
棉红铃虫信息素结合蛋白的同源模建
2013-06-11 10:29:14赵雷修周凡蕊
环境昆虫学报 2013年6期
孙 浩,舒 茂,赵雷修,周凡蕊
(1.西南林业大学理学院,昆明 650224;2.重庆理工大学药学与生物工程学院,重庆 400054)
气味结合蛋白(Odorant Binding Proteins,OBPs)是在脊椎动物和昆虫体内发现的一类嗅觉蛋白。它们是细胞外的较小的可溶性蛋白,位于感觉树突周围的液体中。通常认为它可以捕捉气味和信息素分子,并把它们转运到嗅觉受体上(Vogt et al.,1985;Pelosi and Maida,1990)。昆虫对信息素和其他气味的感知是一系列复杂过程的结果,近年来的研究对这些过程进行了很多阐述。脊椎动物和非脊椎动物都通过它们的嗅觉系统来探测气味,动物的化学感觉对于觅食,交配,躲避有毒物质和恶劣环境来说是必要的(Li et al.,2008)。对于陆生动物,大多数气味都是空气中的疏水小分子,它们必须穿过脊椎动物的粘液、昆虫的感受器淋巴液等亲水介质才能到达嗅觉受体(Olfactory Receptors,ORs)(Jiang et al.,2009;Lagarde et al.2011)。这个克服亲水介质能垒的过程通常需要借助气味结合蛋白(Odorant Binding Proteins,OBPs)来完成(Zhang et al.,2012)。气味分子的转运是陆生动物独有的,就这一点而言,气味结合蛋白(OBPs)从很大程度上体现了陆生动物嗅觉系统的适应性进化(Vogt et al.,1991)。
在昆虫中OBPs 可以分成3种类型(Zhou et al.,2009;Spinelli et al.,2012),分别是:信息素结合蛋白(Pheromone-Binding Proteins,PBPs),一般气味结合蛋白(General Odorant-Binding Proteins,GOBPs)和触角特异性蛋白(Antennal Specific Proteins,ASPs)也称为触角结合蛋白x,(Antennal-Binding Proteinx,ABPx)。昆虫触角上的感器淋巴液(Sensillum Lymph)中含有大量的OBPs(浓度高达10 mM)(Vogt and Riddiford,1981;Klein,1987;Vogt et al.,1989)。
一般认为OBPs 的主要功能是识别和转运气味分子。随着研究的深入,人们又提出了OBPs 其他一些功能:(1)OBPs 可以作为清除剂,清除结合在受体上的信息素和其它外来信息素,以保持受体的活性(Kaissling,1986;Pelosi and Maida,1990);(2)在气味分子浓度过高时,OBPs 可以作为过滤器来降低其浓度,避免发生受体长期灵敏度降低的情况(Pelosi and Maida,1990;Zhong et al.,2012);(3)电生理学数据显示多音大蚕蛾Antheraea polyphemus 的ApolPBPs和Bomby mori 的BmorPBPs 有助于嗅觉受体的激活。这种激活依赖于受体、PBP和信息素的三元缔合物(Pophof,2004)。由此看来,OBPs 还会扮演受体活性激发者的角色,而不仅仅是被动的运输者(Tegoni et al.,2004);(4)来自昆虫不同部位的OBPs 也意味着它们可能有不同的功能。
三维结构在研究OBPs 的功能方面起着至关重要的作用。因为在其它一些实验中得出的看似正确的结论,后来却被晶体结构实验证明是错误的(Mohanty et al.,2004),所以只有掌握OBPs 的三维结构才可以更加清楚明确的解释或者推测OBPs的功能。目前在NCBI 数据库中的PBPs 的序列超过2000个,而PDB 数据库中OBPs 的晶体结构只有几十个,这无疑对OBPs 的研究是一种障碍。不过随着同源模建方法的提出,这一问题被在一定程度上解决了。同源模建可以比较准确的预测一个蛋白质的三维结构,在一致性较高的情况下更是如此。
棉红铃虫Pectinophora gossypiella 是一种世界性棉花害虫,也是我国长江流域锦花生产上的主要害虫之一。从三十年代起,国内外许多学者对锦红铃虫的生物学、生态学特性以及防治技术等方面进行了许多研究。
本文即是通过同源模建方法,预测棉红铃虫信息素结合蛋白的三维结构,并对其进行分析,为OBPs 的功能研究提供理论基础。
1 材料与方法
1.1 模版的选择
根据棉红铃虫 OBP 的一级序列(GI:6272642),用BLAST 程序在PDB 数据库中进行搜索,寻找与其序列一致性较高的X 射线衍射晶体结构。最终找到一系列评估值(Evalue)最趋近于零,且序列一致性较高的晶体结构。从中选出分辨率较高的三个晶体作为模版分子,它们分别为:2P71(分辨率为2.0,Evalue 值为9e-53,A 链一致性为68.94%)、1DQE(分辨率为1.8,Evalue值为3e-55,B 链一致性为69.34%)和2P70(分辨率为2.1,Evalue 值为9e-53,A 链一致性为68.94%)。这三个晶体中,都是家蚕的信息素结合蛋白晶体。
1.2 结构的模建
用Modeller 程序将棉红铃虫的一级序列与模版分子的序列进行比对(图1),然后根据比对结果进行同源模建,生成1000个可能的结构。在这些结构中选取打分最优的一个结构进行研究。目标结构的序列比模版结构的序列长,长出的部分是结构C-端和N-端的loop 区,长出的部分由于没有模版,所以它们的构象是随机的产生的,其意义不大。为了便于研究,将目标结构两端多出的残基删去,仅保留中间的24~155 号残基。
2 结果与分析
2.1 结构评价
为了确定模建结构的合理性,本文使用UCLA网站上的Procheck,Verify3D和ERRAT 三个程序和ProSa2003 对模建结构进行了评价。
2.1.1 Procheck 评价结果
Procheck 程序主要用于评价模型中残基与残基之间的立体化学性质,用Ramachandran 图(拉氏构象图)表示。在Ramachandran 图(图2)中,A、B、L 三个区域为合理区域;a,b,l,p 四个区域为较合理区;~a,~b,~l,~p 为大体可以接受的区域;其余区域为不合理区域。


甘氨酸和脯氨酸在Ramachandran 图中的允许存在的范围比较大,所以非甘氨酸、非脯氨酸残基处于合理范围内的比例成为判断结构合理性的主要依据。目标结构中共含有119个非甘氨酸、非脯氨酸残基,其中112个处于合理区域内,约占总数的94.1%;6个处于比较合理的区域内,约占总数的5.0%;1个处于大体可以接受的范围内,约占总数的0.8%;没有非甘氨酸、非脯氨酸残基处于不合理的区域内。由此可以看出,目标结构总体上是合理的。
2.1.2 Verify 3D 评价结果
Verify3D 用于分析三维结构(3D)与一级序列(1D)的兼容性。一个残基3D-1D 的平均得分大于0.2 时为合理。由图3 可以看出,目标结构所有残基3D-1D 的平均得分都大于0.2,所以目标结构所有残基都是合理的。
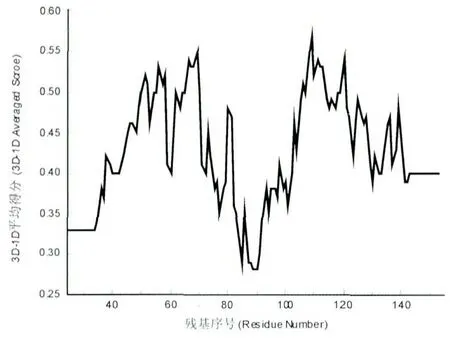
图3 Verify3D 打分图Fig.3 Verify3D scores model structure residues
2.1.3 ERRAT 评价结果
ERRAT 通过统计目标结构中不同原子间非键相互作用,并与高分辨率晶体结构的数据相比较,其误差值在置信区间内的区域被认为是合理的区域,合理区域所占的百分比越高说明结构质量越高。对于那些高分辨率晶体结构,这个百分比的平均值可以达到95%。由图4 可以看出本文目标结构的百分比为100%,说明目标结构的可靠性很高。
2.1.4 ProSa 2003 评价结果
ProSa2003 通过计算结构的能量和相应的标准分数(z-score)来判断其合理性。从计算得到的能量曲线(图5)上看,模建结构各个残基的能量均为负值,说明模建结构在能量方面是合理的。2P71、1DQE、2P70和模建结构的z-score 分别为-10.17、-9.77、-9.82和-10.23。模建结构的z-score 与模板结构的相近,这同样说明了模建结构的合理性。模建结构的z-score 小于模板结构的z-score 表明模建结构比模板结构更加稳定。

图4 ERRAT 计算的模建结构残基误差值图Fig.4 Error values of model structure residues

图5 ProSa2003 计算的模建结构各残基的能量图Fig.5 Energy of model structure residues
2.2 结构分析
从同源模建的结构(图6)中可以看出,棉红铃虫信息素结合蛋白由6个α-螺旋和连接这些螺旋的回折构成,其中5个α-螺旋(α1,α2,α4,α5,α6)构成了锥形的疏水结合口袋的侧壁,螺旋α3 处于锥形口袋的开口的顶部。螺旋α1和α2间,α3和α4 间,以及α5和α6 间的回折形成的边缘也位于锥形口袋的开口的顶部。锥形口袋的窄端是由螺旋α4和α5 之间的回折,以及螺旋α6末端和C-端的回折共同构成的。六个保守的半胱氨酸上的硫原子之间的距离为0.2 nm,处在二硫键键长的范围内,说明这六个半胱氨酸可以形成三个二硫键,从而对结构起到稳定的作用。

图6 棉红铃虫信息素结合蛋白同源模建结构Fig.6 Model structure of Pectinophora gossypiella
3 结论与讨论
本文用同源模建方法构建了棉红铃虫信息素结合蛋白的三维结构。从对结构的评价结果来看,该结构的具有很高的可靠性,在没有晶体结构的情况下,可以代替实验得到的晶体结构对棉红铃虫信息素结合蛋白的性质进行分析,也可以作为分子对接和动力学模拟的基础。从结构看,棉红铃虫信息素结合蛋白的主要结构是6个α-螺旋,其中五个构成锥形的疏水口袋,可以结合信息素分子。六个保守的半胱氨酸可以形成三个二硫键,从而对结构起到稳定的作用。这些信息为棉红铃虫信息素结合蛋白的深入研究打下了基础,为相关农药的研究提供有意义的借鉴,最终为农林虫害的防治提供帮助。
References)
Jiang QY,Wang WX,Zhang ZD,Zhang L,2009.Binding specificity of locust odorant binding protein and its key binding site for initial recognition of alcohols.Insect Biochemistry and Molecular Biology,39:440-447.
Kaissling KE,1986.Chemo-electrical transduction in insect olfactory receptors.Annu.Rev.Neurosci.,9:121-145.
Klein U,1987.Sensillum-lymph proteins from antennal olfactory hairs of the moth Antheraea polyphemus(Saturniidae).J.Insect Biochem.,17:1193-1204.
Lagarde A,Spinelli S,Tegoni M,He XL,Field L,Zhou JJ,Cambillau C,2011.The Crystal structure of odorant binding protein 7 from Anopheles gambiae exhibits an outstanding adaptability of Its binding site.J.Mol.Biol.,414:401-412.
Li S,Picimbon JF,Ji SD,Kan YC,Qiao CL,Zhou JJ,Pelosi PL,2008.Multiple functions of an odorant-binding protein in the mosquito Aedes aegypti.Biochemical and Biophysical Research Communications.,372:464-468.
Mohanty S,Zubkov S,Gronenborn AM,2004.The solution NMR structure of Antheraea polyphemus PBP provides new insight into pheromone recognition by pheromone-binding proteins.J.Mol.Biol.,337:443-451.
Pelosi P,Maida R,1990.Odorant binding proteins in vertebrates and insects:Similarities and possible common function.Chem.Senses,15:205-215.
Pophof B,2004.Pheromone-binding proteins contribute to the activation of olfactory receptor neurons in the silkmoths Antheraea polyphemus and Bombyx mori.Chem.Senses,29:117-125.
Spinelli S,Lagarde A,Iovinella I,Legrand P,Mariella T,Pelosi P,Cambillau C,2012.Crystal structure of Apis mellifera OBP14,a Cminus odorant-binding protein,and its complexes with odorant molecules.Insect Biochemistry and Molecular Biology,42:41-50.
Tegoni M,Campanacci V,Cambillau C,2004.Structural aspects of sexual attraction and chemical communication in insects.Trends Biochem.Sci.,29:257-264.
Vogt RG,Riddiford LM,1981.Pheromone binding and inactivation by moth antennae.Nature,293:161-163.
Vogt RG,Riddiford LM,Prestwich GD,1985.Kinetic properties of a sex pheromone-degrading enzyme:The sensillar esterase of Antheraea polyphemus.Proc.Natl.Acad.Sci.,82: 8827-8831.
Vogt RG,Rybczynski R,Lerner MR,1991.Molecular cloning and sequencing of general odorant-binding proteins GOBP1 and GOBP2 from the tobacco hawk moth Manduca sexta:Comparisons with other insect OBPs and their signal peptides.J.Neurosci.,11:2972-2984.
Vogt RG,Koehne AC,Dubnau JT,Prestwich GD,1989.Expression of pheromone binding proteins during antennal development in the gypsy moth Lymantria dispar.J.Neurosci.,9:3332-3346.
Zhang TT,Wang WX,Gu Shao H,Zhang ZD Wu KM,Zhang YJ,Guo YY,2012.Structure,binding characteristics,and 3D model prediction of a newly identified odorant-binding protein from the cotton bollworm,Helicoverpa armigera.Journal of Integrative Agriculture,11(3):430-438.
Zhou JJ,Robertson G,He XiaoL,Dufour S,Hooper A,Pickett J,Keep N,Field L,2009.Characterisation of Bombyx mori odorant-binding proteins reveals that a general odorant-binding protein discriminates between sex pheromone components.J.Mol.Biol.,389:529-545.
Zhong T,Yin J,Deng SS,Li K,Cao YZ,2012.Fluorescence competition assay for the assessment of green leaf volatiles and trans-b-farnesene bound to three odorant-binding proteins in the wheat aphid Sitobion avenae(Fabricius).Journal of Insect Physiology,58:771-781.
猜你喜欢
——紫 苏
河南农业(2024年1期)2024-01-19 01:56:54
华人时刊(2023年1期)2023-03-14 06:43:36
生物化学与生物物理进展(2022年6期)2022-07-21 11:52:06
汉字汉语研究(2021年2期)2021-08-30 08:58:46
中学生数理化(高中版.高考理化)(2021年2期)2021-03-19 08:52:38
中学生数理化(高中版.高考理化)(2020年2期)2020-04-21 05:33:04
衡阳师范学院学报(2016年3期)2016-07-10 07:16:27
河北书画研究(2016年3期)2016-04-28 08:55:35
池州学院学报(2015年3期)2016-01-05 01:13:04
天津科技大学学报(2015年2期)2015-08-09 01:40:42
