
日本佳盾蜾蠃营巢生物学研究
2013-11-22 08:01:18黄敦元余江帆黄名广朱朝东
环境昆虫学报 2013年6期
黄敦元,何 波,余江帆,谷 平,彭 飞,黄名广,朱朝东
(1.江西环境工程职业学院,江西赣州341000;2.南京师范大学生命科学学院动物学国家重点学科,江苏生物多样性重点实验室,南京210046;3.江西省林业科技培训中心,南昌330038;4.中国科学院动物研究所,动物进化与系统学 (院)重点实验室,北京100101)
胡蜂科Vespidae隶属膜翅目Hymenoptera细腰亚目 Clistogastra,全世界5000余种,262个属,6个 亚 科:Euparagiinae、 Masarinae、Eumeninae(蜾蠃亚科)、Stenogastrinae(狭腹胡蜂亚科)、Polistinae(马蜂亚科)和 Vespinae(胡蜂亚科)(Carpenter,1982;Pickett& Carpenter,2010)。我国已知56属约270种,其中包括38变种,分属马蜂亚科 Polistinae、胡蜂亚科 Vespinae、蜾蠃亚科Eumeninae及细腹胡蜂亚科 Stenogastrinae(李铁生,1982,1985;周鑫等,2011,2012,2013;You et al.,2013)。胡蜂科昆虫多为捕食性昆虫,具有食性广、捕食迅速、食量大等特点,被广泛应用于农、林业害虫的防治,是目前最好的防治体型较大害虫的天敌昆虫之一 (李铁生,1985)。
胡蜂科昆虫大多选择避风雨、遮光的场所筑巢,但是不同种类,筑巢时间、地点、筑巢材料、巢的形状,巢室数目也不同。如凹纹胡蜂Vespa velutina Smith、金环胡蜂Vespa mandarinia Smith在土洞内筑巢 (宁仲根,1996);约马蜂 Polistes johnamae Radoszkowski、小金箍胡蜂 Vespa tropica haematodes Bequaert(马万炎和侯伯鑫,1990)、墨胸胡蜂Vespa velutina nigrithorax Buysson(杨维来和李位三,1997)和黄腰胡蜂Vespa affinis Linnaeus(陈勇和孙希达,1996)一般选择在高大乔木上筑巢;密侧狭腹胡蜂Parischnogaater mellyi Sau.喜欢选择马路、河沟边缘的土坎、岩床下及茅草房和竹楼下筑巢 (董大志和大缘公雄,1994);黑盾胡蜂Vespa bicolor Fabricius(陈勇和童迅,2004)、陆马蜂Polistes rothneyi grahmi Vecht选择在屋檐下或是宅内筑巢;本研究中的日本佳盾蜾蠃偏爱野外枯枝、芦苇、竹节等中空物质中筑巢。
蜾蠃亚科Eumeninae是胡蜂科中最大的一个类群 (Carpenter,1982;Kim et al.,2005),目前研究发现,全世界共有3585种,我国现有记录161种 (李铁生,1982,1985;Yamane 1990;Kim and Yamane,2001;周鑫等,2011,2012,2013;You et al.,2013)。蜾蠃亚科多为独栖物种,无固定居住巢穴,一般交配后开始营巢以产卵,其巢穴有两种类型:一种是呈中空壶状,上部有细颈,开口于颈端部的泥质巢穴 (李铁生,1982);另一种巢是利用开口于竹管、芦苇管等中空材料,衔泥分隔成多个虫室,直接将卵产于竹管或者苇管内壁上,产卵后外出捕捉蛾类幼虫带回虫室中以供其卵孵化出的幼虫食用,最后用泥块密封虫室和巢穴口 (Cowan,1991;Fateryga,2013)。
日本佳盾蜾蠃 Euodynerus nipanicus(Schulthess)属膜翅目Hymenoptera胡蜂科Vespidae蜾蠃亚科 Eumeninae佳盾蜾蠃属Euodynerus Dalla Torre 1904(李铁生,1986),该蜂主要分布在国内江苏、浙江、江西和台湾等地,国外主要分布在日本和韩国 (Kim,2012),是一种重要的独栖性天敌资源昆虫,其捕食凶猛,飞翔迅速,对林区鳞翅目蛾类害虫的控制作用明显(李铁生,1985)。本研究利用人工巢管收集并观察该蜂,旨在进一步明确其巢穴结构和营巢习性,对丰富蜾蠃亚科部分属筑巢的相关资料以及对该蜂的保护利用具有一定的意义。
人工巢穴 (trap-nesting)技术是人为利用芦苇、纸筒、竹子等材料制成一端开口一段封闭的巢管,或是通过直接在木头上钻蛀不同直径、不同深度的巢孔来诱集野生蜂类资源及其寄生蜂(蝇)资源的一种方法 (Karl,1967;王凤鹤等,2001;Andreas and Tscharntke,2002;Tylianakis et al.,2004)。该技术广泛用于野外膜翅目及其寄生蜂 (蝇)标本的采集 (Malaise,1937),生物多样性分析 (Buschini et al.,2010),昆虫生物学特性的研究 (Andreas and Tscharntke,2002),传粉昆虫的放养 (朱涛,1991)及天敌昆虫的放养 (李铁生,1982)等领域。
1 材料与方法
1.1 研究地点
调查地点主要有江西省赣州市赣县沙地镇、江西省新余市渝水区水北镇、江西省宜春市袁州区巫家、江西省赣州市南康市谭东镇、江西省赣州市湖边镇共5个典型样地。样地具体信息见表1。
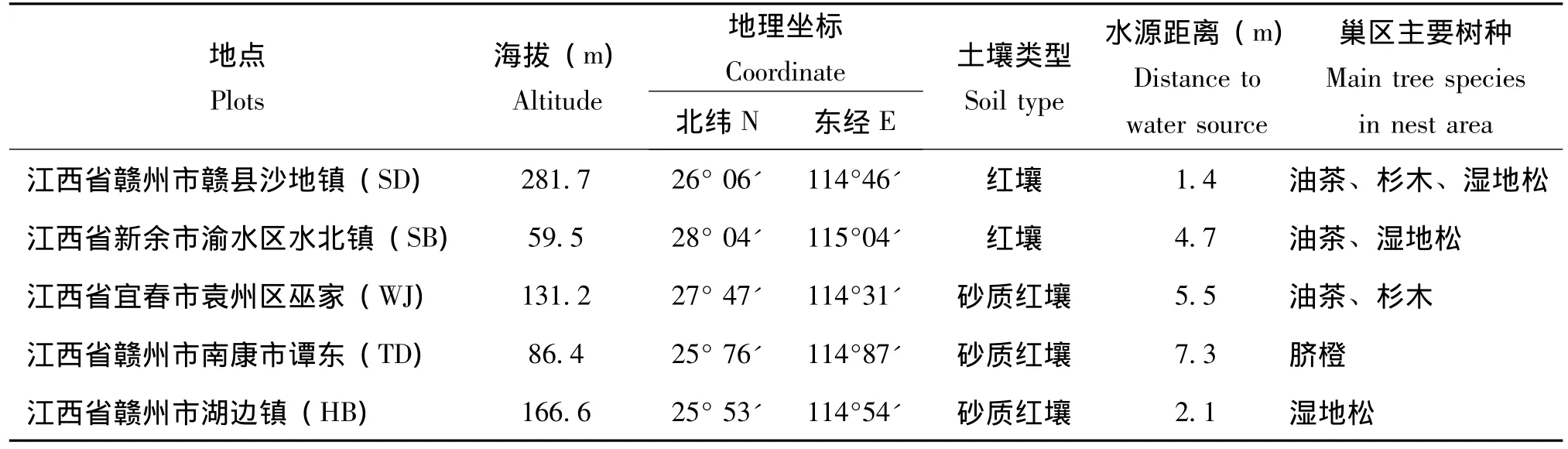
表1 不同样地的基本概况信息Table 1 The general information of different plots
1.2 人工巢穴的构建
人工巢管在野外的放置时间从2011年2月~2013年2月。为了能观察和收集日本佳盾蜾蠃Euodynerus nipanicus,在以上5个典型样地分别设置16个 PVC管材 (直径为 110 mm,长度为350 mn)制作的巢箱,分4处安置,每个支架上安装4个巢箱 (图版1,2),每个巢箱放置70~90个长度和粗细不等一段开口、一段封闭的芦苇管。为了研究该蜂对巢管高度、方向、长度、内径的不同偏好,我们的人工巢箱在同一个样地设置不同的高度和不同的巢口方向且巢箱中放置不同长度和不同粗细的巢管。根据前人的研究结果(Karl,1967;Tscharntke et al.,1998;黄敦元等,2012)和野外实验样地的具体林相,本实验巢箱巢口方向主要为南方、东方和东南方向为主,距离地面高度在1~2 m不等;单个巢管长度在12~30 cm之间,直径在0.6~1.2 cm之间。
1.3 雌性成蜂筑巢行为及蜜源植物观察
选择交配结束刚开始筑巢的雌性个体进行筑巢行为观察 (n=10),采取目测、摄像和巢管解剖相结合的方法进行连续观测,观察该蜂清理巢管杂物的过程及所用时间,巢穴附近500 m内跟踪该蜂 (包括雄性个体)观察并拍摄其到访的植物种类和访花行为。
1.4 雌性成蜂对巢管的选择及巢管结构观察
从9个巢箱中随机抽取36个筑巢巢管带回实验室进行解剖并测量记录虫室数目,虫室长度,空室数目,空室长度,前庭长度等数据,最后将解剖的巢管放入试管中 (管口用脱脂棉堵塞)进行室内饲养观察。
1.5 雌性成蜂日活动规律观察
2012、2013年4月中旬,在赣州市沙地镇样地对巢箱中有日本佳盾蜾蠃筑巢的巢管进行颜料标记,对该蜂越冬后第2代雌性个体全天出、入巢行为进行连续7 d的观察并进行记录,每天的观察时间从7∶00~18∶00,辅以拍照和摄像等方法。将采集的数据进行统计学分析。主要观察记录的数据有:雌性日本佳盾蜾蠃个体每天的活动规律,捕食过程所需要的时间,捕食回巢处理食物的时间,采泥的时间和携带泥土回巢的筑巢时间等。
1.6 数据统计分析
实验数据存储和图形制作采用SPSS 13.0和Excel 2003软件进行处理。
2 结果与分析
2.1 巢箱 (管)选择
研究表明:雌性日本佳盾蜾蠃更青睐巢口方向为东南方向的巢箱;在同一个支架上的4个巢箱中 (图版1),一般选择右下角的巢箱先筑巢,依次是左下角、右上角和左上角;巢管距离地面的高度一般在70~108 cm之间,均值87.53±8.74 cm(n=36)。将筑巢的巢管 (巢口以泥土密封)带回实验室并进行测量发现:该蜂筑巢选择巢管的长度在12~28 cm之间,平均21.03±3.75 cm(n=36),巢管的内径在0.622~1.198 cm之间,平均0.88±0.13 cm(n=36)。
2.2 巢室结构
雌性日本佳盾蜾蠃在自然界中一般在羽化巢管的附近巢管 (箱)筑巢,偶有利用老巢管的现象。该蜂巢穴主要包括虫室、空室和前庭三个部分 (图版10),巢穴中的预封塞物 (deep plug)和几个不同部分之间的隔膜都是雌性个体从巢穴附近衔回巢的湿润泥土构成。通过室内解剖并测量9巢箱36个巢管中的相关数据,可以发现该蜂巢穴中虫室 (brood cell)数目为1~10不等,平均4.75±1.83个 (n=36);空室 (deep cell)的数目一般比虫室的数目略多,一般在2~12之间,平均5.67±1.97个 (n=36);虫室长度在0.824~1.82 cm之间且个体之间的差异不大,平均1.32±0.26 cm(n=171);不同空室的长度在不同巢管中和同一个巢管不同的个体差异性明显,分别为0.254~1.302 cm不等,平均0.91±0.29 cm(n=204);一般每个巢管中都有前庭 (vestibular cell)且不同巢管中差异性较大,前庭的长度分布在0.49~7.02 cm之间,平均2.57±1.74 cm(n=36);每个虫室中的食物数目有一定的差异性,一般在5~15头之间,平均7.51±0.92头 (n=171)(见表2)。
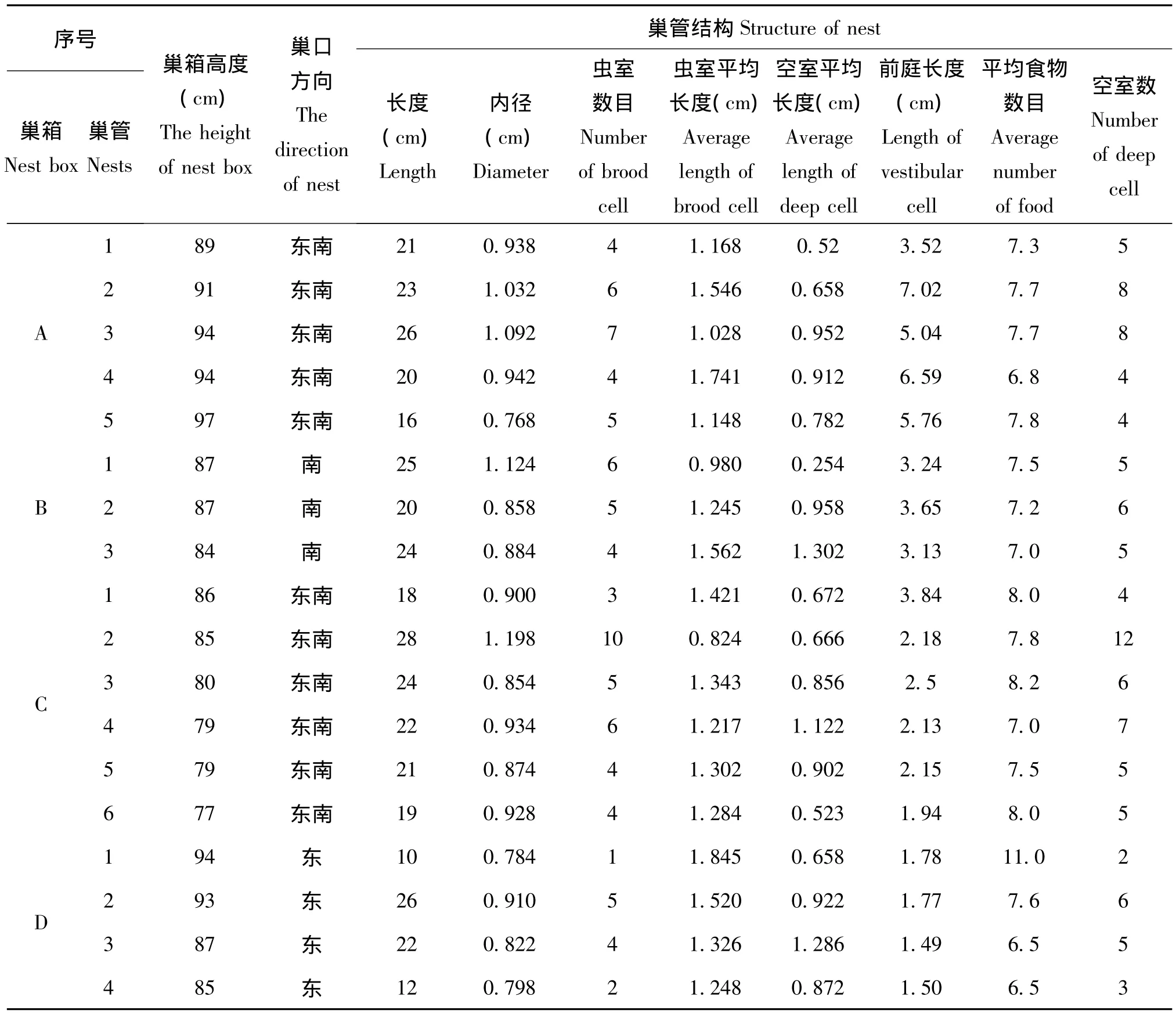
表2 日本佳盾蜾蠃巢管的相关特性Table 2 The related characteristics about nests of Euodynerus nipanicus
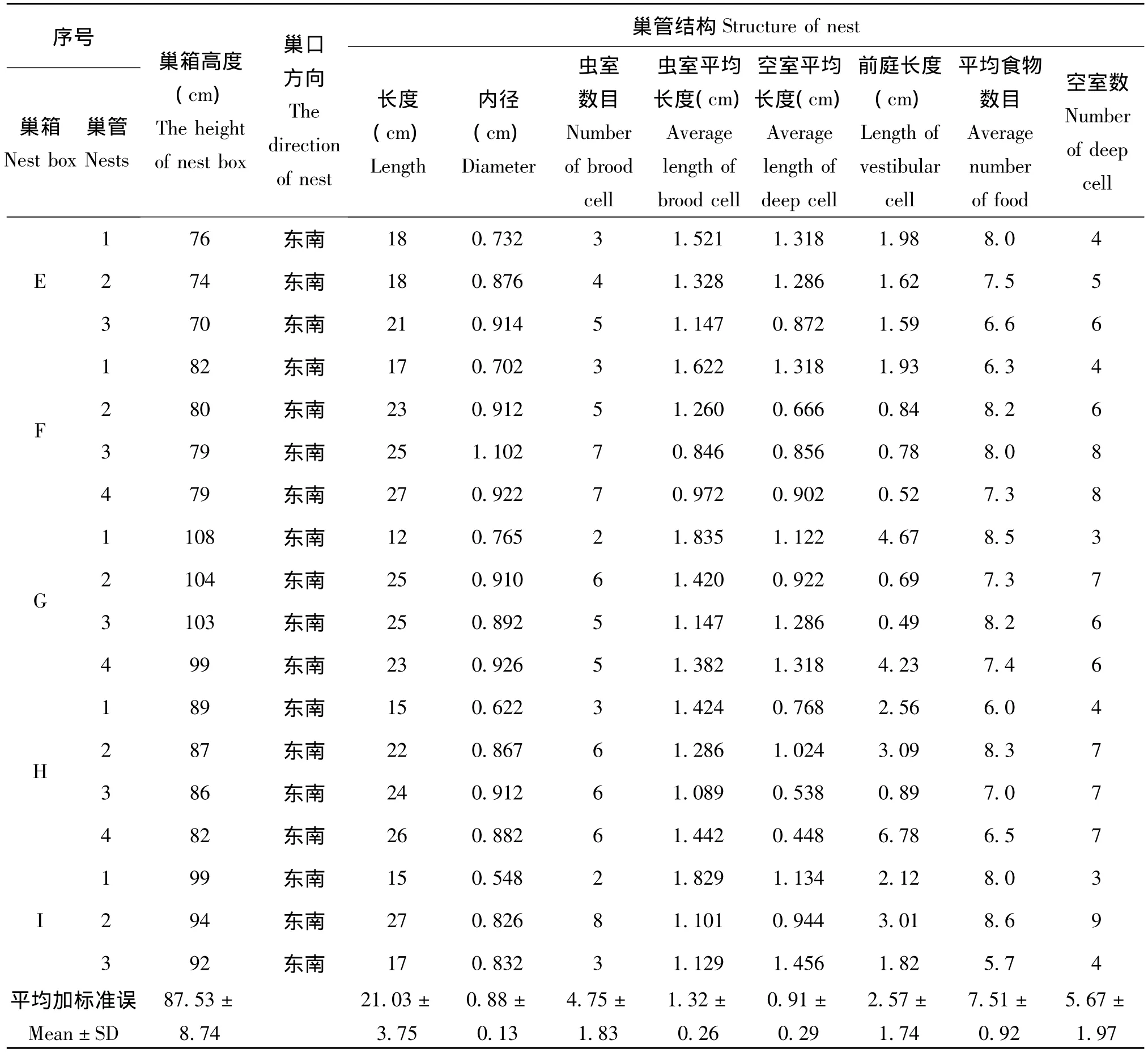
(续上表)
2.3 雌性成蜂筑巢行为观察
2.3.1 巢管杂物清理
雌性日本佳盾蜾蠃个体羽化出巢后可以很快完成交配并开始寻找适宜的筑巢环境和具体的巢管。由于新鲜芦苇管内存在大量絮状物质,不利于筑巢,所以筑巢之前需要对巢管内部进行清理,日本佳盾蜾蠃头部进入选定巢室,用上颚撕咬管内杂物使之碎片化,用上颚咬住和 (或)前足抱住杂物,调头 (巢管内径比较大)或直接退出巢口 (巢管内径比较小)飞向空中将其抛落,后进入巢穴继续清理,直至巢室通畅、整洁。每个巢管的清理时间在20.5±3.67 min(n=20)。
2.3.2 筑巢材料的采集
一般交配后的雌性日本佳盾蜾蠃清理好巢管后开始采集湿润的泥土筑巢 (图版4),先是采集泥块2~4次作为芦苇管的末端预封塞物 (deep plug),然后产卵,偶有巢管没有末端预封塞物。构建一个巢室需要采集泥块次数需要根据雌蜂选择的巢管内径及蜂室和空室隔离层的厚度存在的显著差异而定,一般为6~10次 (譬如内径8 mm的巢管构建5个虫室、4个空室和1个前庭需要来回衔泥75次)。日本佳盾蜾蠃泥土采集一般以就近原则为主,地面土壤湿润时一般在巢穴附近2 m之内采集,但巢穴附近地面土壤干燥时则会选择附近水源地采集湿润的泥土,另外,在夏季天气炎烈的气温条件下,该蜂也会从附近的水源取食一定的水来增加巢穴中先前衔回泥土的湿度,尤其是密封巢管时需要大量的潮湿泥土。采集泥土时,先用上颚倾斜于身体45°向内咬开一边 (图版4),再利用其前足的作用将泥块挤压成椭圆形(图版8)以便运回巢管中,泥块的直径约为3 mm,整个挖掘过程约13 s。
2.3.3 幼虫食物的采集
雌性日本佳盾蜾蠃在巢管虫室中产卵后立即出巢寻找幼虫的食物,一个虫室需要采集5~12只3~5龄的鳞翅目蛾类幼虫 (图版3,9)(根据食物的形态初步观察有5~7种不同的鳞翅目幼虫,计划通过进一步的野外观察和分子鉴定来确定具体物种及其对应的危害植物)。
2.4 蜜源植物
日本佳盾蜾蠃是一种重要的独栖性天敌资源昆虫,雄性个体主要完成交配,不参与筑巢和后代的抚育;雌性个体交配后便选择合适的地方和材料进行筑巢并采集鳞翅目幼虫带回虫室作为其后代的蜂粮,雌性个体在成虫期间也需要到访紫藤Wisteria sinensis Sweet、黄荆Vitex negundo Linn.、红蓼 Polygonum orientale Linn.、野棉花 Anemone hupehensis Lemoine.等植物的花朵取食花蜜补充自身的营养 (图版5)。
2.5 雌性成蜂日活动规律观察
在江西省赣州市等地,日本佳盾蜾蠃每年的活动时间为4月至11月中。晴好天气下,该蜂一般在6∶ 30~18∶ 30时间段出巢活动,其中10∶00~12∶00为该蜂雌性个体的活动频繁阶段,出巢活动主要是捕食鳞翅目蛾类幼虫作为抚育后代的蜂粮或是外出采集泥块构建巢管中的虫室。晴好天气下,雌性成蜂一天共出巢55±5.33次(n=30),其中采集花蜜的次数约7次,每次出巢前一般在巢口停留2~20 s不等,平均9.32±3.26 s(n=100)。该蜂出巢捕食成功率90%左右 (n=100),捕食过程所需时间 (从离开巢到衔食物回巢)在246~965 s之间,平均536.97±110.76 s(n=100)(图1 A),存放食物时间在19~71 s之间,平均40.15±8.46s(n=100)(图1 B);该蜂出巢采泥成功率95% 左右 (n=100),一次采泥过程 (从离开巢到衔泥块回巢)所需要的时间在15~83 s之间,平均39.07±11.15 s(n=100)(图1 C),筑巢时间在 20~119 s之间,平均68.56±13.59s(n=100)(图1 D)。
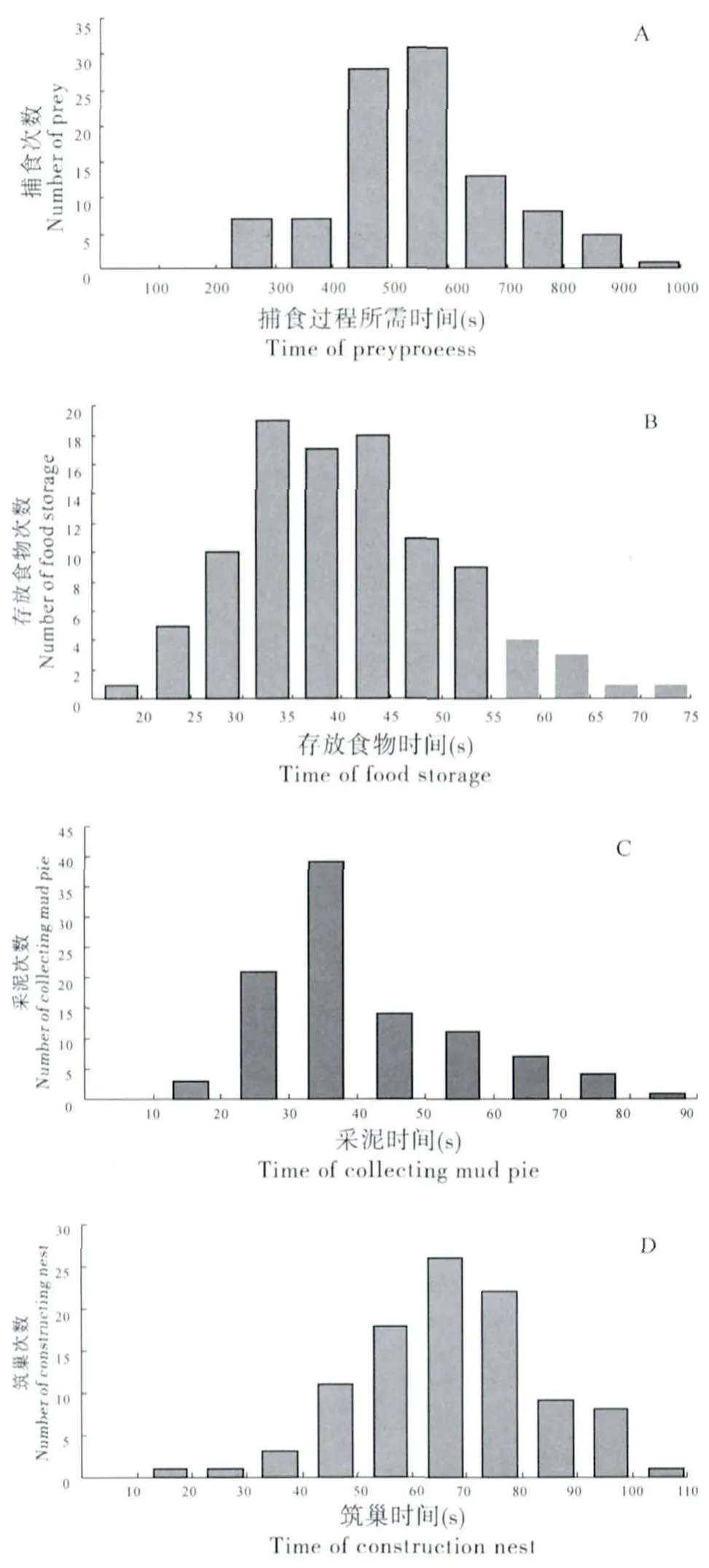
图1 日本佳盾蜾蠃捕食时间 (A)、存放食物时间 (B)、采泥时间 (C)和筑巢时间 (D)Fig.1 The time of prey process(A),food storage(B),collecting mud pie(C)and constructing nest(D)of Euodynerusnipanicus
3 结论与讨论
3.1 蜾蠃亚科的巢穴结构
蜾蠃亚科多为独栖物种,平时营自由生活,腹部第一节延长成柄状的蜾蠃类一般通过自己衔泥制成呈中空壶状的巢穴 (李铁生,1982);腹部没有延长柄状结构的蜾蠃类一般利用苇管、竹管等中空物质来构建自己的巢穴并直接产卵于竹管或者苇管内壁上抚育后代 (周鑫等,2012;Fateryga,2013)。日本佳盾蜾蠃是典型的利用苇管或者竹管筑巢的物种,其完整的巢穴结构包括末端预封塞物、虫室、空室、前庭及室间隔离层等5个部分。该蜂筑巢时从巢管的末端开始,一般按照构筑末端预封塞物、构建虫室、产卵、准备蜂粮、蜜蜂虫室、构建空室、下一个虫室、最后用泥块密封管口意味前庭构筑完成。同一个巢管中,虫室有大小两种,但是差异性不大,两个虫室之间一定有个空室,但是空室的大小差异性比较大;不同的巢管中,前庭一定存在,但是其长度差异性较大,末端预封塞物不一定存在。另外,巢管的内径大小对该蜂在巢管内的行为有一定影响,比较粗的巢管,雌蜂因在巢管内自由掉头而缩短其进出巢时间,但是因筑巢需要的泥块多而增加其生殖成本;比较细的巢管,有利于筑巢,但该蜂无法在巢管内完成掉头动作,只能缓慢退出巢管而增加进出巢时间,因此也会影响到其生殖成本。总之,日本佳盾蜾蠃不同的个体之间筑巢习性基本相近,但是在巢穴内部结构、虫室数目、虫室大小等方面存在一定的差异。由于条件的限制,我们无法观察到日本佳盾蜾蠃在巢管内部的具体行为,如该蜂如何将衔回巢的泥块筑巢以及如何产卵并固定于巢管内壁之上等,这些问题还需要进一步的观察研究。
3.2 雌雄个体差异及性比
日本佳盾蜾蠃雌性成虫个体要明显大于雄性,这种差异也体现在虫室大小、幼虫的发育、蛹的大小和虫室中食物的多少,这种各个性状在性别上的差异在其他蜾蠃亚科物种中也有过报道(Cooper,1953)。对于蜾蠃亚科雌雄个体的差异和性比的影响因素,诸多研究中主要有以下不同观点:虫室中的蜂粮多少是影响雌雄个体大小和性比的直接原因,如 Ancistrocerus antilope Panzer(Cooper,1953)和 Euodynerus foraminatus(David,1981)的巢穴中,雌性个体多来自储存食物量多的虫室,雄性个体发育于食物量小的虫室;Fateryga(2013)通过野外观察和解剖Euodynerus quadrifasciatus的巢管提出:巢管中单个虫室大小是影响不同虫室中卵和幼虫的发育情况进而影响到个体大小和雌雄性比,因为虫室的大小制约虫室中幼虫发育所需的食物数量;季节的更替也是导致雌雄个体差异及性比不同的因素之一 (Fye 1965;Longair 1981)。由于日本佳盾蜾蠃在观察样地一年4代,且有世代交替现象明显,因而增加了研究雌雄个体差异和性比的难度。
3.3 蜂粮的储存
我国是世界上将胡蜂类昆虫用于农林业生产上,消灭农林害虫的国家之一,这种以虫治虫的生物防治方法对降低化学农药污染有一定的意义(李铁生,1985),目前利用比较多的类群主要有蜾蠃类、胡蜂类、马蜂类等,其中蜾蠃类胡蜂对人类的危害相对较低。
雌性蜾蠃捕虫时,以口器咬住猎物颈部,再以蜇针蜇刺捕获幼虫的腹部神经,使其处于麻醉状态,然后抱回巢中储备为食物 (周鑫等,2012)。尽管蜾蠃的蜂毒具有特殊的麻痹和贮存活体寄主的功能 (许俊蜂和韩召军,2009),但是在日本佳盾蜾蠃从卵期到老熟幼虫期大约10天 (赣州地区2012年第2代卵期和幼虫期的数据)的时间如何实现鳞翅目蛾类幼虫的存活还需要进一步研究。
致谢:感谢重庆师范大学李廷景博士和福建农林大学刘长明博士对本文胡蜂标本进行了鉴定;中央财政林业科技推广项目 (JXTG(2012)11)提供了资助。同时感谢匿名审稿人和编辑的指正与帮助。

References)
Andreas K,Tscharntke T,2002.Grazing intensity and the diversity of grasshoppers,butterflies, and trap - nesting bees and wasps.Grazing Intensity and Insect Diversity,6:1570-1580.
Buschini MLT,Borba NA,Brescovit AD,2010.Prey selection in the trap - nesting wasp Trypoxylon(Trypargilum)opacum Brèthes(Hymenprtera:Crabronidae).Brazilian Journal of Biology,70:529-536.
Carpenter JM,1982. The phylogenetic relationships and natural classification of the Vespoidea(Hymenoptera).Systematic Entomology,7(1):11 -38.
Carpenter JM,1986.A synonymic generic checklist of the Eumeninae(Hymenoptera:Vespidae).Psyche,93(1/2):61-90.
Chen Y,Sun XD,1996.A primary study of the living habits and utilization of Vaspa affinis.Journal of Hangzhou Normal University,6:39-43.[陈勇,孙希达.1996.黄腰胡蜂生活习性及其利用的初步研究.杭州师范学院学报,6:39-43]
Chen Y,Tong X,2004.Observations on the bionomics of the haplometrosis of Vespa bicolor.Entomological Knowledge,41(3):263-264.[陈勇,童迅.2004.黑盾胡蜂单母建群习性的观察.昆虫知识,41(3):263-264]
Cooper KW,1953. Biology of eumenin wasps. I. The ecology,predation and competition of Ancistrocerus antilope(Panzer).Trans.American Entomol.Soc.,79:13 -35.
Cowan DP,1991.The solitary and presocial Vespidae.In:The Social Biology of Wasps. Ross KC,RW Matthews,eds. Ithaca and London:Comstock Publishing Associates.678.
David PC,1981.Parental investment in two solitary wasps Ancistrocerus adiabatus and Euodynerus foraminatus(Eumenidae:Hymenoptera).Behavioral Ecology and Sociobiology,9(2):95 -102.
Ding DZ,Kimio O,1994.Biology and distribution of Stenogastridae in southern China(Hymenoptera:Vespoidea).Acta Entomologica Sinica,37(4):450-457.[董大志,大缘公雄,1994.中国南部狭腹胡蜂的生物学与地理分布.昆虫学报,37(4):450-457]
Fateryga AV,2013.The nest structure in four wasp species of the genus Euodynerus(Hymenoptera,Vespidae:Eumeninae).Entomological Review,99(4):412-421.
Fye RE,1965.The biology of the Vespidae,Pompilidae and Sphecidae(Hymenoptera)from trap nests in northwestern Ontario.Canadian Entomol.,97:716-744.
Huang DY,Zhu CD,Xiao ZY,Huang MG,Hu DN,2012.Application of trap-nesting technique in the research of entomology.Journal of Anhui Agri.Sci.,40(10):974-975.[黄敦元,朱朝东,肖忠优,黄名广,胡冬南.2012.人工巢穴技术在昆虫学研究中的应用.安徽农业科学,40(10):974-975]
You J, Chen B, Li TJ,2013. Two new species of the genus Ancistrocerus Wesmael(Hymenoptera:Vespidae:Eumeninae)from China,with a key to the Oriental species.Zookeys,303:77 -86.
Karl VK,1967.Trap-Nesting Wasps and Bees.Life Histories,Nests,and Associates.Washington,D.C.:Smithsonian Press.576.
Kim JG,Choi YC,Choi JY,Kim SE,Kim KY,Kim JK,Lee JE,2005.Environmental evaluation by using Hymenoptera induced by bamboo pipe traps indicated by eumenid wasp(Hymenoptera:Eumenidae).Korean Journal of Applied Entomology,44:307-315.
Kim JK,2012.Taxonomic review of the genus Euodynerus(Hymenoptera:Vespidae:Eumeninae)in the Korean Peninsula.Anim.Syst.Evol.Divers.,28(3):161 -167.
Kim JK, Yamane S, 2001. A revision of Eumenes Latreille(Hymenoptera:Vespidae)from the Far East Asia,with descriptions of one new species and one new subspecies.Entomological Science,4(2):139 -155.
Lee TS, 1982. Wasps in Rural Region of China. Beijing :Agricultural Publishing House.64-115.[李铁生,1982.中国农区胡蜂.北京:农业出版社.64-115]
Lee TS, 1985. Economic Insect Fauna of China, Fasc. 30,Hymenoptera:Vespoidea.Beijing:Science Press.1-159.[李铁生,1985.中国经济昆虫志.第三十册.胡蜂总科.北京:科学出版社.1-159]
Longair RW,1981.Sex-ratio variations in xylophilous aculeate Hymenoptera.Evolution,35:597-600.
Ma WY,Hou BX,1990.Observe nest structure of Vespa tropica haematodes using nest anatomy.Entomological Knowledge,27(1):32-34.[马万炎,侯伯鑫,1990.小金箍胡蜂越冬期剖巢观察.昆虫知识,27(1):32-34]
Martins RP,Pimenta HR,1993.Ecologiae comportamento de vespas solitárias predadoras.Ciência Hoje,15:14 -19.
Ning ZG,1996.The living habits and control methods of Vespa mandarinia.Apicultural Science and Technology,45(4):155-162.[宁仲根,1996.金环胡蜂的生活习性及防治方法.养蜂科技,45(4):155-162]
Pickett KM and Carpenter JM,2010.Simultaneous analysis and the origins of sociality in the Vespidae(Insecta:Hymenoptera).Arthropod Systematics and Phylogeny,68(1):3 -33.
Tscharntke T,Gathmann A,Steffan-Dewenter I,1998.Bioindication using trap-nesting bees and wasps and their natural enemies:community structure and interactions.Journal of Applied Ecology,35:708-719.
Tylianakis J,Veddeler D,Lozada T,2004.Biodiversity of land-use systems in coastal ecuador and bioindication using trap-nesting bees,wasps,and their natural enemies.Lyonia,2:7 -15.
Wang FH,Guo ZH,Xu XL,Chen CG,2001.The main influence factors about retrieving of osmia and it's improvement mechanism.Beijing Forestry Science,4:29-30.[王凤鹤,郭志弘,徐希莲等,2001.壁蜂回收的主要影响因素及其改进方法.北京林业科学,4:29-30]
Xu JF,Han ZJ,2009.Studies on toxic characteristics of wasp venom from Rhynchium brunneum Fabricius(Hymenoptera:Eumenidae).Journal of Nanjing Agricultural University,32(4):89-93.[许俊蜂,韩召军,2009.黄唇蜾蠃蜂蜂毒的毒性特征研究.南京农业大学学报,32(4):89-93]
Yamane S, 1990. A revision of the Japanese Eumenidae(Hymenoptera,Vespoidea).Insecta Matsumurana,New Series,43:1-189.
Yang WL,Li WS,1997.The biological observations about Vespa velutina nigrithorax Buysson.Entomological Knowledge,34(1):29-30.[杨维来,李位三,1997.墨胸胡蜂生物学特性的观察.昆虫知识,34(1):29-30]
Zhou X,Chen B,Li TJ,2012.Two new species and a key to species of the genus Eumenes Latreille (Hymenoptera:Vespidae:Eumeninae)from southwestern China. Entomotaxonomia, 34(2):467-474.
Zhou X,Chen B,Li TJ,2013.Two new species of the genus Discoelius Latreille(Hymenoptera:Vespidae:Eumeninae)from China,with a key to the Chinese species.Journal of Hymenoptera Research,32:45-54.
Zhou X,Li TJ,Chen B,2011.The taxonomic research progress of Eumeninae(Hymenoptera:Vespidae).Journal of Chongqing Normal University(Natural Science),28(6):22-29.[周鑫,李廷景,陈斌,2011.蜾蠃亚科 (膜翅目:胡蜂科)分类研究进展.重庆师范大学学报 (自然科学版),28(6):22-29]
猜你喜欢
林业科学(2021年12期)2021-02-12 05:57:40
中国蜂业(2020年7期)2020-08-21 08:04:40
广西林业科学(2020年2期)2020-08-08 05:49:30
生物学通报(2020年8期)2020-06-21 06:51:58
武夷科学(2019年2期)2019-12-20 08:39:16
中国蜂业(2017年10期)2017-10-21 06:50:42
辽宁大学学报(自然科学版)(2017年1期)2017-04-06 01:39:46
武夷科学(2017年0期)2017-03-10 10:03:31
安徽农业科学(2016年18期)2016-09-24 01:54:29
小星星·阅读100分(高年级)(2009年9期)2009-10-19 09:07:42
