
Enhanced tolerance to drought in transgenic rice plants overexpressing C4photosynthesis enzymes
2013-05-08 08:24:41JunFeiGuMingQiuJianChangYang
The Crop Journal 2013年2期
Jun-Fei Gu,Ming Qiu,Jian-Chang Yang*
Key Laboratory of Crop Genetics and Physiology of Jiangsu Province,College of Agriculture,Yangzhou University,Yangzhou,Jiangsu,China
Enhanced tolerance to drought in transgenic rice plants overexpressing C4photosynthesis enzymes
Jun-Fei Gu,Ming Qiu,Jian-Chang Yang*
Key Laboratory of Crop Genetics and Physiology of Jiangsu Province,College of Agriculture,Yangzhou University,Yangzhou,Jiangsu,China
A R T I C L E I N F O
Article history:
Received 14 July 2013
Received in revised form
7 September 2013
Accepted 12 October 2013
Available online 24 October 2013
Pyruvate orthophosphate dikinase (PPDK)
C4-specific phosphoenolpyruvate
carboxylase(PEPC)and PPDK(PCK)
Transgenic rice
Photosynthesis
Drought tolerance
Maize-specific pyruvate orthophosphate dikinase(PPDK)was overexpressed in rice independently or in combination with the maize C4-specific phosphoenolpyruvate carboxylase(PCK).The wild-type(WT)cultivar Kitaake and transgenic plants were evaluated in independent field and tank experiments.Three soil moisture treatments, well-watered(WW),moderate drought(MD)and severe drought(SD),were imposed from 9 d post-anthesis till maturity.Leaf physiological and biochemical traits,root activities, biomass,grain yield,and yield components in the untransformed WT and two transgenic rice lines(PPDK and PCK)were systematically studied.Compared with the WT,both transgenic rice lines showed increased leaf photosynthetic rate:by 20%–40%under WW,by 45%–60%under MD,and by 80%–120%under SD.The transgenic plants produced 16.1%, 20.2%and 20.0%higher grain yields than WT under the WW,MD and SD treatments, respectively.Under the same soil moisture treatments,activities of phosphoenolpyruvate carboxylase(PEPC)and carbonic anhydrase(CA)in transgenic plants were 3–5-fold higher than those in WT plants.Compared with ribulose-1,5-bisphosphate carboxylase,activities of PEPC and CA were less reduced under both MD and SD treatments.The transgenic plants also showed higher leaf water content,stomatal conductance,transpiration efficiency,and root oxidation activity and a stronger active oxygen scavenging system than the WT under all soil moisture treatments,especially MD and SD.The results suggest that drought tolerance is greatly enhanced in transgenic rice plants overexpressing C4photosynthesis enzymes.This study was performed under natural conditions and normal planting density to evaluate yield advantages on a field basis.It may open a new avenue to droughttolerance breeding via overexpression of C4enzymes in rice.
©2013 Crop Science Society of China and Institute of Crop Science,CAAS.Production and hosting by Elsevier B.V.All rights reserved.
1.Introduction
Many important crops including rice(Oryza sativa L.),wheat (Triticum aestivum L.),soybean(Glycine max L.),and potato (Solanum tuberosum L.)are classified as C3plants,in which the first product of the Calvin cycle is 3-phosphoglycerate(3-PGA), whose production is catalyzed by ribulose 1,5-bisphosphate carboxylase/oxygenase(Rubisco).However,competition of O2with CO2at the catalytic site of Rubisco results in a loss of up to 50%of carbon fixation via photorespiration[1].Compared with C3plants,C4crops such as maize(Zea mays L.)and sorghum[Sorghum bicolor(L.)Moench]have evolved a C4-metabolism system that concentrates CO2in the vicinity of Rubisco and thereby substantially increases the ratio of RuBP carboxylation to oxygenation.This strategy suppresses photorespiration by more than 80%[2]and is accompanied by higher photosynthetic rates,yields,and increased water-and nitrogen-use efficiency,especially under low CO2concentration, high temperature,high light intensity,drought,and other stresses[3–6].Since the discovery of C4photosynthesis and its agronomic advantages,the genetic transformation of C3photosynthesis pathway into a C4systemhas become highly desirable.
The C4pathway in a C4crop such as maize(NADP malic enzyme(NADP-ME)C4cycle[7])consists of three key steps: (i)initial fixation of CO2by phosphoenolpyruvate carboxylase (PEPC)to form a C4acid;(ii)decarboxylation of C4acid to release CO2near the site of the Calvin cycle in bundle sheath cells by NADP-ME;and(iii)regeneration of the primary CO2acceptor phosphoenolpyruvate(PEP)by pyruvate orthophosphate dikinase(PPDK)[8].The transfer of C4key enzymes from C4plants to C3plants could contribute to introducing a C4system into C3plants,improving the rates of photosynthesis (Pn)and increasing crop yields[4,9].By use of an Agrobacteriumbased transformation system,genes that encode key C4enzymes such as PEPC,PPDK and NADP-ME have been successfully introduced and expressed in rice plants[9–14]. The transgenic rice plants have shown higher photosynthesis rates and often higher grain yield[4,10,15],although opposite results have also been reported[9,12,16,17].In addition, enzymes involved in C4photosynthesis play important roles in plant defense responses to biotic and abiotic stresses [4,15,18–20].However,the photosynthetic characteristics and grain yield of transgenic rice,especially under drought environments,have not been systematically examined.Few studies have been conducted under naturalfield conditions and normal planting densities to determine whether overexpressing C4photosynthesis in rice can result in a real improvement yield in terms of grain yield on a field basis[21].
Here we describe the photosynthetic characteristics and drought tolerance of transgenic rice overexpressing the maize C4PPDK enzyme independently or in combination with maize PEPC enzymes(PEPC+PPDK,PCK).By applying different levels of water stress during grain filling,we aimed to provide experimental evidence leading to an understanding of the mechanismunderlying the enhanced photosynthesis and grain yield in these transgenic plants under drought environments.
2.Materials and methods
2.1.Plant materials and cultivation
Two independent experiments(field and cement tank experiments)were conducted at a research farm of Yangzhou University,Jiangsu Province,China(32°30′N,119°30′E).The soil used in the experiments was a sandy loam(Typic Fluvaquent,Etisol)with 24.5 g kg−1organic matter,106 mg kg−1alkali-hydrolyzable N,33.8 mg kg−1Olsen-P,and 66.4 mg kg−1exchangeable K.An untransformed wild type(WT,Oryza sativa L.ssp.japonica cv.Kitaake)and two transgenic rice genotypes, homozygous transgenic rice overexpressing the maize PPDK, PEPC+PPDK(PCK and provided by Prof.MSB Ku,School of BiologicalSciences,Washington State University),were included in the study.Seedlings were raised in a seedbed and 20-day-old seedlings were then transplanted into both paddy field and cement tanks.
2.2.Soil moisture treatments
Both field and tank experiments were conducted.The field experiment was a three by three(three rice genotypes and three levels of soil moisture)factorial design with nine treatments,each with three replicates.Plot size was 4×3 m and plots were separated by an alley of 40 cm wide with plastic film inserted into the soil to a depth of 50 cm to form a barrier.Seedlings were transplanted at a hill spacing of 0.20×0.15 m with two seedlings per hill.N(60 kg ha−1as urea),P(30 kg ha−1as single superphosphate),and K(40 kg ha−1as KCl)were applied and incorporated justbefore transplanting. N as urea was also applied at mid-tillering(40 kg ha−1)and at panicle initiation(25 kg ha−1).All the genotypes headed on 13–15 July(50%of plants)and were harvested on 25 August. The water level in the field was kept at 1–2 cm until 9 days post-anthesis(DPA),when water stress treatments were initiated.From 9 DPAuntilmaturity,three treatments including well-watered(WW),moderate drought(MD),and severe drought(SD)were applied.The WW regime was flooded with 1–2 cm water depth.Soil water potential was maintained at−25±5 kilopascals(kPa)for the MD treatment and at−50± 5 kPa for the SD treatment.In each plot,four tensiometers (Institute of Soil Science,Chinese Academy of Sciences, Nanjing,China)consisting of a sensor of 5 cm length were installed to monitor soil water potential at 15–20 cm depth. Tensiometer readings were recorded every 4 h from 6:00 to 18:00.When the readings reached the desired values,tap water was added to the plot to maintain the values.
In the cement tank experiment,plants were grown in nine cement tanks in open-field conditions.Each tank(0.3 m height,1.5 m wide,and 9 m length)was filled with sandy loam soil with the same nutrient contents as in the field experiment.Twenty-day-old seedlings raised in the field were transplanted into the tanks at a hill spacing of 0.15×0.20 m with two seedling per hill.N(8 g m−2as urea),P(4 g m−2as single superphosphate),and K(5 g m−2as KCl)were applied and incorporated before transplanting.N as urea was also applied at mid-tillering(5 g m−2)and at panicle initiation (3 g m−2).The three treatments of WW,MD and SD were imposed from 9 DPA till maturity.The treatment details were the same as in the field experiment.Plot size was 3.0×1.5 m and each treatment had three replicates.A rain shelter consisting of a steel frame covered with plastic sheeting was used to minimize the effect of rainfall precipitation on the treatments,and was removed after rain.
2.3.Sampling and measurements
Six flag leaves from each treatment were sampled at 14 and 28 DPA for measurement of leaf water content.Leaf watercontent as a percentage of fresh mass was calculated according to the following equation:leaf water content(%)= 100×(FM−DM)/FM,where DM and FM denote respectively dry matter and fresh matter of the flag leaves.
Photosynthesis,chlorophyll and nitrogen content,and activities of PEPC,Rubisco and carbonic anhydrase(CA)in flag leaves were measured at 14 DPA and 21 DPA.Photosynthesis was determined under the conditions of 28°C,ambient CO2concentration,and 65%–70%relative humidity using a LI-6400 portable photosynthesis system(LI-COR,Lincoln, Nebraska,USA).The photosynthetic photon flux density(PPFD) was controlled by a LED light source built into the portable photosynthesis system and was set to 1500 μmol m−2s−1.Six leaves were measured for each treatment on each measurement date.Chlorophyll was extracted by shaking in methanol overnight and measured as described by Holden[22].Leaf nitrogen content was determined by micro Kjeldahl digestion, distillation,and titration[23].Activities of PEPC,Rubisco and CA were assayed according to the methods of Gonzalez et al.[24], Weiet al.[25]and Guo et al.[26],respectively.At 10,17,24 and 31 DPA,superoxide dismutase(SOD)activity and malondialdehyde (MDA)content of the flag leaves were determined according to the methods described by Giannopolitis and Ries[27]and by Zhao et al.[28],respectively.
Root exudates and root oxidation activity(ROA)were determined at 14 and 28 DPA.Six hills of plants from each treatment were used for collection of root exudates.Each plant was cut at an internode about 12 cm above the soil surface at 18:00 h.An absorbent cotton ball was placed on the top of each decapitated stem and covered with a polyethylene sheet.The cotton ball with exudates was collected after 6 h. The volume of exudates was estimated from the increase in cotton weight with the assumption that the specific gravity of the exudation sap was 1.0.For ROA measurement,a cube of soil (20×20×20 cm)around each individual hill was removed using a soil sampling corer.Plants of three hills from each plot formed a sample at each measurement.The roots of each hill were carefully rinsed with a hydropneumatic elutriation device (Gillison's Variety Fabrications,Benzonia,MI,USA).The equipment employs a high-kinetic-energy first stage in which water jets erode the soil from the roots followed by a second low-kinetic-energy flotation stage that deposits the roots on a submerged sieve[29].All the roots were detached manually from their nodal bases.A portion(10 g)of each root sample was used for measurement of ROA.The remaining roots were dried in an oven at 70°C for 72 h and weighed.The method for measurement of ROA was according to Yang et al.[30].Root activity was expressed asμgα-alpha-naphthylamine(α-NA)per gram dry weight(DW)per hour(μg α-NA g−1DW h−1).
Shoot biomasses were determined at 9 DPA and maturity. Plants from five hills were sampled from each plot at each measurement time.Measurement of grain yield and yield components at maturity followed Yoshida et al.[31].Plants in the two rows on each side of the plot were discarded to avoid border effects.In each plot,grain yield was determined from a harvest area of 5.0 m2in the field experiment and 2.0 m2in the tank experiment and adjusted to 14%moisture.Yield components(number of panicles per square meter,number of spikelets per panicle,percentage of filled grains,and grain weight)were determined from plants of 10 hills(excluding border plants)sampled randomly from each plot.The percentage of filled grains was defined as the number of filled grains(of specific gravity≥1.06 g cm−3)as a percentage of the total number of spikelets.
2.4.Statistical analysis
Analysis of variance was performed using the SAS/STAT statistical analysis package(version 6.12,SAS Institute,Cary, NC,USA).Data from each sampling date were analyzed separately.Means were tested by least significant difference at P=0.05(LSD0.05).
In this experiment,transgenic rice plants overexpressing maize PEPC,the rice NADP-ME,were also studied,and results from these plants were very similar to those of PPDK and PEPC+PPDK(PCK).For brevity only the results of WT and transgenic plants PPDK and PCK are reported here.
3.Results
3.1.Leaf physiological and biochemical characteristics
Fig.1 illustrates the progression of leaf water content after the water treatments.Average leaf water content fell from 76.0% at 14 DPA to 68.2%at 28 DPA.Transgenic plants(PPDK and PCK)consistently showed higher leaf water content than WT under different soil moisture treatments at DPA of 14 and 28. As water stress increased,transgenic plants showed greater ability to preserve higher leaf water content than WT plants, especially at 14 DPA.Average leaf water contents of transgenic plants at 14 DPA under the WW,MD and SD treatments were respectively 3.4%,3.5%and 4.7%higher than those of WT plants(Fig.1).
Daily changes in photosynthetic rate were evaluated in the tank experiment(Table 1).All the genotypes showed the same pattern of circadian rhythm of photosynthesis.Transgenic plants(PPDK and PCK)consistently showed higher Pnthan the WT during the day(P<0.05)under all three treatments,and no significant difference(P>0.05)was observed between the two transgenic lines(Table 1).On average,Pnlevels under the MD and SD treatments decreased by respectively 41.9%and 59.3%in WT plants,14.8%and 33.5%in PPDK,and 18.5%and 35.1%in PCK,relative to Pnunder the WW treatment, indicating that the transgenic plants had greater drought tolerance than WT plants in photosynthesis.
During the soil moisture treatments,photosynthesis was also measured at 14 DPA and 21 DPA in the field experiment (Table 2).The transgenic plants(PPDK and PCK)had higher Pn, gsand TE than the WT plants.When drought became severe, Pnand stomatal conductance(gs)of all the genotypes decreased,but transpiration efficiency(TE)increased,with greater increases in the transgenic than in the WT plants. Compared with WW treatments,for example,Pndecreased under the MD and SD treatments by respectively 25.8%and 56.4%for WT plants,but by only 13.1%and 35.7%for PPDK and by 13.8%and 34.9%for PCK.
Similar to leaf photosynthesis,chlorophyll and nitrogen contents in leaves and activities of Rubisco,PEPC and CA decreased under the soil-drying treatments,with greaterreduction in WT plants than in transgenic plants(Table 3). Under the same soil moisture levels,activities of PEPC and CA in transgenic plants were 3–5-fold higher than those in WT plants.Compared with the activity of Rubisco,activities of PEPC and CA were less reduced under both MD and SD, especially under MD(Table 3),suggesting that the enzymes involved in C4photosynthesis are more resistant to drought than that involved in C3photosynthesis.
The effect of drought on the antioxidative system in the form of MDA content and SOD activity was investigated (Fig.2).MDA content increased with increasing drought level and leaf age(Fig.2A–C),suggesting the production of excessive reactive oxygen species(ROS)caused by drought and leaf aging. Compared with WT rice,transgenic rice showed a significant lower content of MDA under all the soil moisture treatments (P<0.05),suggesting an improved tolerance of ROS damage especially under drought environments.In contrast to MDA content,SOD activities were higher for transgenic plants, especially for PPDK,than for WT plants under the MD and SD treatments(P<0.05,Fig.2D–F).
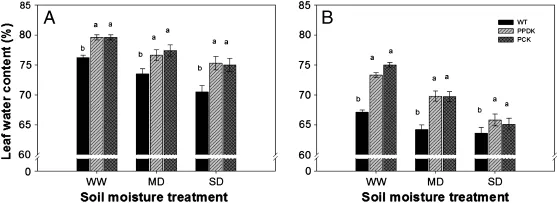
Fig.1–Leaf water content in wild-type(WT,cv.Kitaake)and two transgenic rice plants overexpressing pyruvate orthophosphate dikinase(PPDK);phosphoenolpyruvate carboxylase(PEPC),and PPDK(PCK),at different water treatments (well-watered,WW;moderate drought,MD;severe drought,SD),at(A)14 days post-anthesis(DPA)and(B)28 DPA in a tank experiment.Data are means of six plants per treatment±standard error.Letters a–c above bars indicate significant differences at P=0.05 among the three rice genotypes within the same soil moisture treatment.
3.2.Root exudates and root oxidation activity
Under the same soil moisture and on the same measurement dates,the volumes of root exudates of transgenic plants were much greater than those of WT plants(Fig.3).For the same genotype,the volume of root exudates decreased with increased soil drought,with a greater reduction for WT than for transgenic plants.In comparison with the WW treatment, under the MD and SD treatments the volumes of root exudates at 14 DPA decreased by respectively 27.0%and 66.1%for the WT,by 21.5%and 56.9%for PPDK,and by 6.3% and 50.7%for PCK and the volume of root exudates at 28 DPA decreased by respectively 42.1%and 71.4%for the WT,by 33.7%and 66.3%for PPDK,and by 20.7%and 63.6%for PCK.
The transgenic plants also showed higher root oxidation activity(ROA)than WT plants,especially under the drought treatments(Fig.4).For example,the ROA under the MD and SD treatments decreased by respectively 16%and 75%for WT plants,by 9.5%and 62.0%for PPDK plants,and by 12%and 65% for PCK plants,compared with that under the WW treatment.

Table 1–Daily patterns of photosynthetic rate(Pn)of wild-type rice(WT,cv.Kitaake),PPDK transgenic rice(PPDK),and PEPC+PPDK transgenic rice(PCK)in a tank experiment at 15 d post anthesis.

Table 2–Photosynthetic rate(Pn,μmol CO2m−2s−1),stomatal conductance(gs,mol CO2m−2s−1),and transpiration efficiency(TE,μmol CO2/mmol H2O)at 14 and 21 days post-anthesis(DPA)in the field experiment.
3.3.Biomass and grain yield
Although water stress significantly reduced biomass production for all the genotypes,transgenic genotypes consistently showed higher biomass at maturity than WT under all the soil moisture treatments(Table 4).For example,in the field experiment the transgenic plants had respectively 19.0%, 24.5%and 23.5%more biomass than WT under the WW,MD and SD treatments.
The transgenic plants showed higher grain yields than WT plants in both field and tank experiments and under the soil moisture treatments(Fig.5).On average in the field experiment,transgenic plants had 15.5%,22.4%and 21.0%higher grain yields than WT under WW,MD and SD treatments, respectively.In the tank experiment,transgenic plants had 16.7%,18.0%and 19.0%higher grain yields than WT under WW, MD and SD treatments,respectively,indicating an enhanced tolerance to drought in transgenic rice plants.
Because the soil drought treatments were imposed beginning at 9 DPA,panicle number per area and spikelet number per panicle were not affected by the treatments(Table 5).In comparison with the WW treatments,the percentages of filled grains and grain weight decreased under both MD and SD treatments,with greater decreases under the SDthan under the MD treatment.Under the same soil moisture,especially under the MDand SD treatments,both PPDK and PCK plants showed a greater percentage of filled grains than WTplants.Grain weight and harvest index varied with genotype and soil moisture treatment.Generally,the PCK plants exhibited higher grain weight and harvest index than WT plants under both MD and SD treatments(Table 5).
4.Discussion
Photosynthesis is fundamental to biomass production,but sensitive to drought.Improving photosynthesis-related physiological traits is thought to be a useful approach to increase yield and drought tolerance[10,32–34].Researchers worldwide have attempted to improve photosynthesis and crop yield by introducing C4cycle in plants by transgenic approaches [4,11,12,15,35,36].But there is a longstanding controversy as to whether an increase in leaf-level photosynthesis would increase yield[37–41].In the present study,transgenic plants overexpressing key C4enzymes not only had higher photosynthetic rates,but produced higher grain yields than WTplants.Given that the WT cv.Kitaake and the two transgenic plants(PPDK and PCK)have the same genetic background and the only difference is in the expression level of several C4key enzymes,our results provided direct evidence that increasing photosynthesis could result in a yield increase.
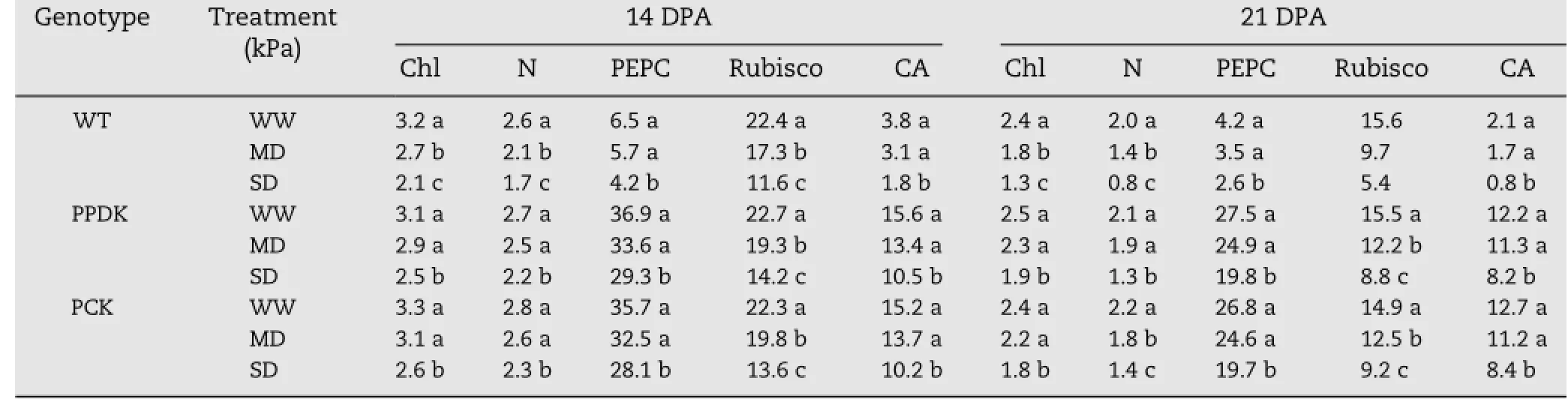
Table 3–Chlorophyll content(Chl,mg g−1DW),nitrogen content(N,%),and activities(μmol mg−1protein min−1)of phosphoenolpyruvate carboxylase(PEPC),ribulose-1,5-bisphosphate carboxylase(Rubisco),and carbonic anhydrase(CA)in the flag leaf at 14 and 21 days post-anthesis(DPA)in the field experiment.
The present results agree well with previous reports that transgenic rice plants show improved Pnand yield[4,10]. Transgenic plants showed Pnsuperior to the WT throughout the day and across the different grain filling stages(e.g.at 14 and 21 DPA).Ku et al.[4]observed that improved Pnwas caused by increased stomatal conductance and increased internal CO2concentration.In our study,higher leaf water content was observed for transgenic plants than for WT plants.The high leaf water content would contribute to increased stomatal conductance[42].Increased stomatal conductance may also be accounted for by the directregulation by C4enzymes such as PEPC of organic acid metabolism in guard cell to influence stomatal conductance [43,44].
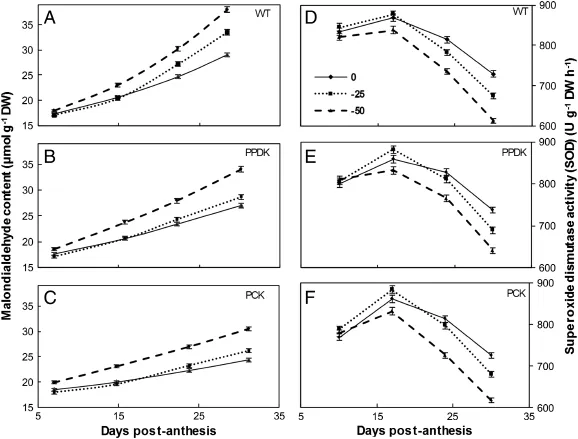
Fig.2–Malondialdehyde(MDA)content(A–C)and superoxide dismutase(SOD)activity(D–F)of wild-type(WT,cv.Kitaake)and two transgenic rice plants overexpressing pyruvate orthophosphate dikinase(PPDK);phosphoenolpyruvate carboxylase (PEPC),and PPDK(PCK),under well-watered(WW,solid lines),moderate drought(MD,dotted lines)and severe drought(SD, dashed lines)treatments in the tank experiment.The data represent means of five measurements from independent samples.
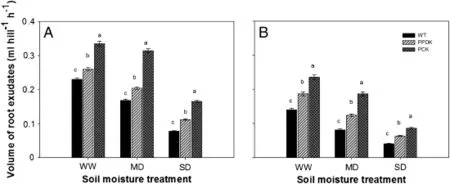
Fig.3–Volume of root exudates of wild-type(WT,cv.Kitaake)and two transgenic rice plants overexpressing pyruvate orthophosphate dikinase(PPDK);phosphoenolpyruvate carboxylase(PEPC),and PPDK(PCK),under different soil moisture treatments(well-watered,WW;moderate drought,MD;severe drought,SD),at(A)14 days post-anthesis(DPA)and(B)28 DPA in the tank experiment.Data represent means of five plants per treatment±standard error.Letters a–c above bars indicate significant differences at P=0.05 among different genotypes under the same soil moisture treatment.
Activities of PEPC and CA in transgenic plants were 3–5-fold higher than those in WT plants and decreased much less than did Rubisco activities under both MD and SD treatments.We speculate that the enzymes involved in C4photosynthesis are more tolerant to drought than those involved in C3photosynthesis.
Interestingly,we observed that the transgenic plants exhibited higher root activities than the wild type,as reflected by larger volumes of root exudates and higher root oxidation activity.High root activity would accelerate the absorption of water and nutrients from soil and exert feed-forward effects on leaf-level traits,resulting in higher leaf water content and photosynthetic rate and more active oxygen-scavenging systems in leaves of transgenic plants.Previous reports have also suggested the importance of root activity for maintaining higher source capacity and sink activity[45,46].The results suggest that improved root–shoot interaction in transgenic plants is one of the factors contributing to the increase in grain yield.Although the PCK transgenic plant showed higher root activities than PPDK(Figs.3 and 4),the expected advantages of PCK over PPDK in photosynthesis and yield were not observed,indicating a need for further investigation.
Enzymes involved in C4photosynthesis are known to increase in leaves of both C3and C4plants under abiotic stress [47–51].These enzymes play important roles in plant response to drought[4,15,45].For example,these enzymes can effectively reduce reactive oxygen species and membrane lipid peroxidation[15,18,19,51]an activity confirmed in our experiments (Fig.2,Table 3).This activity could account for the enhanced tolerance to drought shown by transgenic plants overexpressing these C4photosynthesis enzymes.
Usually drought reduces transpiration and simultaneously photosynthesis.We observed,however,that the extent to which the photosynthetic rate was reduced by the drought was much lower in transgenic than in WT plants and that the reduction of photosynthetic rate was lower than that of transpiration under drought,leading to increased transpiration efficiency(TE)for the transgenic plants(Table 2).This finding may have great significance for improving both grain yield and water use efficiency by transgenic approaches.
It is noteworthy that the WT cultivar Kitaake used in our study had a very low yield(2.15 t ha−1under the well-watered field condition);further studies should be conducted with high-yielding modern rice cultivars.However,transgenic plants showed a greater percentage of filled grains than WT plants,especially under the soil drought treatments(Table 5).It has been reported that the main limitation on yield potentialfor modern high-yielding cultivars,especially for the new“super rice”cultivars,is poor grain filling[51].It may be that overexpressing C4enzymes in these cultivars will increase source activity,thereby improving grain filling.
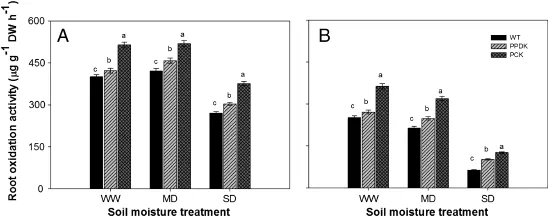
Fig.4–Root oxidation activity of wild-type(WT,cv.Kitaake)and two transgenic rice plants overexpressing pyruvate orthophosphate dikinase(PPDK);phosphoenolpyruvate carboxylase(PEPC),and PPDK(PCK),under different soil moisture treatments(well-watered,WW;moderate drought,MD;severe drought,SD),at(A)14 days post-anthesis(DPA)and(B)28 DPA in a tank experiment.Data represent means of five plants per treatment±standard error.Letters a–b above bars indicate significant differences at P=0.05 among different genotypes under the same soil moisture treatment.

Table4–Biomassofwildtyperice(WT)andtwo transgenic rice(PPDK and PCK)at 9 days post-anthesis (DPA)and maturity under different soil moisture treatments.
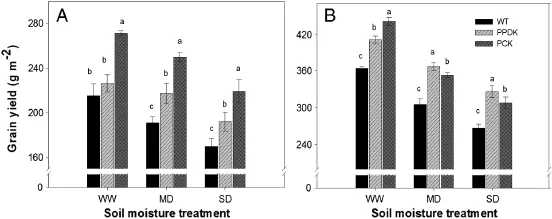
Fig.5–Yield of wild-type(WT,cv.Kitaake)and two transgenic rice plants overexpressing pyruvate orthophosphate dikinase (PPDK);phosphoenolpyruvate carboxylase(PEPC),and PPDK(PCK)under different soil moisture treatments(well-watered, WW;moderate drought,MD;severe drought,SD)in a field experiment(A)and a cement tank experiment(B).Grain yield was determined from a harvest area of 5.0 m2in the field experiment and 2.0 m2in the tank experiment.Data are means of three replicates per treatment±standard error.Letters a–c above bars indicate significant differences at P=0.05 among different genotypes under the same soil moisture treatment.
It should also be noted that the C4photosynthetic pathway is a set of complex physiological and biochemical processes. Some researchers argue that the presence of Kranz leaf anatomy is essential for C4photosynthesis function.Enzymes involved in the C4pathway are compartmentalized between the mesophyll and bundle sheath cells[52].But a single-cell C4pathway has also been found[53],and the presence of a C4-mini cycle in C3plants has been reported[54,55].Overexpression of C4photosynthesis enzymes could strengthen the C4-mini cycle and contribute to improving C3photosynthesis [56].But the exact mechanism of carbon assimilation at the molecular and biochemical level awaits elucidation.
5.Conclusions
Transgenic rice plants overexpressing C4photosynthesis enzymes(PPDK and PCK)exhibited higher grain yields than WTplants,especially under soildroughtconditions.Better yield performance and higher drought tolerance of the transgenic rice were associated with greater photosynthetic rate in leaves, higher leaf water content,chlorophyll and nitrogen content, transpiration efficiency,PEPC and CA activities in leaves,higher root oxidation activity,and a stronger active oxygen scavenging system.These results provide experimental evidence thattransgenic rice plants overexpressing C4photosynthesis enzymes may show improved grain yield,especially under drought environments–a finding that may open a new avenue to physiological breeding under drought by means of overexpressing C4enzymes in C3crops such as rice.
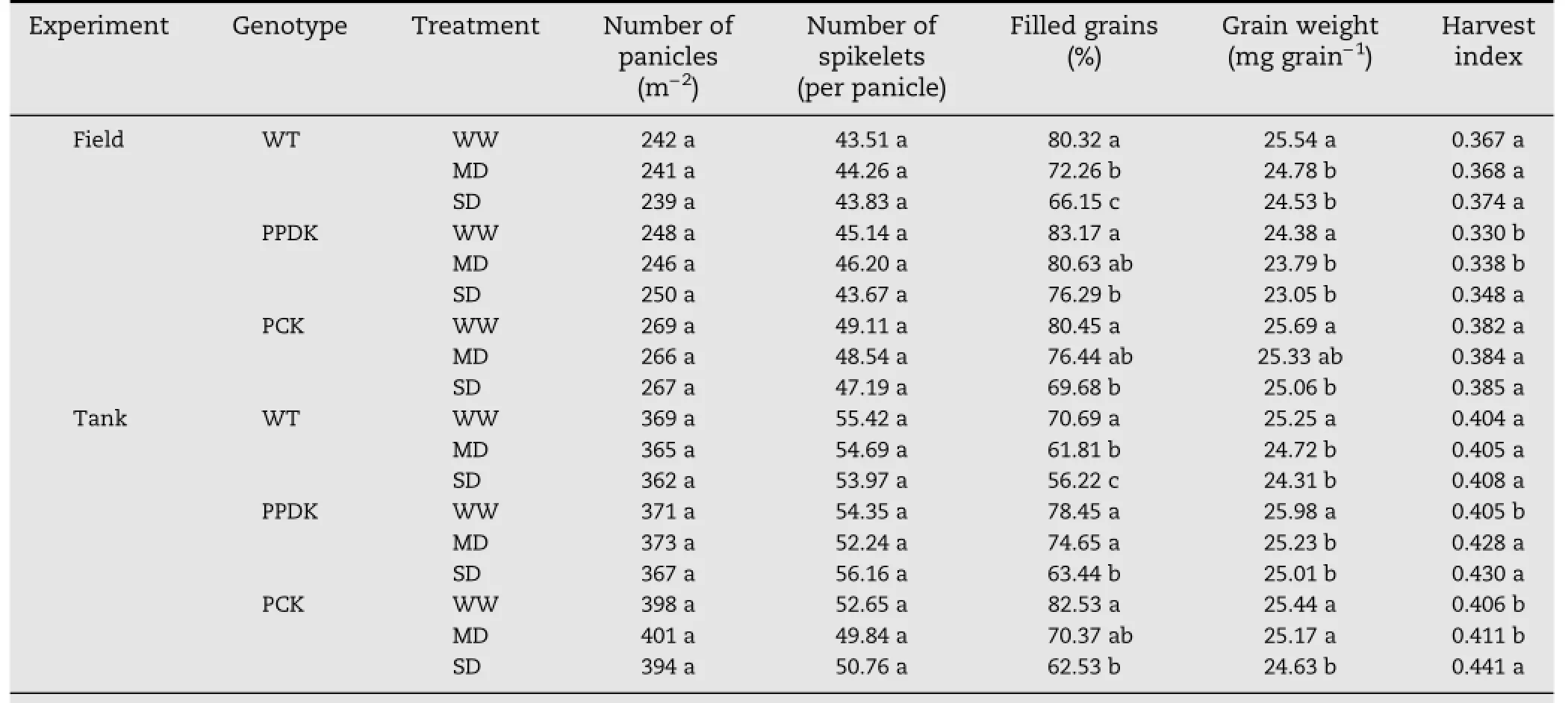
Table 5–Effect of soil drought on yield components and harvest index of wild type(WT)and transgenic(PPDK and PCK)rice.
Acknowledgments
We thank Prof.MSB Ku,School of Biological Sciences, Washington State University for providing transgenic rice materials overexpressing C4photosynthesis enzymes,and acknowledge grants from the National Basic Research Program (973 Program,2012CB114306),the National Natural Science Foundation of China(31061140457;31071360;31271641),the National Key Technology Support Program of China (2011BAD16B14;2012BAD04B08),China National Public Welfare Industry(Agriculture)Plan(200803030;201203079)and Jiangsu Advantages of Key Construction Projects(JS 2011).
R E F E R E N C E S
[1]W.L.Ogren,Photorespiration:pathways,regulation,and modification,Annu.Rev.Plant Physiol.35(1984)415–442.
[2]R.F.Sage,F.Kocacinar,D.S.Kubien,C4photosynthesis and temperature,in C4photosynthesis and related CO2concentrating mechanisms,in:A.S.Raghavendra,R.F.Sage (Eds.),Springer,Dordrecht,The Netherlands,2011,pp.161–195.
[3]R.F.Sage,X.Zhu,Exploiting the engine of C4photosynthesis, J.Exp.Bot.62(2011)2989–3000.
[4]M.S.Ku,D.Cho,U.Ranade,T.Hsu,X.Li,D.Jiao,J.Ehleringer, M.Miyao,M.Matsuoka,Photosynthetic performance of transgenic rice plants overexpressing maize C4photosynthesis enzymes,in:J.E.Sheehy,P.L.Mitchell,B.Hardy(Eds.), Redesigning Rice Photosynthesis to Increase Yield, Elsevier/IRRI,Amsterdam/Oxford,2000,pp.193–204.
[5]M.R.Schmitt,G.E.Edwards,Photosynthetic capacity and nitrogen use efficiency of maize,wheat,and rice:a comparison between C3and C4photosynthesis,J.Exp.Bot.32(1981)459–466.
[6]J.A.Langdale,C4cycles:past,present,and future research on C4photosynthesis,Plant Cell 23(2011)3879–3892.
[7]L.Taiz,L.Zeiger,Plant Physiology,5th edition Sinauer Associates,Inc.,Publishers,Sunderland,Massachusetts,2010.
[8]M.D.Hatch,C4photosynthesis,a unique blend of modified biochemistry,anatomy,and ultrastructure,Biochim. Biophys.Acta 895(1987)81–106.
[9]R.E.Häusler,H.J.Hirsch,F.Kreuzaler,C.Peterhänsel, Overexpression of C4-cycle enzymes in transgenic C3plants: a biotechnological approach to improve C3-photosynthesis, J.Exp.Bot.53(2002)591–607.
[10]M.S.Ku,S.Agarie,M.Nomura,H.Fukayama,H.Tsuchida,K. Ono,S.Hirose,S.Toki,M.Miyao,M.Matsuoka,High-level expression of maize phosphoenolpyruvate carboxylase in transgenic rice plants,Nat.Biotechnol.171(1999)76–80.
[11]S.von Caemmerer,W.P.Quick,R.T.Furbank,The development of C4rice:current progress and future challenges,Science 336 (2012)1671–1672.
[12]C.Peterhansel,Bestpractice procedures forthe establishmentof a C4cycle in transgenic C3plants,J.Exp.Bot.62(2011)3011–3019.
[13]Y.Taniguchi,H.Ohkawa,C.Masumoto,T.Fukuda,T.Tamai, K.Lee,S.Sudoh,H.Tsuchida,H.Sasaki,H.Fukayama, Overproduction of C4photosynthetic enzymes in transgenic rice plants:an approach to introduce the C4-like photosynthetic pathway into rice,J.Exp.Bot.59(2008)1799–1809.
[14]M.Miyao,C.Masumoto,S.I.Miyazawa,H.Fukayama,Lessons from engineering a single-cell C4photosynthetic pathway into rice,J.Exp.Bot.62(2011)3021–3029.
[15]D.Jiao,X.Huang,X.Li,W.Chi,T.Kuang,Q.Zhang,M.S.Ku,D. Cho,Photosynthetic characteristics and tolerance to photo-oxidation of transgenic rice expressing C4photosynthesis enzymes,Photosynth.Res.72(2002)85–93.
[16]S.Agarie,A.Miura,R.Sumikura,S.Tsukamoto,A.Nose,S. Arima,M.Matsuoka,M.Miyao-Tokutomi,Overexpression of C4PEPC caused O2-insensitive photosynthesis in transgenic rice plants,Plant Sci.162(2002)257–265.
[17]M.Miyao,Molecular evolution and genetic engineering of C4photosynthetic enzymes,J.Exp.Bot.54(2003)179–189.
[18]B.Y.Zhou,Z.S.Ding,M.Zhao,Alleviation of drought stress inhibition on photosynthesis by overexpression of PEPC in rice,Acta Agron.Sin.37(2011)112–118(in Chinese with English abstract).
[19]Z.S.Ding,B.Y.Zhou,X.F.Sun,M.Zhao,High light tolerance is enhanced by overexpressed PEPC in rice under drought Stress,Acta Agron.Sin.38(2012)285–292.
[20]M.Jeanneau,D.Gerentes,X.Foueillassar,M.Zivy,J.Vidal,A. Toppan,P.Perez,Improvement of drought tolerance in maize:towards the functional validation of the Zm-Asr1 gene and increase of water use efficiency by over-expressing C4–PEPC,Biochimie 84(2002)1127–1135.
[21]A.Bandyopadhyay,K.Datta,J.Zhang,W.Yang,S. Raychaudhuri,M.Miyao,S.Datta,Enhanced photosynthesis rate in genetically engineered indica rice expressing pepc gene cloned from maize,Plant Sci.172(2007)1204–1209.
[22]Holden M.Chlorophyll,in:T.W.Goodwin(Ed.),2nd edn, Chemistry and Biochemistry of Plant Pigments,vol.2, Academic Press,London and,New York,1976,pp.1–37.
[23]S.Yoshida,D.Forno,J.Cock,K.Gomez,Laboratory Manual for Physiological Studies of Rice,International Rice Research Institute,The Philippines,1976.14–17.
[24]D.H.Gonzalez,A.A.Iglesias,C.S.Andeo,On the regulation of phosphoenolpyruvate carboxylase activity from maize leaves by L-malate:effects of pH,J.Plant Physiol.116(1984)425–429.
[25]J.C.Wei,R.L.Wang,G.Y.Cheng,Studies on the kinetic properties of ribulose-1,5-bisphosphate carboxylase from F1hybrid rice,Acta Phytosiol.Sin.20(1994)56–60(in Chinese with English abstract).
[26]M.L.Guo,Y.Z.Gao,Z.Wang,Determination of plant carbonic anthydrase activity with pH-meter,Plant Physiol.Commun.6 (1988)59–61(in Chinese with English abstract).
[27]C.N.Giannopolitis,S.K.Ries,Superoxide dismutases:I. Occurrence in higher plants,Plant Physiol.59(1977)309–314.
[28]S.Zhao,C.Xu,Q.Zou,Q.Meng,Improvements of method for measurement of malondiadehyde in plant tissues,Plant Physiol.Commun.30(1994)207–210(in Chinese with English abstract).
[29]F.Asch,M.Dingkuhn,A.Sow,A.Audebert,Drought-induced changes in rooting patterns and assimilate partitioning between root and shoot in upland rice,Field Crop Res.93 (2005)223–236.
[30]C.Yang,L.Yang,Y.Yang,Z.Ouyang,Rice root growth and nutrient uptake as influenced by organic manure in continuously and alternately flooded paddy soils,Agric. Water Manag.70(2004)67–81.
[31]S.Yoshida,D.Forn,J.Cock,K.Gomez,Laboratory Manual for Physiological Studies of Rice,International Rice Research Institute,The Philippines,1976.74–77.
[32]J.Gu,X.Yin,P.C.Struik,T.J.Stomph,H.Wang,Using chromosome introgression lines to map quantitative trait loci for photosynthesis parameters in rice(Oryza sativa L.) leaves under drought and well watered field conditions, J.Exp.Bot.63(2012)455–469.
[33]J.Gu,X.Yin,T.J.Stomph,H.Wang,P.C.Struik,Physiological basis of genetic variation in leaf photosynthesis among rice(Oryza sativa L.)introgression lines under drought and well-watered conditions,J.Exp.Bot.63(2012)5137–5153.
[34]P.L.Mitchell,J.E.Sheehy,Supercharging rice photosynthesis to increase yield,New Phytol.171(2006)688–693.
[35]H.Fukayama,M.D.Hatch,T.Tamai,H.Tsuchida,S.Sudoh, R.T.Furbank,M.Miyao,Activity regulation and physiological impacts of maize C4-specific phosphoenolpyruvate carboxylase overproduced in transgenic rice plants, Photosynth.Res.77(2003)227–239.
[36]Y.M.Wang,W.G.Xu,L.Hu,L.Zhang,Y.Li,X.H.Du,Expression of maize gene encoding C4-pyruvate orthophosphate dikinase (PPDK)and C4-phosphoenolpyruvate carboxylase(PEPC)in transgenic Arabidopsis,Plant Mol.Biol.Report.30(2012)1367–1374.
[37]L.T.Evans,R.L.Dunstone,Some physiological aspects of evolution in wheat,Aust.J.Biol.Sci.23(1970)725–742.
[38]L.Borrás,G.A.Slafer,M.E.Otegui,Seed dry weight response to source–sink manipulations in wheat,maize and soybean: a quantitative reappraisal,Field Crop Res.86(2004)131–146.
[39]S.P.Long,X.G.Zhu,S.L.Naidu,D.R.Ort,Can improvement in photosynthesis increase crop yields?Plant Cell Environ.29 (2006)315–330.
[40]C.E.Jahn,J.Mckay,R.Mauleon,J.Stephens,K.L.McNally,D.R. Bush,H.Leung,J.E.Leach,Genetic variation in biomass traits among 20 diverse rice varieties,Plant Physiol.155(2011)157–168.
[41]J.Gu,X.Yin,T.J.Stomph,P.C.Struik,Can exploiting natural genetic variation in leaf photosynthesis contribute to increasing rice productivity?A simulation analysis,Plant Cell Environ.(2013),http://dx.doi.org/10.1111/pce.12173,(in press).
[42]T.Lawson,S.von Caemmerer,I.Baroli,Photosynthesis and stomatal behavior,in:U.Lüttge,W.Beyschlag,B.Büdel,D. Francis(Eds.),Progress in Botany,vol.72,Springer, Berlin/Heidelberg,2011.
[43]A.Vavasseur,A.S.Raghavendra,Guard cell metabolism and CO2sensing,New Phytol.165(2005)665–682.
[44]V.Cotelle,J.N.Pierre,A.Vavasseur,Potential strong regulation of guard cell phosphoenolpyruvate carboxylase through phosphorylation,J.Exp.Bot.50(1999)777–783.
[45]S.Ramasamy,H.F.M.Ten Berge,S.Purushothaman,Yield formation in rice in response to drainage and nitrogen application,Field Crop Res.51(1997)65–82.
[46]H.Zhang,S.Zhang,J.Yang,J.Zhang,Z.Wang,Postanthesis moderate wetting drying improves both quality and quantity of rice yield,Agron.J.100(2008)726–734.
[47]T.Fushimi,M.Umeda,T.Shimazaki,A.Kato,K.Toriyama,H. Uchimiya,Nucleotide sequence of a rice cDNA similar to a maize NADP-dependent malic enzyme,Plant Mol.Biol.24 (1994)965–967.
[48]M.H.Walter,J.Grima-Pettenati,C.Feuillet,Characterization of a bean(Phaseolus vulgaris L.)malic-enzyme gene,Eur.J. Biochem.224(1994)999–1009.
[49]J.Schaaf,M.H.Walter,D.Hess,Primary metabolism in plant defense(regulation of a bean malic enzyme gene promoter in transgenic tobacco by developmental and environmental cues),Plant Physiol.108(1995)949–960.
[50]S.Liu,Y.Cheng,X.Zhang,Q.Guan,S.Nishiuchi,K.Hase,T. Takano,Expression of an NADP-malic enzyme gene in rice(Oryza sativa L.)is induced by environmental stresses;over-expression of the gene in Arabidopsis confers salt and osmotic stress tolerance,Plant Mol.Biol.64(2007) 49–58.
[51]J.Yang,J.Zhang,Grain-filling problem in‘super'rice,J.Exp. Bot.61(2010)1–5.
[52]S.Covshoff,J.M.Hibberd,Integrating C4photosynthesis into C3crops to increase yield potential,Curr.Opin.Biotechnol.23 (2012)209–214.
[53]G.Bowes,K.R.Srinath,M.E.Gonzalo,B.R.Julia,C4mechanisms in aquatic angiosperms:comparisons with terrestrial C4systems,Funct.Plant Biol.29(2002)379–392.
[54]J.M.Hibberd,W.P.Quick,Characteristics of C4photosynthesis in stems and petioles of C3flowering plants,Nature 24(2002) 451–454.
[55]B.H.Ji,S.Q.Zhu,D.M.Jiao,Photosynthetic C4-microcycle in transgenic rice plant lines expressing the maize C4-photosynthetic enzymes,Acta Agron.Sin.30(2004) 536–543(in Chinese with English abstract).
[56]L.Hu,Y.Li,W.G.Xu,Q.C.Zhang,L.Zhang,X.L.Qi,H.B.Dong, Improvement of the photosynthetic characteristics of transgenic wheat plants by transformation with the maize C4phosphoenolpyruvate carboxylase gene,Plant Breed.131 (2012)385–391.
*Corresponding author.
E-mail address:jcyang@yzu.edu.cn(J.-C.Yang).
Peer review under responsibility of Crop Science Society of China and Institute of Crop Science,CAAS.

- The Crop Journal的其它文章
- QTL mapping of starch granule size in common wheat using recombinant inbred lines derived from a PH82-2/Neixiang 188 cross
- Identification and functional analysis of miRNAs in developing kernels of a viviparous mutant in maize
- Molecular approaches unravel the mechanism of acid soil tolerance in plants
- Enhanced tolerance to drought in transgenic rice plants overexpressing C4 photosynthesis enzymes
- HrcQ is necessary for Xanthomonas oryzae pv.oryzae HR-induction in non-host tobacco and pathogenicity in host rice
- Isolation and characterization of an isoamylase gene from rye