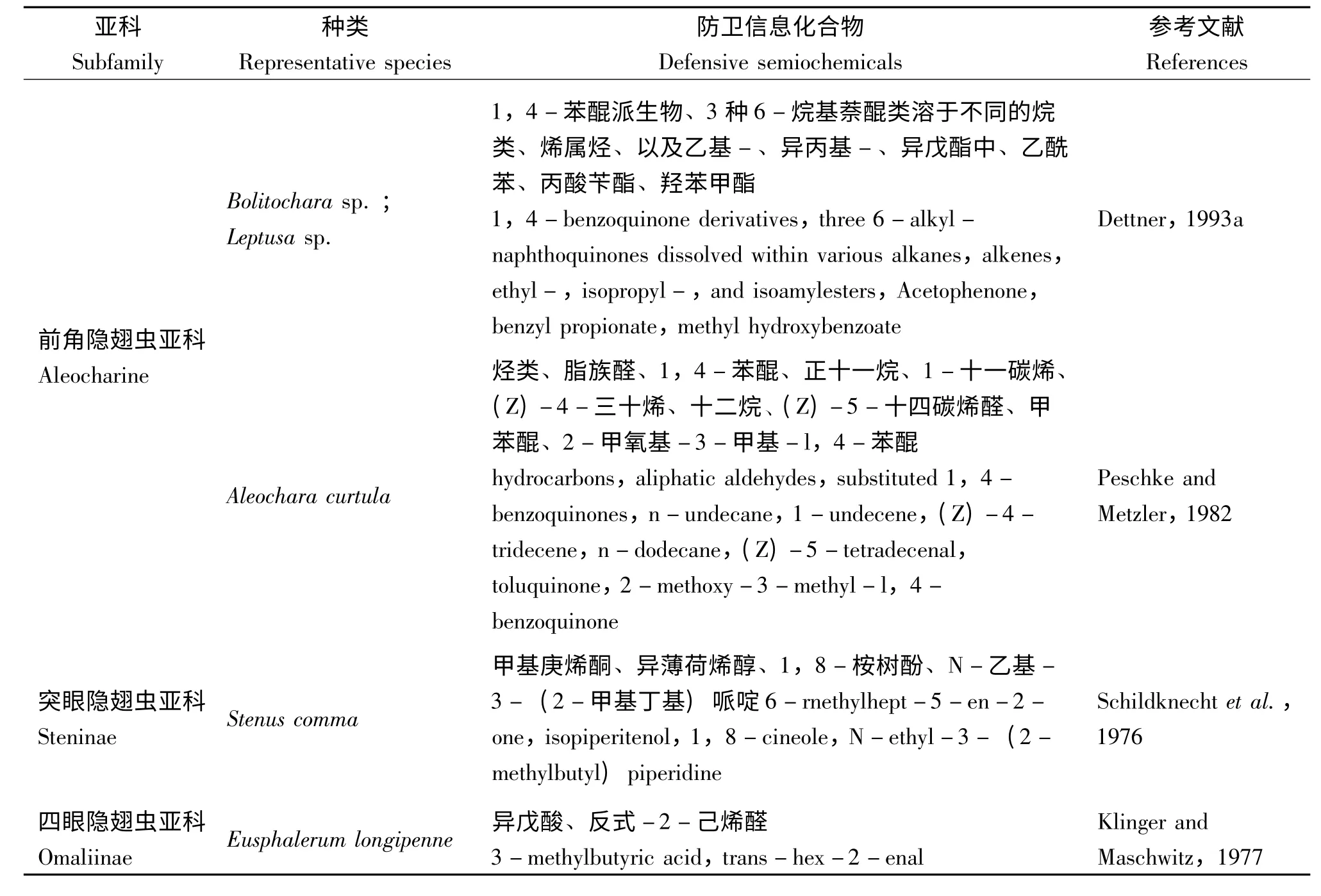
(续上表)
隐翅虫防卫信息化合物可以以液滴形式形成于防卫腺囊的开口处(异形隐翅虫亚科Oxytelinae,前角隐翅虫亚科Aleocharinae,四眼隐翅虫亚科Omaliine),也可以将腺囊部分外翻,湿润虫体(Badgley,1956;Dettner,1991ab),或者通过腹部的活动瞄准靶标生物Algon grandicollis(Kanehisa et al.,1984)和某些腐尸甲虫,有时甚至用防卫分泌物直接喷雾,如Bolitochara Leptusa(Eisner and Meinwald,1982)。
某些隐翅虫还可以模拟其它物种的化学信息物质而进行自身保护。喜与蚁同居的Pella属隐翅虫Pella funestus和P.humeralis 的分泌物包括醌类及不同的脂肪族化合物,如正十一烷和6-甲基-5-庚烯-2-酮(甲基庚烯酮)等。行为试验证明十一烷是对“侵略警报”产生诱导作用的信息素,6-甲基1-5 庚烯-2 酮是对“恐慌警报”产生诱导作用的背部腺体化合物,而这两种物质也存在于Lasius fuliginosus(Hymenoptera:Formicidae)信息素腺体中,Pella属隐翅虫通过模拟共栖昆虫的警报信息素避免受到攻击(Dettner,1993;Stoeffler et al.,2007)。
2.3 防卫信息化合物的多效应现象(multifunction)
大部分昆虫的信息化合物具有简约性(parsimony),简约性一般与多效性相联系,即一种信息化合物具有多种功能(多效应现象),从而使信息化合物的生物合成途径、器官或原料等方面节省。隐翅虫防卫信息素可以防御捕食者,还可以防御病原微生物的侵害,同时通过释放量的变化对交配行为产生影响,因此具有明显的简约性。Peschke(1983)研究表明Aleochara curtula 高浓度的背板腺分泌物(tergal gland secretion)能抑制雄虫的交配反应,起抑制作用的化学成分被认为是警报物质,如正十一烷、1-十一碳烯、正十二醛、甲苯醌、2-甲氧基-3-甲基-1,4-苯醌。而当隐翅虫受到轻微干扰的时候,背板腺分泌物的分泌量较小,交配行为明显增强。对于交配行为起催情作用的主要成分包括(Z)-4-十三烯、n-十二醛、(Z)-5-tetradecenal。如果雌性严重受伤或者释放大量防卫分泌物,雄性则不会表现出性行为,而且会远离这样的雌性并净化触角(Peschke and Metzler,1982;Peschke,1983)。
防卫信息化合物在雄性性内竞争过程中也扮演重要角色。为了接近雌性,获得更多的交配机会,雄性A.curtula 在其聚集地通过格斗行为垄断资源,保卫其领地,互相撕咬足和触角以及用其灵活的腹部攻击对手(Peschke,1987ab)。在这些斗争中,背部第6/7 腹节节间褶上的防卫腺体都会释放防卫信息化合物:有毒的2-甲基-1,4 苯醌、2-甲氧基-3-甲基-1,4 苯醌以及一些脂族醛和烃类,如n-十二醛、(Z)-5-tetradecenal、正十一烷、1-十一碳烯及(Z)-4-十三烯(Peschke and Metzler,1982)。其中2-甲基-1,4 苯醌作为分泌物的背板腺分泌物的目标化合物,释放量在雄性竞争过程中逐步增加。由于雄性信息素、防卫信息化合物两者的生物合成相互关联,而雌性可能根据雄性信息素判断雄虫品质,进而选择交配对象,所以防卫信息化合物释放量影响交配选择(Boppré and Schneider,1989;Holz et al.,1994;Peschke et al.,1996)。
3 隐翅虫生殖行为
3.1 交配行为与性信息素
大眼隐翅虫Stenus(s.str.)sp.交配行为包括相遇、抱对、锁结、离开等几个过程(郑英奎等,2009)。A.curtula 雄性的性行为在羽化2 周后发生,而雌性性信息素的产生在羽化前和羽化后的几天内迅速增加。雌雄虫胸部和腹部的信息素含量相当,并经体外实验与身体碎片实验表明,表皮上分散的腺体会产生性信息素,经鉴定属于表皮烃的组分(Peschke,1986)。关于其它种类隐翅虫生殖行为方面的研究鲜见报道,只局限于A.curtula(Peschke,1987ab;Gack et al.,1994;;Peschke et al.,1999;Schlechter-Helas,et al.,2011;Schlechter-Helas,et al.,2012)、L.versicolor(Forsyth and Alcock,1988;Forsyth and Alcock,1990b)、Ontholestes cingulatus、Aleochara bilineata(Lizé et al.,2009)等几个代表种类。
而关于隐翅虫性信息素方面,目前只有A.curtula 的雌性性信息素得到分离鉴定。A.curtula通过背部防卫腺体分泌物组分进行种内信息交流,同时还起到刺激交配的作用(Peschke,1982),其雌性信息素来源于上表皮的酯类(Peschke,1978)。Peschke和Metzler(1986)认为表皮烃成分对A.curtula 交配识别、雄性侵略以及雌性选择起调节作用。经鉴定A.curtula 雌性性信息素的主要组分为(Z)7-二十一烯和(Z)7-二十三烯(诱虫烯)。在聚集场所,雄性在短距离(大约2 mm)内通过触角的嗅觉感受器感知低挥发性的雌性信息素,引起交配反应(Peschke and Metzler,1987)。野外试验中,栖息着隐翅虫的腐肉会产生特异的引诱信息素(Peschke et al.,1999),同种的雌雄性个体会被吸引并将其作为交配场所(Peschke et al.,1987a),Isopropyl(Z9)-hexadecenoate 及活体雄性分别与新鲜的腐肉味结合,均比仅用新鲜腐肉能吸引更多的隐翅虫。表明隐翅虫性信息素通讯中,腐烂物质的挥发性成分起到很大的辅助作用,雌虫可以通过腐烂物的气味找到雄虫,一旦交配成功,雌性还可就近产卵,这与苹果蠹蛾雌虫在有苹果气味存在时提高性信息素的释放量,并将求偶时间提前的情况相似(戈峰,2008)。
3.2 雄性交配权竞争(mating right competition)
大部分隐翅虫种类多次交配,雄性个体表现出较强烈的交配权竞争。通常情况下,雌性个体耐心选择配偶,以便占有优质遗传资源,而雄虫的交配权竞争主要表现为对食物及交配对象的竞争。L.versicolor 成虫聚集在脊椎动物的粪和腐尸上,以双翅目成虫为食。雄性互相斗争排斥,有明显的领地行为(territorial behavior)(个体所占据并积极保护不让其它成员侵入),胜利者可以获得资源的垄断权,在雌虫接近资源时,雄虫则获得交配的机会。雄虫的上颚因用于争斗而特别发达,同时个体较大的雄性在竞争中往往具有优势。而一些体型小的个体,或者用上颚争斗不能获胜的个体,则会分泌与雌性分泌物类似的信息化合物,避免雄性之间的斗争,以继续留在粪上搜寻食物,有时甚至会获得雄性竞争对手的示爱(Forsyth and Alcock,1990)。这种“种内化学信息欺骗”也存在于A.curtula种类中,发育不成熟或饥饿的雄虫会模拟产生雌性的信息素组分,以避免雄性之间的斗争,成功接近取食和交配场所(Peschke,1985;1987b)。
3.3 精子传递方式及精子竞争机制
Parker 于 1970年提出精子竞争(sperm competition)的概念,有多次交配习性的雌虫,交配后,来自不同雄性的精子为了争夺对卵的授精权而展开的竞争,是生殖竞争的最后阶段。精子竞争的模式包括最先雄虫精子优先、最后雄虫精子优先和无雄虫精子优先(滕兆乾和张青文,2006)。精子的竞争机制主要有:(1)精子失能:交配雄性通过其精子形态上的适应或精液生理上的适应而杀死或抑制雌性储精器官内先前存储的精子(Diesel,1990);(2)精子移除:雄性用类似刷子的结构或特殊的生殖器构造(Helversen and Helversen,1991)将雌性储精器官中先前存储的精子移除,是雄性生殖器的形态学适应(Waage 1979,1982;Miller and Miller,1981;Yokoi,1990);(3)精子层化:来自不同雄性的精子成层布局在雌性的储精器官中,雌性产卵时最后交配雄性的精子在受精时先排出储精器官,造成“最后雄性授精优势”(Waage,1984;滕兆乾和张青文,2006);(4)精子冲洗(精子取代):之前交配的雄性的精包(精液)可能被随后交配雄性个体的精液冲出雌性储精器官或被挤压到受精囊的边缘(Smith,1979ab;Ono et al.,1989);(5)雌性的隐秘选择(cryptic female choice):一个雌性可与多个雄性交配,雌性可控制雄性精子在受精囊中的空间分布、受精过程中精子的使用情况以及交配中雄性所输送精子的数量,然后对精子进行有选择性的受精(王会琴和李金钢,2011)。
在隐翅虫中,最后交配雄虫精子优先及精子冲洗机制已得到验证。A.curtula 雌虫具有“U”形的受精囊,受精囊具有瓣阀构造和两个骨化齿,交配时在鞭状的雄虫内阳茎引导下,精包管伸入受精囊管。交配拆对后,精包管进一步伸长,在经过受精囊瓣阀时初生精包管(primary tube)的顶端破裂,在初生精包管内形成的次生精包管(second tube)穿过瓣阀继续前伸到球形的受精囊体内,后弯并膨大呈球形,充实受精囊体腔大部。受精囊肌肉挤压使受精囊内壁的齿状构造刺穿精包体,释放精子,冲洗前雄性交配者遗留的精子,实现精子替代(Gack and Peschke,1994;Förster et al.,1998)。
雄性为了在精子竞争中取得胜利,发展了形态、生理、行为等方面的适应机制。A.curtula(Gack and Peschke,1994)、O.cingulatus(Alcock,1991)、L.versicolor(Alcock and Forsyth,1988)和Eumicrota属(Ashe,1987)雄性延长交配时间及交配后的“保卫配偶(Mate-Guarding)”行为强化精子竞争(Parker,1970)。一般认为,“阻止雌性再次交配”、“增加精子输入量”和“雄性通过延长交配时程评估雌性身体质量状况”等是隐翅虫的延长交配时间的原因(Alcock,1994),而交配后“保卫配偶”行为则使雄虫能最大程度为卵授精,实现最大的繁殖价值。“保卫配偶”行为的代价在于处于保卫期的雄性不能寻找其它雌性并与之交配,根据进化理论,当生殖利益大于其付出代价时,如可交配的雌性少、雌性多次交配或驱赶其它竞争者使自己的精子为交配后的雌性授精,雄虫才会有该行为。
在交配期间A.curtula 雄性将抑性信息素(antiaphrodisiac pheromone)通过精包输入雌虫生殖腔,进而扩散到雌性体表,降低其吸引力,阻止与其它雄性交配,该行为被视为隐翅虫对精子竞争适应的生理适应机制。与短距离外可被察觉的挥发性的性信息素相比,抑性信息素只有通过阳基侧突的接触才能察觉到。所以,刚刚交配过的雌性对于其它雄性来讲,不是很适宜的交配对象。由于其生殖腔被精包堵塞,雌性通过摆动腹部拒绝雄性的求爱(Schlechter-Helas et al.,2011),减小了精子竞争的风险,同时雄性利用表皮烃识别个体,不会再接触交配过的雌性,以避免时间和能量的浪费,而雌虫也可以避免来自其它雄虫的侵扰。
4 结语
隐翅虫资源丰富,分布广泛,是鞘翅目中的一大类群,在生物多样性和生态平衡中占有重要地位。隐翅虫食性复杂,捕捉-口前消化的方式是捕食性隐翅虫常见的取食方式,Stenus属隐翅虫进化形成了特殊的捕食器官(stick-capture apparatus)(Koerner et al.,2012);隐翅虫口器和消化道的构造有一定的系统分类学意义,一定意义上说明其构造与功能的适应性进化较为快速,同时也说明在进化历史上,隐翅虫可能存在食物范围、生存空间不断扩大的扩张时期。毒隐翅虫亚科、前角隐翅虫亚科、异形隐翅虫亚科、突眼隐翅虫亚科、隐翅虫亚科、四眼隐翅虫亚科、血红隐翅虫亚科、原隐翅虫亚科、毛角隐翅虫亚科、胸片隐翅虫亚科单个或成对的防卫腺体囊位于腹部第6~10 背板,并有逐渐向末端移动的趋势(Dettner,1993),目前,已分离鉴定的防卫信息化合物主要包括醌类、内酯、胺类、酮类、醛类、烷类及烯属烃类等(Abou-Donia et al.,1971;Wheeler et al.,1972;Fish and Pattenden,1975;Schildknecht et al.,1976;Peschke and Metzler,1982;Dettner,1987;Steidle and Dettner,1995)。隐翅虫生殖行为研究主要侧重于交配保护“Mate-Guarding”行为(Alcock and Forsyth,1988;Alcock,1991;Gack and Peschke,1994)以及雄性精子竞争行为,多次交配的个体最后交配的雄虫精子优先使用的机制,即精子冲洗机制,在A.curtula中已经得到验证(Gack and Peschke,1994;Förster et al.,1998)。A.curtula 的雌性性信息素(诱虫烯)也已分离鉴定((Z)7-二十一烯和(Z)7-二十三烯)(Peschke,1978、1982;Peschke and Metzler,1986),其它种类的性信息素研究急待加强。
目前,除隐翅虫防卫行为的研究较为广泛外,其它行为的研究均较为薄弱,主要体现在以下几个方面:(1)隐翅虫的基础研究工作仍需进一步加强,目前许多种类基本的生物生态学特性知之甚少,大部分种类隐翅虫食性及捕食机制都不明确;(2)隐翅虫生殖行为的研究处于起步阶段,目前除A.curtula 之外,其它种类的性信息素均未分离鉴定,交配行为机制也不明确;(3)隐翅虫各种行为(如寄主选择、捕食、防御、生殖行为)之间化学信息交流机制的研究需要进一步深入,信息化合物的分离鉴定工作也亟待加强。随着现代研究方法和技术条件的不断进步以及隐翅虫经济意义的提高,上述研究可望取得更快的进展。
References)
Abou-Donia SA,Fish LJ,Pattenden G,1971.Iridodial from the odoriferous glands of Staphylinus olens(Coleoptera,Staphylinidae).Tetrahedron.Lett.,43:4037-4038.
Alcock J,1991.Adaptive mate-guarding by males of Ontholestes cingulatus(Coleoptera:Staphylinidae).J.Insect.Behav.,4(6):763-771.
Alcock J,1994.Postinseminational associations between males and females in insects:the mate guarding hypothesis.Ann.Rev.Entomol.,19:1-21.
Alcock J,Forsyth A,1988.Post-copulatory aggression toward their mates by males of the rove beetle Leistotrophus versicolor(Coleoptera: Staphylinidae).Behav.Ecol.Sociobiol.,22:303-308.
Ashe JS,1984.Major features of the evolution of relationships between gyrophaenine staphylinid beetles(Coleoptera: Staphylinidae:Aleocharinae)and fresh mushrooms,227-255.In Wheeler Q,Blackwell M,eds.Fungus-insect Relationships;Perspectives in Ecology and Evolution.New York:Columbia University Press.
Ashe JS,1987.Egg chamber production,egg protection and the clutch size among fungivorus beetles of the genus Eumicrota(Coleoptera:Staphylinidae) and their evolutionary implications.Zool.J.Linn.,90:255-273.
Badgley ME.1956.Biology of Ohgota oviformis Casey(Coleoptera:Staphylinidae).Ann.Entomol.Soc.Am.,49:501-502.
Bauer T,Pfeiffer M,1991.‘Shooting’springtaiis with a sticky rod:the flexible hunting behaviour of Stenus comma(Coleoptera:Staphylinidae)and the counter-strategies of its prey.Anim.Behav.,41:819-828.
Benken T,Knaak A,Gack C,Eberle M,Peschke K,1999.Variation of sperm precedence in the rove beetle Aleochara curtula(Coleoptera:Staphylinidae).Behaviour,136,1065-1077.
Betz O,1996.Function and evolution of the adhesion-capture apparatus of Stenus species(Coleoptera,Staphylinidae).Zoomorphology,116:15-34.
Betz O,1998.Comparative studies on the predatory behaviour of Stenus spp.(Coleoptera:Staphylinidae):the significance of its specialized labial apparatus.J.Zool.Lond.,244(4):527-544.
Betz O,Thayer MK,Newton AF,2003.Comparative morphology and evolutionary pathways of the mouthparts in spore-feeding Staphylinoidea(Coleoptera).Acta Zoologica.,84(3):179-238.
Bonacci T,Massolo A,Brandmayr P,Brandmayr TZ,2006.Predatory behaviour on ground beetles(Coleoptera:carabidae)by Ocypus Olens(MüLLER)(Coleoptera:Staphylinidae) under laboratory conditions.Ento.News.,117(5):545-551.
Boppré M,Schneider D,1989.The biology of Creatonotos(Lepidoptera: Arctiidae) with special reference to the andoroconical system.J.Linn.Soc.,96:339-356.
Chang YD,Kang L,2002.Strategies used by male and female insects in multiple mating behavior and sperm competition.Acta Entomologica Sinica,45(6):833-839.[长有德,康乐,2002.昆虫在多次交配与精子竞争格局中的雌雄对策.昆虫学报,45(6):833-839]
Chatzimanolis S,2003.Natural history and behavior of Nordus fungicola(Sharp)(Coleoptera:Staphylinidae).Ann.Entomol.Soc.Am.,96(3):225-230.
Crowson RA,1960.The phylogeny of Coleoptera.Annu.Rev.Entomol.,(5):111-134.
Dettner K,Schwinger G,1982.Defensive secretions of three Oxytelinae rove beetles(Coleoptera:Staphylinidae).J.Chem.Ecol.,8(11):1411-1422.
Dettner K,Schwinger G,Wunderle P,1985.Sticky secretion from two pairs of defensive glands of rove beetle Deleaster dichrous(Grav.)(Coleoptera:Staphylinidae)gland morphology,chemical constituents,defensive functions,and chemotaxonomy.J.Chem.Ecol.,11(7):859-885.
Dettner K,Schwinger G,1986.Volatiles from the defensive secretions of two rove beetle species(Coleoptera,Staphylinidae).Z.Naturforsch.,41:366-368.
Dettner K,1987.Chemosysternatics and evolution of beetle chemical defenses.Annu.Rev.Entomol.,32:17-48.
Dettner K,1991a.Dabbing and shotting of benzo-and naphthoquinone secretions:the defensive strategies of bark inhabiting Aleocharine(Col.:Staphylinidae)and Tenebrionid(Col.:Tenebrionidae)beetle larvae.J.Chem.Ecol.,44:50-58.
Dettner K,1991b.Solvent-dependent variability of effectiveness of quinone-defensive systems of Oxytelinae beetles(Coleoptera:Staphylinidae).Ent.Gen.,15:275-292.
Dettner K,1993.Dabbing and shooting of benzo-and naphthoquinone secretions:defensive strategies of barking-inhabiting Aleocharine(Col.:Staphylinidae)and Tenebrionid(Col.:Tenebrionidae)beetle larvae.J.Chem.Ecol.,19(7):1337-1356.
Diesel R,1990.Sperm competition and reproductive success in the decapod Inachus phalangium(Majidae):a male ghost spider crab that seals off rival's sperm.J.Zool.Lond.,220:213-223.
Eisner T,Meinwald J,1982.Defensive spray mechanism of a silphid beetle(Necrodes surinamensis).Psyche.,89(3-4):357.
Evans MEG,1963.A comparative account of the feeding methods of the beetles Nebria brevicollis(F.)(Carabidae)and Philonthus decorus(Grav.)(Staphylinidae).Trans.R.Soc.Edinb.,66:91-109.
Fish LJ,Pattenden G,1975.Iridodial and a new alkanone,4-methyl-hexan-3-one,in the defensive secretion of the beetle Staphylinus olens.J.Insect.Physiol.,21(4):741-744.
Förster M,Gack C,Peschke K,1998.Function and morphology of the spermatophore of the rove beetle,Aleochara curtula.Zoology,101:34-44.
Forsyth A,Alcock J,1990a.Ambushing and prey-luring as alternative foraging tactics of the fly-catching rove beetle Leistotrophus versicolor(Coleoptera:Staphylinidae).J.Insect Behav.,3(6):703-720.
Forsyth A,Alcock J,1990b.Female mimicry and resource defense polygyny by males of a tropical rove beetle,Leistotrophus versicolor(Coleoptera: Staphylinidae).Behav.Ecol.Sociobiol.,26:325-330.
Frank JH,Barrera R,2010.Natural history of Belonuchus Nordmann spp.and allies(Coleoptera:Staphylinidae)in Heliconia L.(Zingiberales:Heliconiaceae)flower bracts.Insecta Mundi,10:1-12.
Frank JH,Thomas MC,2011.Rove beetles of the world,Staphylinidae(Insecta:Coleoptera:Staphylinidae).University of Florida.IFAS Extension,114:1-8.
Gack C,Peschke K,1994.Spermathecal morphology,sperm transfer and a novel mechanism of sperm displacement in the rove beetle,Aleochara curtula(Coleoptera,Staphylinidae).Zoomorphology,114:227-237.
Gao JY,Ji BZ,Liu SW,2011.A new species of Placusa Erichson(Coleoptera,Staphylinidae,Aleocharinae)from China.Zootaxa,3094:43-51.
Gao JY,Ji BZ,Liu SW,Wang GX,Kuang MH,Gao YG,2010.Population composition and niche of arthropod community in pine shoot tunnel bored by Dioryctria rubella.Chinese Journal of Ecology,29(2):363-369.[高江勇,嵇保中,刘曙雯,王国兴,况美华,高玉国,2010.微红梢斑螟蛀道节肢动物种群结构及生态位.生态学杂志,29(2):363-369]
Gao JY,Ji BZ,Liu SW,2011.A new species of Placusa Erichson(Coleoptera,Staphylinidae,Aleocharinae)from China.Zootaxa,3094:43-51.
Ge F,2008.Principle and Method of Insect Ecology.BeiJing:Higher Education Press.[戈峰,2008.昆虫生态学理论与方法.北京:高等教育出版社]
Gnanasunderam C,Young H,Utcher CFB,Hutchins RFN,1981.Ethyl decanoate as a major component in the defensive secretion of two New Zealand Aleocharine(Staphylinidae) Beetles-Tramiathaea cornigera(Broun) and Thamiaraea fuscicornis(Broun).J.Chem.Ecol.,7(1):197-204.
Gregor K,2000.The ultrastructure of glands and the production and function of the secretion in the adhesive capture apparatus of Stenus species(Coleoptera:Staphylinidae).Can.J.Zool.,78:465-475.
Happ GM,Happ CM,1973.Fine structure of the pygidial glands of Bledius mandibularis(Coleoptera:Staphylinidae).Tissue and Cell,5(2):215-231.
Helversen D,Helversen O,1991.Pre-mating sperm removal in the bushcricket Metaplastes ornatus Ramme 1931(Orthoptera,Tettigonoidea,Phaneropteridae).Behav.Ecol.Sociobiol.,28:391-396.
Holz C,Streil G,Dettner K,Dütemeyer,Boland W,1994.Intersexual transfer of a toxic terpenoid during copulation and its parental allocation to developmental stages:quantification of cantharidin in cantharidin-producing oedemerids(Coleoptera:Oedemeridae) and cantharophilous pyrochroids(Coleoptera:Pyrochroidae).Z.Naturforsch.,49(11):856-864.
Huth A,Dettner K,1990.Defense chemicals from abdominal glands of 13 rove beetle species of Subtribe Staphylinina(Coleoptera:Staphylinidae,Staphylininae).J.Chem.Ecol.,16(9):2691-2712.
Jefson M,Meinwald J,Nowicki S,Hicks K,Eisner T,1983.Chemical defense of a rove beetle(Creophilus maxillosus).J.Chem.Ecol.,9(1):159-182.
Jenkins MS,1957.The morphology and anatomy of the pygidial glands of Dianous coerulescens(Coleoptera:Staphylinidae).Proc.R.Entomol.Soc.Lond.,Ser.A.,32:159-168.
Kanehisa K,Shiraga T,Wawazu K,1984.Defensive secretory organs of the rove beetles(Coleoptera:Staphylinidae).Nogaku Kenkyu,60:111-121.
Kistner DH,1979.Social and evolutionary significance of social insect symbionts.In Social Insects vol.1.Hermann HR,ed.339-413.New York:Academic Press.
Klinger R,Maschwitz U,1977.The defensive gland of Omaliinae(Coleoptera:Staphylinidae)I.Gross morphology of the gland and identification of the scent of Eusphalerum longipenne Erichson.J.Chem.Ecol.,3(4):401-410.
Koernera L,Gorbb SN,Betza O,2012.Functional morphology and adhesive performance of the stick-capture apparatus of the rove beetles Stenus spp.(Coleoptera,Staphylinidae).Zoology,115:117-127.
LI X,2002.Preliminary Study for Predatory Behavior of Adults of Paederus fuscipes Curtis.Journal of Sichuan Teachers College(Natural Science),23(1):11-14.[李霞,2002.梭毒隐翅虫成虫捕食行为初步研究.四川师范学院学报(自然科学版),23(1):11-14]
Lizé A,Cortesero A,Poinsot D,Boivin G,2009.Mating strategy in Aleochara bilineata: sperm storage and allocation.Physiol.Entomol.,34:129-135.
Miller PL,Miller CA,1981.Field observations on copulatory behaviour in Zygoptera,with an examination of the structure and activity of the male genitalia.Odonatologica,10:201-218.
Newton AF,Thayer MK,1992.Current classification and family-group names in Staphyliniformia(Coleoptera).Fieldiana,67:1-92.
Nield CE,1976.Aspects of the biology of Staphylinus olens(Müller),Britain's largest Staphylinid beetle.Ecol.Entomol.,1(2):117-126.
Ono T,Siva-Jothy MT,Kato A,1989.Removal and subsequent ingestion of rival's semen during copulation in a tree cricket.Physiol.Entomol.,14:195-502.
Parker GA,1970.Sperm competition and its evolutionary consequences in the insects.Biol.Rev.,45:525-567.
Peschke K,1978.The female sex pheromone of the staphylinid beetle,Aleochara curtula.J.Insect.Physiol.,24:197-200.
Peschke K,Metzler M,1982.Defensive and pheromonal secretion of the tergal gland of Aleochara curtula(Goeze)I.The chemical composition.J.Chem.Ecol.,8(4):773-785.
Peschke K,1983.Defensive and pheromonal secretion of the tergal gland of Aleochara curtula II.Release and inhibition of male copulatory behavior.J.Chem.Ecol.,9(1):13-33.
Peschke K,1986.Development,sex specifity,and site of production of aphrodisiac pheromones in Aleochara curtula.J.Insect.Physiol.,32:687-693.
Peschke K,1987a.Cuticular hydrocarbons regulate mate recognition,male aggression,and female choice of the rove beetle,Aleochara curuda.J.Chem.Ecol.,13(10):1993-2008.
Peschke K,1987b.Male aggression,female mimicry and female choice in the rove beetle,Aleochara curtula(Coleoptera:Staphylinidae).Ethology,75:265-284.
Peschke K,1990.Chemical traits in sexual selection of the rove beetle,Aleochara curtula(Coleoptera: Staphylinidae).Entomol.Gener.,15:127-132.
Peschke K,Friedrich P,Gantert C,Metzler M,1996.The use of the tergal gland defensive secretion in male intrasexual aggression of the rove beetle,Aleochara curtula(Coleoptera:Staphylinidae),measured by closed-loop-stripping-analyses and tandem bioassay-mass fragmentography.Chemoecology,7:24-33.
Peschke K,Friedrich P,Kaiser U,Franke S,Francke W,1999.Isopropyl(Z9)-hexadecenoate as a male attractant pheromone from the sternal gland of the rove beetle Aleochara curtula(Coleoptera:Staphylinidae).Chemoecology,9:47-54.
Qi N,2009.A Taxonomic Study on the Genus Gabrius(Coleoptera:Staphylinidae:Staphylininae) of China.Shanghai: Shanghai Normal University Thesis for Master Degree.[齐楠,2009.中国佳隐翅虫属分类研究(鞘翅目:隐翅虫科:隐翅虫亚科).上海:上海师范大学硕士学位论文]
Quennedeya A,Drugmand D,Deligne J,2002.Morphology and ultrastructure of paired prototergal glands in the adult rove beetle Philonthus varians(Coleoptera,Staphylinidae).Arthropod.Struct.Dev.,31:173-183.
Schierling A,Dettner K,Schmidt J,Seifert K,2012.Biosynthesis of the defensive alkaloid cicindeloine in Stenus solutus beetle.Naturwissenschaften,99(8):665-669.
Schildknecht H,Berger D,Krauss D,Connert J,Gehlhaus J,Essenbreis H,1976.Defensive chemistry of Stenus comma(Coleoptera:Staphylinidae)LXI.J.Chem.Ecol.,2(1):1-11.
Schlechter-Helas J,Schmitt T,Peschke K,2011.A contact anti-aphrodisiac pheromone supplied by the spermatophore in the rove beetle Aleochara curtula:mode of transfer and evolutionary significance.Naturwissenschaften,98:855-862.
Schlechter-Helas J,Schmitt T,Peschke K,2012.Learning individual signatures:rove beetle males discriminate unreceptive females by cuticular hydrocarbon patterns.Anim.Behav.,84:369-376.
Shimoda T,Shinkaji N,Amano H,1997.Prey stage preference and feeding behaviour of Oligota kashmirica benefica(Coleoptera:Staphylinidae),an insect predator of the spider mite Tetranychus urticae(Acari:Tetranychidae).Exp.Appl.Acarol.,21:665-675.
Shower JA,Kistner DH,1977.The natural history of the myrmecophilus tribe Pygostenini(Coleoptera: Staphylinidae)section 4,glandular anatomy of the Pygostenini.Sociobiol,2:305-326.
Smith RL,1979a.Paternity assurance and altered roles in the mating behaviour of a giant water bug,Abedus herberti(Heteroptera:Belostomatidae).Anim.Behav.,27:716-725.
Smith RL,1979b.Repeated copulation and sperm precedence:paternity assurance for a male brooding water bug.Science,205:1029-1031.
Steidle JLM,Dettner K,1995.The chemistry of the abdominal gland secretion of six species of the rove beetle Genus Bledius.Biochem.Syst.Ecol.,23(7/8):757-765.
Stoeffler M,Maier TS,Tolasch T,Steidle JLM,2007.Foreign-language skills in rove beetles?evidence for chemical mimicry of Ant alarm pheromones in myrmecophilous Pella beetles(Coleoptera: Staphylinidae).J.Chem.Ecol.,33: 1382-1392.
Teng ZQ,Zhang QW,2006.Sperm competition and its avoidance mechanisms in insects.Journal of China Agricultural University,11(6):7-12.[滕兆乾,张青文,2006.昆虫精子竞争及其避免机制.中国农业大学学报,11(6):7-12]
Thayer MK,1987.Revision of the austral genus Metacorneolabium and studies in the systematics and biogeography of Omaliine Staphylinidae(Coleoptera).Syst.Ent.,12:389-289.
Waage JK,1979.Dual function of the damsel fly penis:sperm removal and sperm transfer.Science,203:916-918.
Waage JK,1982.Sperm displacement by male Lestes vigilax Hagen(Zygoptera:Lestidae).Odonatologica,11:201-209.
Waage JK,1984.Sperm competition and the evolution of Odonate mating systems.In:Smith RL ed,Sperm Competition and the Evolution of Animal Mating Systems.Academic Press,New York,251-290.
Wang HQ,Li JG,2011.Advances of Research the Sperm Competition of Insect.Anhui Agri.Sci.Bull.17(9):41-44.[王会琴,李金钢,2011.昆虫精子竞争的研究进展.安徽农学通报.17(9):41-44]
Wheeler JW,Happ GM,Araujo J,Pasteels JM,1972.γ-Dodecalactone from rove beetles.Tetrahedron Lett.,46:4635-4638.
Xu W,2009.A Taxonomic Study on the Genus Platydracus(Coleoptera:Staphylinidae:Staphylininae)of China.Shanghai:Shanghai Normal University Thesis for Master Degree.[许 旺,2009.中国普拉隐翅虫属分类研究(鞘翅目:隐翅虫科:隐翅虫亚科).上海:上海师范大学硕士学位论文]
Yokoi N,1990.The sperm removal behavior of the yellow spotted longicorn beetle Psacothea hilaris(Coleoptera:Cerambycidae).Appl.Entomol.Zool.,25:383-388.
Zhang Y,2004.A Taxonomic Study on the Genus Tachinus(Coleoptera:Staphylinidae:Tachyporinae)of China.Shanghai:Shanghai Normal University Thesis for Master Degree.[张 艳,2004.中国圆胸隐翅虫属分类研究(鞘翅目:隐翅虫科:隐翅虫亚科).上海:上海师范大学硕士学位论文]
Zheng FK,1992.Morphological taxonomy of the Staphylinidae.SiChuan:Sichuan University Press.[郑发科,1992.隐翅虫形态分类学纲要.四川:四川大学出版社]
Zheng YK,Zheng FK,Wang WL,Hou Y,Zheng H,Xu LJ,2009.Mating Behavior of the Stenus sp.(Coleoptera:Staphylinidae).Journal of China West Normal University(Natural Sciences),30(3):241-244.[郑英奎,郑发科,王伟连,侯勇,郑欢,徐莉娟,2009.大眼隐翅虫的交配行为.西华师范大学学报(自然科学版),30(3):241-244]
猜你喜欢
甘肃农业科技(2021年6期)2021-07-25 16:11:05
四川蚕业(2021年1期)2021-02-12 02:03:12
昆虫学报(2020年6期)2020-08-06 06:42:52
武夷科学(2019年2期)2019-12-20 08:39:16
科学导报(2019年35期)2019-09-03 04:35:11
西北农林科技大学学报(自然科学版)(2018年5期)2018-05-24 12:33:34
武夷科学(2017年0期)2017-03-10 10:03:31
百科知识(2016年13期)2016-08-05 18:23:30
中学生物学(2016年8期)2016-01-18 09:08:26
中国马铃薯(2015年3期)2015-12-19 08:03:56
