
建植第1年刈割对黑麦草+白三叶草地杂草空间格局的影响
2013-04-25 09:41:28罗天琼王普昶赵丽丽王应芬
草业科学 2013年8期
罗天琼,王普昶,赵丽丽,牟 琼,王应芬,黎 俊
(1.贵州省草业研究所,贵州 贵阳 550006; 2.贵州大学动物科学学院,贵州 贵阳 550025;3.贵州牧草种籽繁殖场,贵州 独山 558202)
栽培草地是采用农业技术措施栽培而成的草地,其目的是获得高产优质的牧草,以补充天然草地的不足,满足家畜的饲料需求。足够的栽培草地对减少家畜因冬、春饲料不足而掉膘或死亡损失,增加畜产品产量和提高土地利用率等均有重要意义[1]。因此,栽培草地面积的多少,常是衡量一个地区或国家畜牧业发达程度的重要标志[2]。1980年以来,随着我国南方草地资源的开发利用,贵州、云南、湖南、湖北等地成为大面积栽培草地的重点推广区域。随着我国栽培草地的大面积推广,栽培草地退化快、利用年限短等问题逐渐受到人们的关注。特别是杂草大量入侵,导致栽培草地的特征基本消失,严重抑制了草地畜牧业的发展[3-4]。因此,栽培草地的稳定性是草地建植与管理的核心问题之一,而草地抗杂草入侵是其稳定性的重要表现形式。生物量是指个体、种群或群落生物数量的多少,是度量个体、种群的大小以及个体、种群在群落中的地位和作用的指标,同时也是反映群落或生态系统功能强弱的重要指标。关于栽培草地系统杂草入侵对刈割强度的响应已有一些学者从不同角度进行了研究[5-8],但从空间异质性角度探讨刈割影响下草地群落杂草入侵生物量分布规律的研究还鲜有报道。
“地统计学”一词首先由G.Matheron提出,后来在生态学中得到广泛的发展与应用[9-10]。从20世纪70年代开始,应用地统计学理论研究生态因子空间异质性规律被国外一些学者所使用。20世纪80年代初,我国才开始从事这方面的研究。20世纪90年代以后,对土壤、植被、水分等方面的空间异质性的研究较多[11-14,15-16]。但以往的研究多集中在天然植被、土壤系统方面,针对小尺度栽培草地系统杂草空间分布的报道相对较少。本研究应用地统计分析方法研究不同刈割强度下多年生黑麦草(Loliumperenne)+白三叶(Trifoliumrepens)混播草地杂草地上生物量的空间分布格局与异质化程度,并利用Kriging法绘制杂草地上生物量的空间分布图,从而探讨地上杂草的异质化与其在刈割强度下的生态学过程,旨在为栽培草地的建植和持续利用提供基础资料和依据。
1 材料与方法
1.1试验地概况 试验地位于贵阳市花溪区西郊农场,26°15′ N,106°40′ E,海拔1 000 m,该区为典型的喀斯特地貌。年均气温14.9 ℃,年平均降水量1 100~1 200 mm,且主要集中在5-7月,属高原湿润季风气候。主要植被为亚热带石灰岩乔灌丛,土壤为壤质黄壤,0~30 cm土层pH值为5.0~6.2,速效氮16.2 mg·kg-1,速效磷3.6 mg·kg-1,速效钾53.4 mg·kg-1,有机质0.810 3 mg·kg-1[7]。进行试验的多年生黑麦草+白三叶草混播草地建植于2011年4月,其杂草主要有狗尾草(Setairaviridis)、荩草(Arthraxonhispidus)、马唐(Digitariasanguinalis)、小藜(Chenopodiumserotinum)、丛枝蓼(Polygonumcaespitosum)、铁苋菜(Acalyphaaustralis)、无芒稗(Echinochloacrusgalivar.mitis)、画眉草(Eragrostispilosa)、百脉根(Lotuscornioulatus)、鸭茅(Dactylisglomerata)、平车前(Plantagodepressa)和雀稗(Paspalumscrobiculatum)等20余种。
1.2试验设计与试验方法
1.2.1空间取样设计和分析 试验设对照(不刈割)、轻度刈割(留茬10 cm)、中度刈割(留茬5 cm)和重度刈割(留茬2 cm) 4个处理,每处理3次重复。于2011年4月进行人工撒播,播后围封管理,至样方整体株高40 cm时进行刈割处理。在试验区不同处理地段布设面积为4 m×4 m的样方,完全随机设计,样方间隔2 m。然后采用邻接格子样方法,在大样方内将其分割成100个20 cm×20 cm的小样方作为最小的格子单位,于2011年8月末记录样方内每个格子的坐标并齐地刈割所有格子内的杂草,带回实验室。将样品置于80 ℃下烘至质量恒定,后称量每个格子内的杂草总生物量干质量。
1.2.2数据处理 利用地统计软件(GS+9.0)的半方差函数分析植物个体生物量的空间变异特征,并把样方内杂草生物量的点坐标数据转变为连续空间变量进行实际变异函数值的计算和理论曲线的拟合[12-14]。
2 结果与分析
2.1不同刈割强度条件下栽培草地杂草地上生物量的空间异质性分析 根据4个刈割处理条件下多年生黑麦草+白三叶混播草地杂草地上生物量的数据,对其进行半方差函数分析及模型拟合得到各处理杂草地上生物量变异函数模型(表1,图1)。可以看出,4个刈割处理条件下多年生黑麦草+白三叶混播草地的杂草地上生物量均能较好地拟合为指数模型和高斯模型,且4个刈割处理条件下杂草生物量的决定系数均较高,具有明显的空间结构特征。
由变异函数的基台值可知,重度刈割条件下基台值最高,达12.530,这说明重度刈割条件下,栽培草地杂草地上生物量的空间分布格局异质性最强。中度刈割条件下基台值最低,为3.149,表明其格局异质性很低。轻度刈割和对照条件下基台值居中,分别为4.557和4.751,表明植被地上生物量的空间分布格局异质性较强。
植被地上生物量空间相关范围分析表明,重度刈割条件下空间相关范围最大,为1 004.1 cm;中度刈割条件下最低,为122.1 cm;轻度刈割和对照条件下居中,分别为161.9和219.1 cm。
结构方差比可用来衡量空间自相关程度。空间变异比(表1)分析表明,不同刈割强度条件下栽培草地杂草地上生物量的结构方差比变化介于0.503~0.953,空间自相关引起的空间异质性均超过50%,空间自相关程度达到中等。从轻度刈割到中度刈割、重度刈割条件下,引起的空间异质性随机因素呈增-减-增的变化趋势,空间自相关因素呈减-增-减的变化趋势。因此,空间自相关因素是不同刈割强度条件下栽培草地杂草地上生物量的杂草地上生物量空间异质性的主要因素。
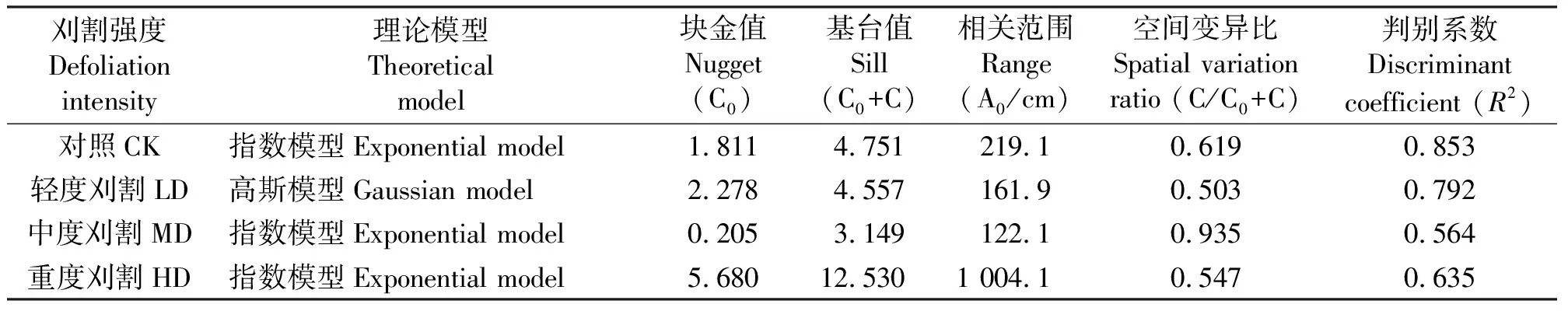
表1 不同刈割强度条件下黑麦草栽培草地杂草地上生物量空间变异的半方差特征参数Table 1 Semi-variogram model parameters of the aboveground biomass of invading specie under different mowing intensity
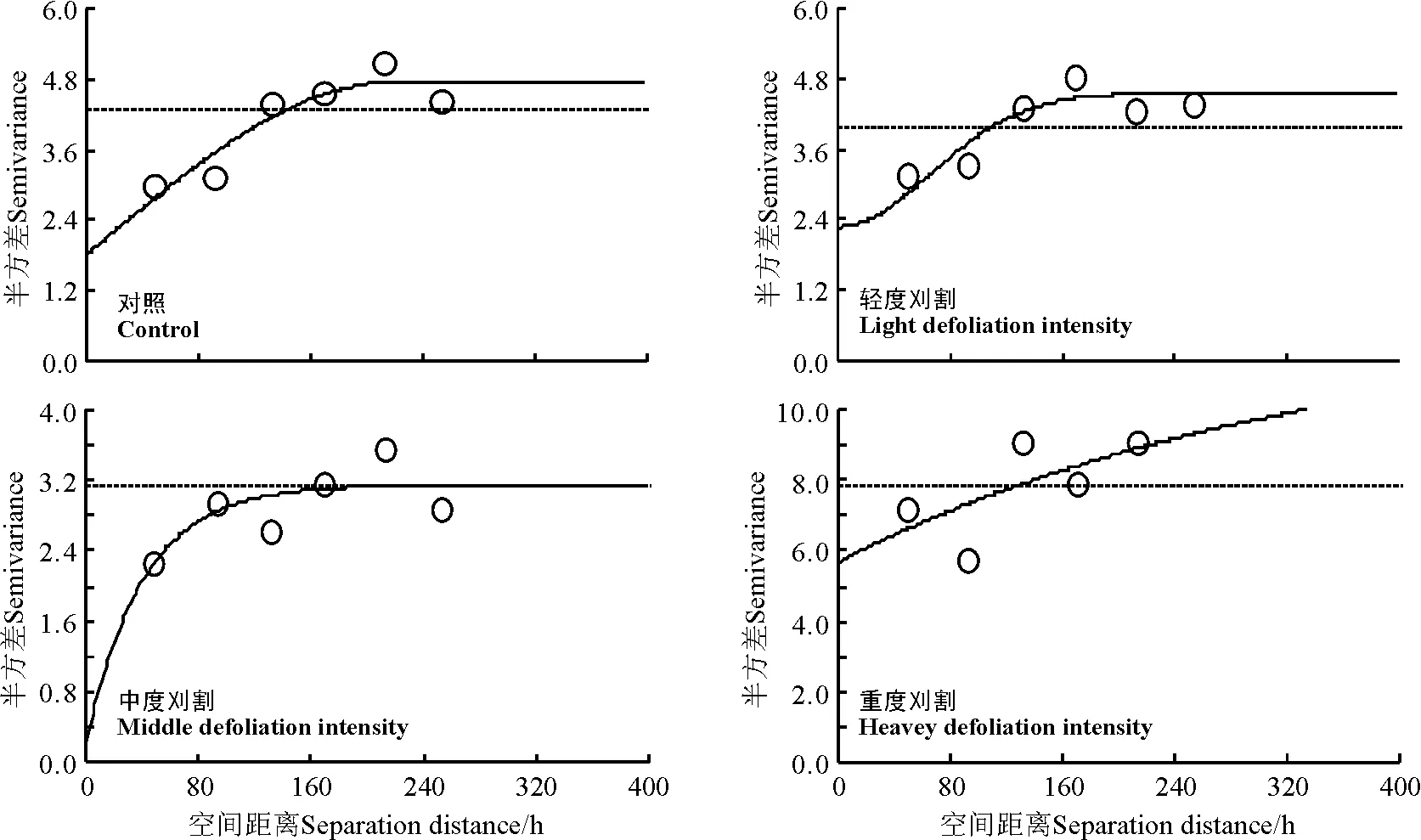
图1 不同刈割强度条件下黑麦草草地杂草地上生物量变异函数散点图及变异曲线Fig.1 Semi-variogram of spatial pattern for the aboveground biomass of invading specie under different mowing intensity
2.2不同刈割强度条件下栽培草地杂草地上生物量的克里格分析 由Kriging maps可以看出,不同刈割强度条件下栽培草地杂草地上生物量的空间格局和变化过程较显著(图2)。未进行刈割条件下栽培草地杂草地上生物量具有较大尺度格局,斑块的数量较少、面积较大;中、高生物量的斑块占主要部分,低生物量的斑块较少,且呈聚集分布。轻度刈割草地上,杂草地上生物量格局减弱,表现为斑块的面积减少,斑块数量增多,斑块的等高线数量和密度增加;高生物量的杂草个体稍有减少,低生物量的杂草占较多数。格局斑块中高生物量的斑块减少。中度刈割条件下栽培草地杂草地上生物空间格局表现为斑块较小,斑块破碎化严重;大部分斑块为中、低生物量的斑块,这与其杂草个体生物量低,且个体之间生物量差异不大有关。重度刈割条件下栽培草地,杂草地上生物量分布格局显著增强,表现为格局的尺度减小,斑块的面积减小、数量增多。低生物量斑块和高生物量斑块中生物量相差较大,且呈聚集分布。
3 讨论与结论
栽培草地群落抗杂草入侵是其功能的重要表现形式,也是栽培草地稳定性的重要理论基础。栽培草地建植后必须及时地施以适度放牧利用、刈割、施肥等措施,这些措施(干扰)极大地影响着植物群落的结构、功能及动态[17-18]。因此,如何有效地通过干扰调控的手段实现栽培草地的稳定性、持久性利用是草业科学的研究热点。关于栽培草地杂草调控方面,研究栽培禾草与杂草生物量在空间上的相互作用过程及其机理意义重大[18]。
不同刈割强度条件下多年生黑麦草+白三叶混播草地杂草地上生物量的空间格局异质性程度差异很大,这是由于栽培草地的建立破坏原有植被的自然平衡[3,16]。在无任何农业管理措施的多年生栽培草地上,杂草和栽培禾草的生长表现主要取决于二者的相对竞争力。栽培草地建植初期,杂草出苗早,生长快,初始占空能力较强,相对竞争力高于栽培禾草,从而在生长竞争中占据了优势地位[3,17]。植被分布格局也由均质化向异质化发展,从而加速了栽培草地的杂草化过程。
要维持栽培草地长期处于稳定状态,就必须使其一直处于适宜的干扰之下。刈割是控制杂草的较好手段之一[5-8],本研究从空间异质性角度分析刈割影响下杂草的入侵动态。空间异质性的组成分析中,块金值(C0)表示随机部分的空间异质性;拱高(C)表示系统变异的空间异质性;基台值(C0+C)表示系统内总的变异,是系统或系统属性中最大的变异[11,19-20];变程(A0)表示区域化变量在各个方向上自相关平均影响范围的定量指标[19-20]。本研究发现,在不同刈割强度中,基台值表现为重度刈割最高、中度刈割最低、轻度刈割和对照居中;植被地上生物量空间相关范围(A0)则是重度刈割最大、轻度刈割和对照居中、中度刈割最低,这表明不同刈割强度条件下多年生黑麦草栽培草地入侵杂草地上生物量空间格局异质化程度及其相关范围变化非常明显。重度刈割条件下多年生黑麦草+白三叶混播草地杂草地上生物量的空间分布格局异质性最强,中度刈割条件下格局异质性很低。结构方差比可用来衡量空间自相关程度。不同刈割强度条件下栽培草地杂草地上生物量空间自相关引起的空间异质性均超过50%,从轻度刈割到中度刈割、重度刈割条件下,引起的空间异质性随机因素呈增-减-增的变化趋势,空间自相关因素呈减-增-减。空间分布格局图分析进一步显示,不同刈割强度条件下多年生黑麦草+白三叶混播草地杂草地上生物量格局变化显著。本研究仅分析了多年生黑麦草+白三叶混播草地建植初期刈割强度对其杂草生物量空间异质性分布的影响,并初步判定中度刈割条件下栽培草地格局异质性低的结论,其潜在的生物学机制是补偿效应作用的结果。而有些控制试验得出刈割强度愈大,杂草愈容易入侵的结论,这是否是因为栽培草地对多年自然选择效应还是别的原因还有待进一步研究。
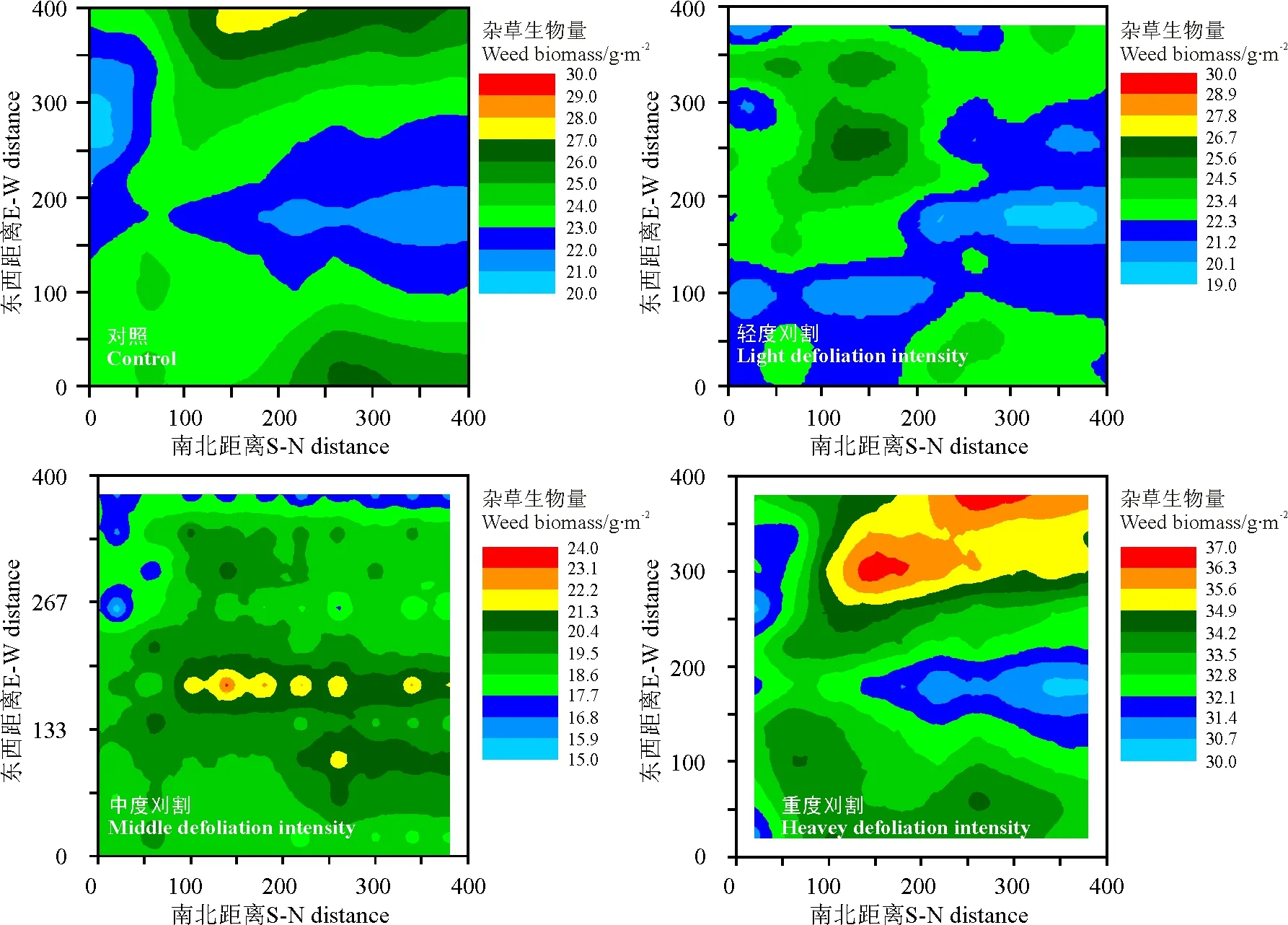
图2 不同刈割强度条件下黑麦草草地杂草地上生物量等值线和空间分布格局Fig.2 Kriging maps of spatial pattern for the aboveground biomass of weed under different mowing intensities
[1] 胡自治.人工草地在我国21世纪草业发展和环境治理中的重要意义[J].草原与草坪,2000(1):12-16.
[2] 江小雷,张卫国,严林,等.植物群落物种多样性对生态系统生产力的影响[J].草业学报,2004,13(6):8-13.
[3] 王刚,吴明强,蒋文兰.人工草地杂草生态学研究——杂草入侵与放牧强度之间的关系[J].草业学报,1995,4(3):75-80.
[4] 李昂,顾梦鹤,张世挺,等.青藏高原东缘人工草地群落物种丰富度对杂草入侵的影响[J].生态学杂志,2009,28(2):177-181.
[5] 邱正强,马玉寿,施建军,等.刈割频次与留茬高度对人工草地生产力的影响[J].青海畜牧兽医杂志,2008,38(5):12-14.
[6] 张卫国,江小雷,杨振宇.多花黑麦草在高寒牧区的引种研究[J].草业学报,2004,13(2):51-56.
[7] 王普昶,牟琼,王晓力,等.贵州黑麦草人工草地杂草入侵对刈割强度的响应[J].中国草地学报,2011,33(4):117-121.
[8] 锡文林,张仁平.混播比例和刈割期对混播草地产草量及种间竞争的影响[J].中国草地学报,2009,31(4):36-41.
[9] Legender P,Fortin M J.Spational pattern and ecological analysis[J].Vegetation,1989,80:107-138.
[10] Levin S A.The problem of patten and scale in ecology[J].Ecology,1992,73:1943-1967.
[11] 王晓春,韩士杰,邹春静,等.长白山岳桦种群格局的地统计学分析[J].应用生态学报,2002,13(3):781-784.
[12] 辛晓平,李向林,杨桂霞,等.放牧和刈割条件下草山草坡群落空间异质性分析[J].应用生态学报,2002,13(4):449-452.
[13] 刘先华,韩苑鸿,李凌浩.放牧率对内蒙古典型草原物种分布空间异质性的影响[J].草地学报,1998,6(4):293-299.
[14] 程晓莉,安树青,钦佩,等.鄂尔多斯草地退化过程中植被地上生物量空间分布的异质性[J].生态学报,2003,23(8):1526-1533.
[15] 赵丽丽,王普昶,张锦华,等.贵州喀斯特暖性草丛土壤物理因子空间异质性分析[J].草业科学,2011,28(7):1234-1238
[16] 吕杰,郭君喜,陈俊,等.不同草地植被群落的结构特征研究[J].草业科学,2011,28(6):1058-1065.
[17] 杜峰,梁宗锁,徐学选,等.陕北黄土丘陵区撂荒群落土壤养分与地上生物量空间异质性[J].生态学报,2008,28(1):13-22.
[18] 董世魁,龙瑞军,胡自治,等.高寒地区多年生禾草人工草地杂草种群动态研究[J].兰州大学学报(自然科学版),2003,39(5):82-87.
[19] 王政权.地统计学及在生态学中的应用[M].北京:科学出版社,1999:150-156.
[20] 李春喜,王志和,王文林.生物统计学[M].北京:科学出版社,2000:117.
猜你喜欢
奥秘(创新大赛)(2022年8期)2022-09-14 05:44:42
科教新报(2022年22期)2022-07-02 12:34:28
中国果业信息(2021年10期)2021-12-07 04:06:06
今日农业(2020年19期)2020-12-14 14:16:52
养殖与饲料(2020年6期)2020-02-18 16:01:26
中国种业(2019年8期)2019-08-26 08:41:18
中国科技信息(2015年2期)2015-11-16 08:18:32
现代农业(2015年5期)2015-02-28 18:40:49
植物营养与肥料学报(2014年1期)2014-03-11 20:24:38
杂草学报(2012年1期)2012-11-06 07:08:33
