
喀纳斯景区山地草甸土壤理化性质与植物多样性━生产力关系
2013-04-25 10:13郑伟
草业科学 2013年12期
郑 伟
(1.新疆农业大学草业与环境科学学院,新疆 乌鲁木齐 830052;2.新疆维吾尔自治区草地资源与生态重点实验室, 新疆 乌鲁木齐 830052)
植物多样性是生物多样性的重要组成部分,是受环境中各种干扰影响的最直观表征者,也是抵抗各种干扰首当其冲者[1]。植物多样性包含着许多复杂的生物生态学信息[2-6]。在很大程度上,可通过对植物多样性的间接观察、了解甚至调控来改变生态系统的结构和功能[7-8]。关于植物多样性与生态系统功能的关系研究,主要集中在物种多样性、功能群多样性与生态系统过程[9]、生产力[10]、持续性[11]、稳定性[12]及外界干扰因素对其影响[13]等方面。另外,土壤等非生物因素与植物多样性和生态系统结构、功能的交互作用也受到很大关注[14-15]。Dybzinski等[16]认为,增加土壤养分(主要是N)的可利用性和持续性后,群落物种多样性增加可以使群落生产力得到提高。Reich等[17]和Dijkstra等[18]均认为增加土壤N的供应和CO2浓度后,物种多样性的增加可导致群落生产力提高,而且土壤N和CO2的这种增产效应是分离的。从另外一个方面来讲,高物种多样性也有利于土壤营养的保持[19-20]。因而,理解植物多样性、生产力与土壤理化性质之间的关系,对于解释植物多样性与生态系统结构、功能和过程之间的联系是有帮助的[21-22]。研究这种关系,还可增进人们对生命系统和地球系统界面之间相互作用的了解,甚至预测环境格局的变化[23]。但针对植物与土壤理化性质关系的研究主要集中在植物群落数量特征与土壤理化性质的相关关系[24]、植物多样性(特别是植物物种多样性)与土壤理化性质的相关关系[25-26]以及植物物种多样性与土壤保持功能关系[23]等方面,缺乏定量分析整个植物多样性体系(植物物种多样性、功能群多样性和群落多样性)及其生产力与土壤理化性质关系的研究。因此,本研究将植物物种多样性、功能群多样性和群落多样性及生产力与土壤理化性质的相关关系进行定量化分析,探讨旅游对植物多样性及其生产力与土壤理化性质关系以及对植物多样性与生态系统结构和功能维持机制的影响,为深入研究喀纳斯景区生物多样性的可持续管理提供依据。
1 材料与方法

1.2样地设置与调查 样地设置在景区主要景点附近,海拔1 300~1 430 m。样地的群落调查采取距离取样法,即首先按照草地植被状况选择能代表山地草甸群落特征的典型地段,再根据这些地段距离旅游景点的远近选择样地,并将这些地段划分为:1)旅游核心地段(CZ,样地距景点0~100 m);2)旅游外围地段(PZ,样地距景点100~300 m);3)旅游边缘地段(MZ,样地距景点300~500 m);4)未旅游地段(NdZ,样地距景点500~800 m)。每个典型地段均设置3个样地,共12个样地,每块样地的面积均为10 m×10 m(表1)。分别于2006、2007和2008年8月上旬在每块样地内随机设置10个面积为0.5 m×0.5 m的样方,共360个样方。分别记录样方内的植物种类、高度、密度和盖度。在群落生物量最高的时间(8月上旬),齐地剪取样方内的地上部分,分种测定其地上生物量。同时,在每个样地内随机设置5个样点,利用SC-900土壤紧实度仪对每个样点的土壤紧实度进行测定;对每个样点按深度分0-10、10-20和20-30 cm 3个层次采集混合土样。采集的土样经风干、去杂、粉碎过筛后,进行理化性质的分析。土壤容重采用环刀法、土壤含水量采用铝盒烘干法、土壤有机质采用重铬酸钾容量法-外加热法、碱解氮采用碱解扩散法、有效磷采用0.5 mol·L-1NaHCO3浸提钼锑抗比色法、速效钾采用1 mol·L-1NH4OAc浸提-火焰光度计法进行测定,具体步骤参见鲍士旦[28]的方法。
1.3植物多样性测度指标
1.3.1物种多样性层次
生态优势度指数(Ecological Dominance Index,EDI):EDI=∑Pi2.
式中,H为Shannon-Wiener指数,S为样方内所有物种数,Pi为样方内种的重要值,Pi=(相对密度+相对高度+相对盖度+相对生物量)/400。H值较低,表明群落α多样性较低;H值较高,表明群落α多样性较高。而EDI值低,表明群落由若干优势度相近的种群组成,群落环境被较充分利用,物种多样性及维持的群落功能(恢复能力)较高;EDI值高则表明群落仅有少数优势种控制,群落物种多样性及稳定性较低[29-31]。
1.3.2功能群多样性层次
植物功能群依据生活型和草地植物的生态经济类群[32]划分为:①灌木和半灌木(Shrubs and Semi-shrubs,Sh);②多年生丛生禾草(Perennial Bunch Grasses,PbG);③多年生根茎禾草(Perennial Rhizome Grasses,PrG);④多年生莎草(Perennial Sedges,PS);⑤多年生杂类草(Perennial Forbs,PF);⑥多年生豆科植物(Perennial Legumes,PL);⑦一、二年生草本(Annuals and Biennials,AB)。
功能群丰富度(Functional Group Richness,FGR)FGR=1-∑GPi。式中,N为样方内所有功能群种类数[11]。
功能群优势度指数(Group Dominance Index,GDI)GDI=1-∑GPi,式中,GPi为样方内顶级群落优势功能群的重要值[32],在本研究中分别为多年生根茎禾草、多年生杂类草和多年生豆科植物。GDI值低,表明群落由若干顶级群落优势功能群组成,群落处于演替顶级阶段,群落恢复能力较高;GDI值高则表明群落中顶级群落优势功能群由于干扰减少或消失,群落处于逆向演替阶段,群落恢复能力较低[33]。
1.3.3群落多样性层次 多样性主要用以测度沿着环境梯度变化植被群落间物种组成的差异,表现出不同环境梯度群落间共有种越少其多样性值就越高的特征[34]。Wilson and Shmida指数βt=[g(H)+l(H)]/2X,式中,g(H)是沿生境梯度H增加的物种数目;l(H)是沿生境梯度H失去的物种数目,即在上一个梯度中存在而在下一个梯度中没有的物种数目;βt受α多样性的影响较小,其测度结果直观地反映了梯度上群落物种组成的变化[35]。
群落构成指数(Community Structure Index,CSI)GSI=1-∑PiCFV,式中,Pi同上,CFV是各植物种的饲用价值评分值,取值范围为0~1.00;优等牧草饲用价值评分值为0.90~1.00,优良等牧草为0.80~0.89,良等牧草为0.70~0.79,中等牧草为0.60~0.69,劣等牧草为0.01~0.59,有毒有害植物为0[36]。CSI值低表明群落中优良牧草比例高,草地品质高;CSI值高表明群落中劣等牧草、毒杂草比例高,草地品质低。
1.4数据处理 所有数据为3年测定数据的均值。对不同旅游地段的土壤理化性质、植物多样性及生产力等数据采用SPSS for Windows 13.0软件包的One-way ANOVA进行方差分析、计算标准误,并利用LSD对上述指标进行比较。采用SPSS for Windows 13.0软件对土壤理化性质与植物多样性及生产力进行相关分析、曲线拟合分析。采用EXCEL 2007绘制图表。
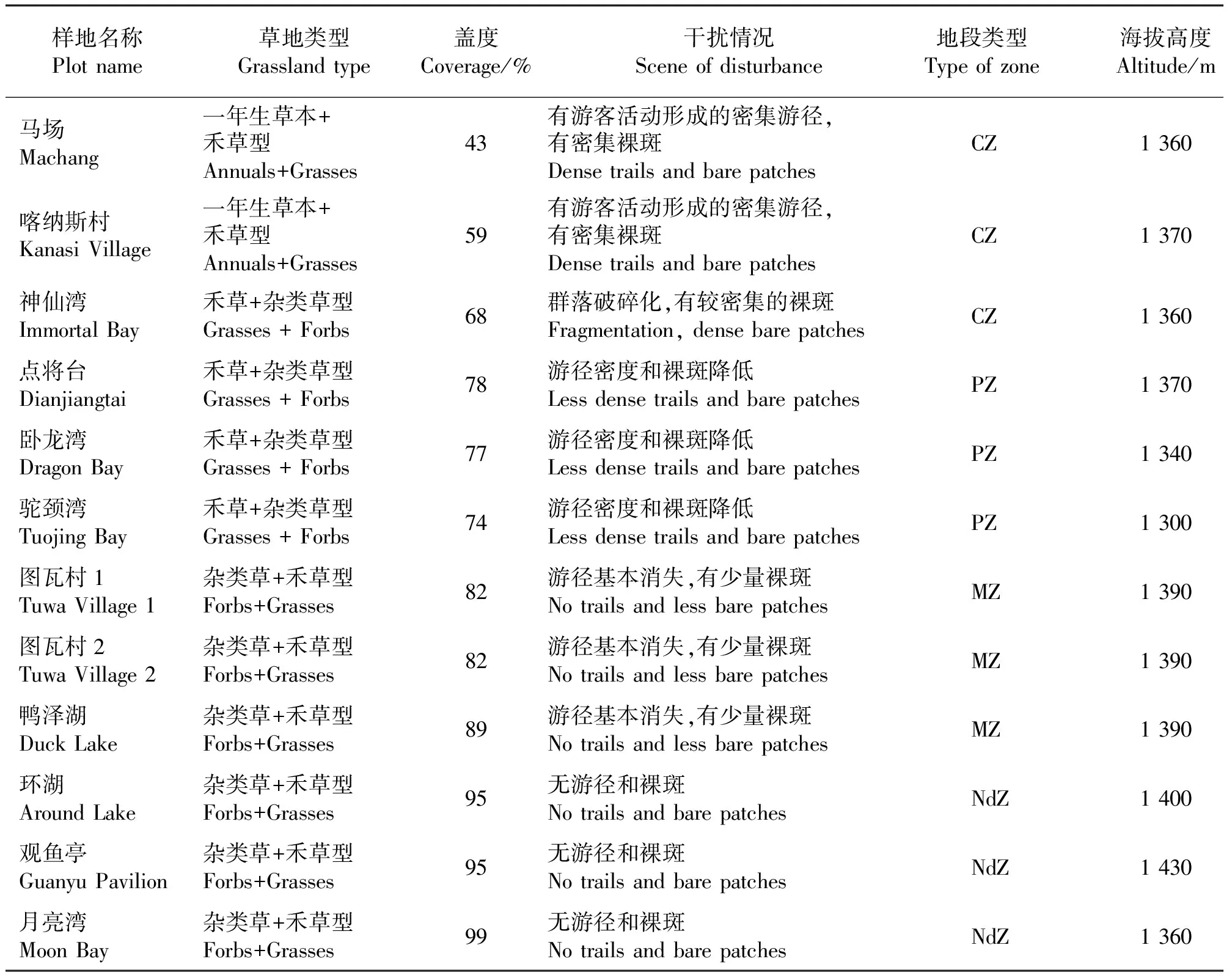
表1 样地类型及其群落特征与干扰情况Table 1 Grasslande types, characteristics and disturbance of grassland communities in this study
2 结果与分析
2.1山地草甸植物多样性及其生产力的变化
2.1.1物种多样性及其生产力的变化 从物种多样性的分布规律看,4个地段共发现96种维管束植物,分属27科78属,其中CZ有26种,分属14科24属;PZ有36种,分属19科31属;MZ有61种,分属24科57属;NdZ有71种,分属21科61属(表2)。样方中出现最多的有禾本科(12种)、菊科(12种)、蔷薇科(10种)、豆科(9种)及莎草科(2种),出现率分别为97.222%、88.889%、73.611%、46.759%和77.315%。不同地段植物物种分布具有明显差异,其中,CZ以蓼科、禾本科、藜科和十字花科植物为主,PZ以菊科、禾本科、莎草科及蓼科植物为主,MZ以莎草科、禾本科、蔷薇科及菊科植物为主,而NdZ则以禾本科、莎草科、蔷薇科、茜草科及豆科植物为主。不同地段群落优势种也发生了变化(表2)。CZ是生长点低、营养繁殖能力强的扁蓄(Polygonumaviculare)与草地早熟禾占统治地位,二者重要值之和大于0.6;PZ则是草甸早熟禾与繁殖能力强的千叶蓍(Achilleamillefolium)占统治地位,二者重要值之和大于0.5;MZ是生长点低与侵占力较强的短柄苔草、绿草莓、紫苞鸢尾占统治地位,但三者重要值之和小于0.5;NdZ则是生长点高、有性繁殖占主导的地榆、白花砧草(Galiumboreale)、狭颖鹅观草、短柱苔草与草地早熟禾占统治地位,但五者重要值之和小于0.5。短柄苔草与草地早熟禾在喀纳斯景区草地群落中分布较广,在NdZ中是群落下层主要优势种;在MZ和PZ中逐渐成为群落上层优势种;而在CZ中其优势地位被扁蓄取代。

表2 不同旅游地段主要植物种分布的变化Table 2 Species distribution among different tourism zones
从物种多样性指数变化规律来看,距离旅游景点越远的地段,拥有较高的H值和较低的EDI值,而距离旅游景点越近的地段,拥有较低的H值和较高的EDI值,且各地段间有显著差异(P<0.05)(图1)。根据群落主要优势种的生产力(生物量)变化规律(表3),可将研究区域植物种分为3种类型:第1种,随着与旅游景点距离的增加,其生物量呈上升趋势,如短柄苔草、地榆、白花砧草、狭颖鹅观草;第2种,随着与旅游景点距离的增加,其生物量呈下降趋势,如扁蓄、灰藜(Chenopodiumalbum);第3种,随着与旅游景点距离的增加,其生物量呈先上升后下降趋势,如绿草莓、千叶蓍、药用蒲公英(Taraxacumofficinale)、岩蒿(Artemisiarupestris)等。由此可见,喀纳斯景区各景点附近草地受人为干扰后,群落物种多样性明显下降,物种组成发生明显改变,群落结构被破坏,其维持的功能也呈下降趋势。
2.1.2功能群多样性及其生产力的变化 从功能群多样性变化规律来看,距离旅游景点越远的地段,拥有越高的FGR值和较低的GDI值,而距离旅游景点越近的地段,拥有越低的FGR值和较高的GDI值,除MZ与NdZ的FGR无显著差异外,其它地段之间均有显著差异(P<0.05)(图2)。这与物种多样性变化规律相似。从功能群生产力(生物量)变化规律来看(表3),PS、PF和PL随着与旅游景点距离的增加其生物量呈上升趋势,AB则呈下降趋势,PrG呈先下降再上升趋势,而PbG在NdZ和PZ出现两个峰值。从地段来看,CZ以AB占主导地位,PZ以PF和PbG占主导地位,MZ以PF和PS占主导地位,NdZ以PF、PS和PbG占主导地位。
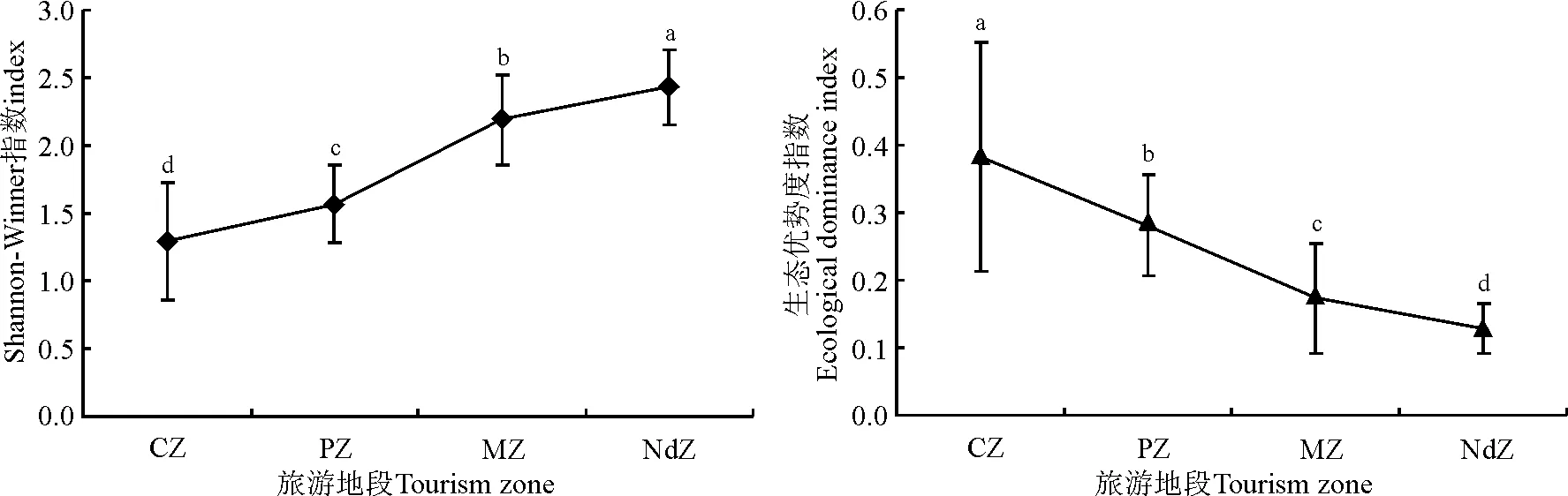
图1 不同旅游地段物种多样性的变化Fig.1 Species diversity of different tourism zones
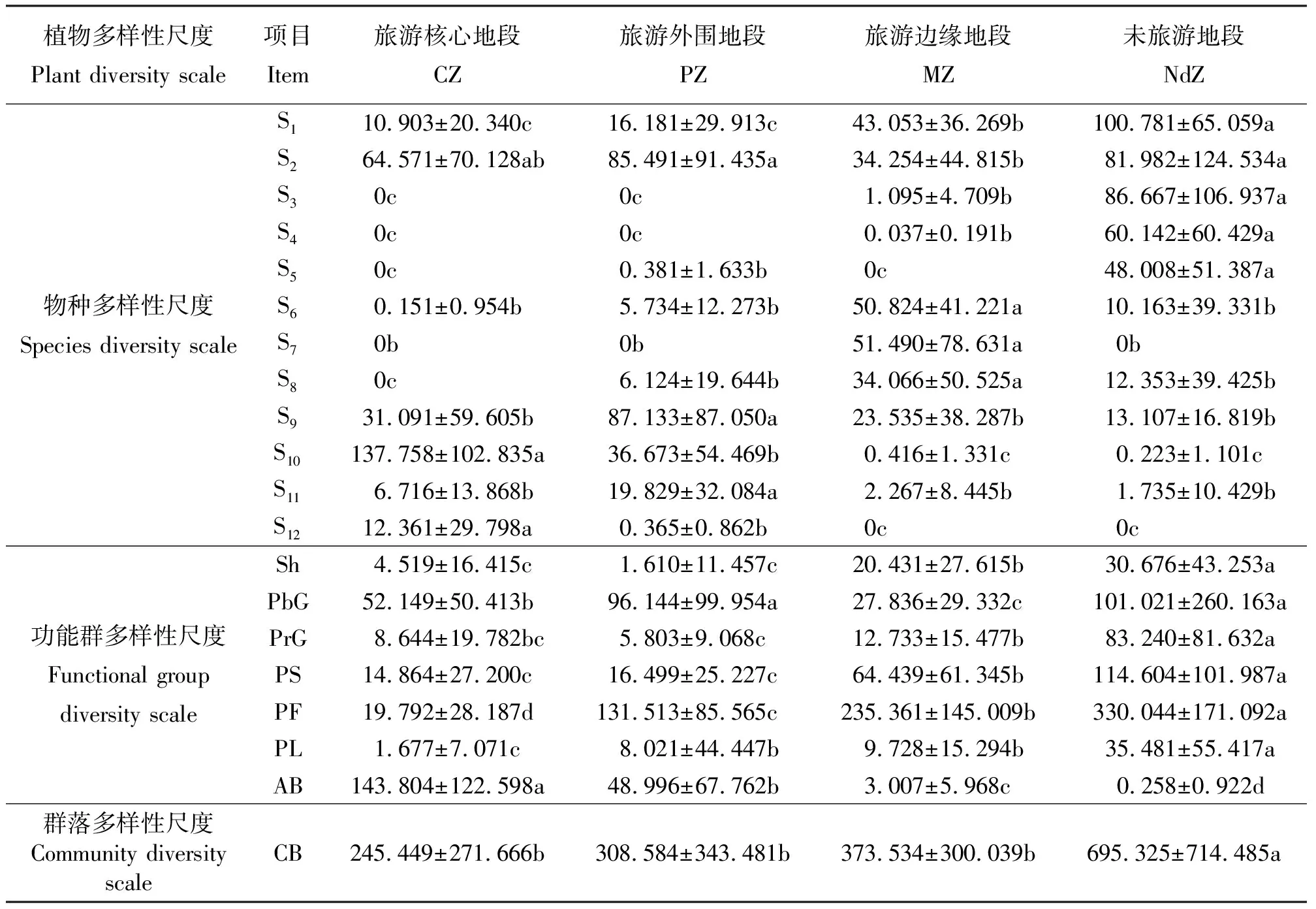
表3 不同旅游地段物种、功能群及群落多样性尺度的生物量变化Table 3 Biomass of species, functional group and community of different tourism zones
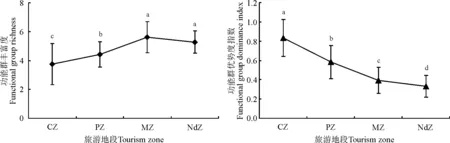
图2 不同旅游地段功能群多样性的变化Fig.2 Functional group diversity of different tourism zones
2.1.3群落多样性及其生产力的变化 Wilson and Shmida指数在CZ-PZ的值较低,PZ-MZ的值较高,而在MZ-NdZ的值最高(图3)。这表明CZ与PZ群落间共有种较多,而NdZ与MZ群落间共有种较少。CSI反映了草地利用率或者草地品质[33,37]。距离旅游景点越远的地段,拥有较低的CSI值,而距离旅游景点越近的地段,拥有较高的CSI值,且CZ显著高于其它地段(P<0.05)。从群落生产力(生物量)变化规律来看(表3),随着远离旅游景点其生物量呈上升趋势,且NdZ与其它地段有显著差异。
2.2植物多样性及其生产力与土壤理化性质的关系
2.2.1物种多样性及其生产力与土壤理化性质的关系 不同地段0-30 cm土壤理化性质测定结果表明(表4),随着与旅游景点距离的增加,土壤含水量呈上升趋势,土壤容重、紧实度及速效钾呈下降趋势;NdZ的土壤有机质和碱解氮显著高于其它区,而有效磷则是CZ显著高于其它区(P<0.05)。物种多样性尺度各指标和土壤理化性质的相关分析结果表明(表5),H与土壤含水量呈极显著正相关(P<0.01),与土壤有机质呈显著的正相关,与土壤容重、紧实度、速效钾及有效磷呈极显著负相关,与土壤碱解氮相关不显著。而EDI与H正好相反。曲线拟合可知,H和EDI与土壤含水量、紧实度和速效钾可拟合为二项式,EDI与土壤容重可拟合为指数函数。由此可见,物种多样性与土壤理化性质并不是简单的正相关或负相关关系[37]。
从群落主要优势种的生产力(生物量)与土壤理化性质的相关系数来看(表6),主要优势种可被分为3种类型:第1种与土壤含水量、有机质呈显著正相关(P<0.05)或极显著正相关(P<0.01),与土壤容重、紧实度、速效钾及有效磷呈极显著或显著负相关,与土壤碱解氮呈正相关,如S1、S3、S4和S5;第2种与土壤含水量呈极显著负相关,与土壤容重、紧实度、速效钾及有效磷呈显著或极显著正相关,与土壤有机质、碱解氮呈极显著负相关或负相关,如S10和S12;第3种与大部分土壤理化指标相关不显著(P>0.05),如S2、S7、S8、S9和S11等。
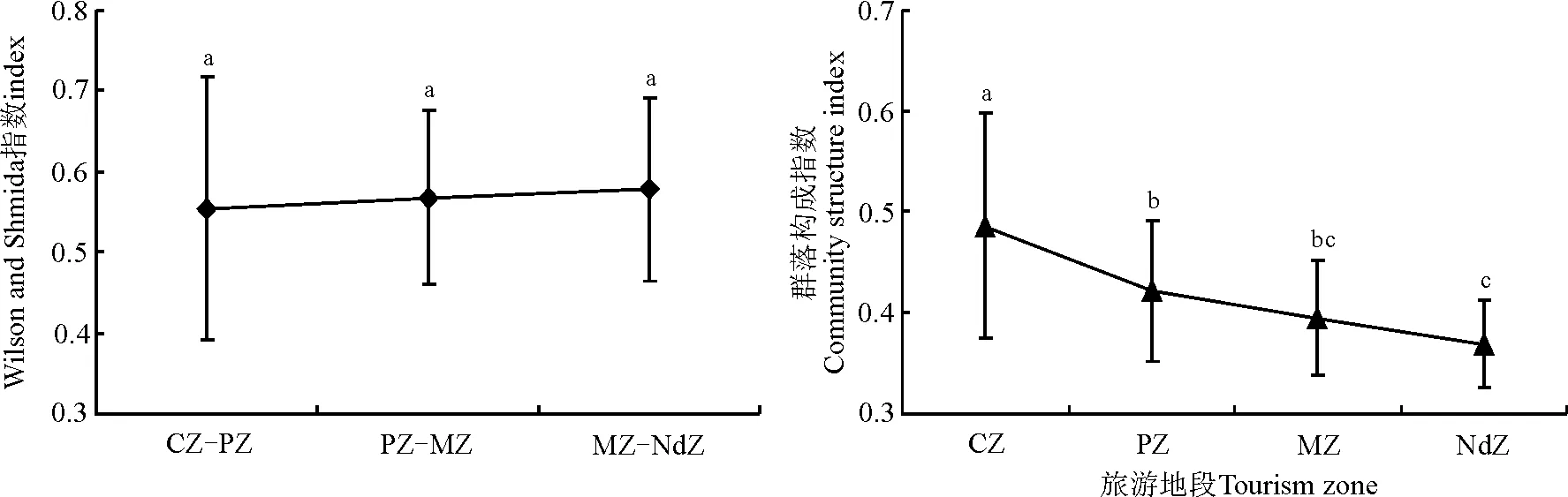
图3 不同旅游地段群落多样性的变化Fig.3 Community diversity of different tourism zones
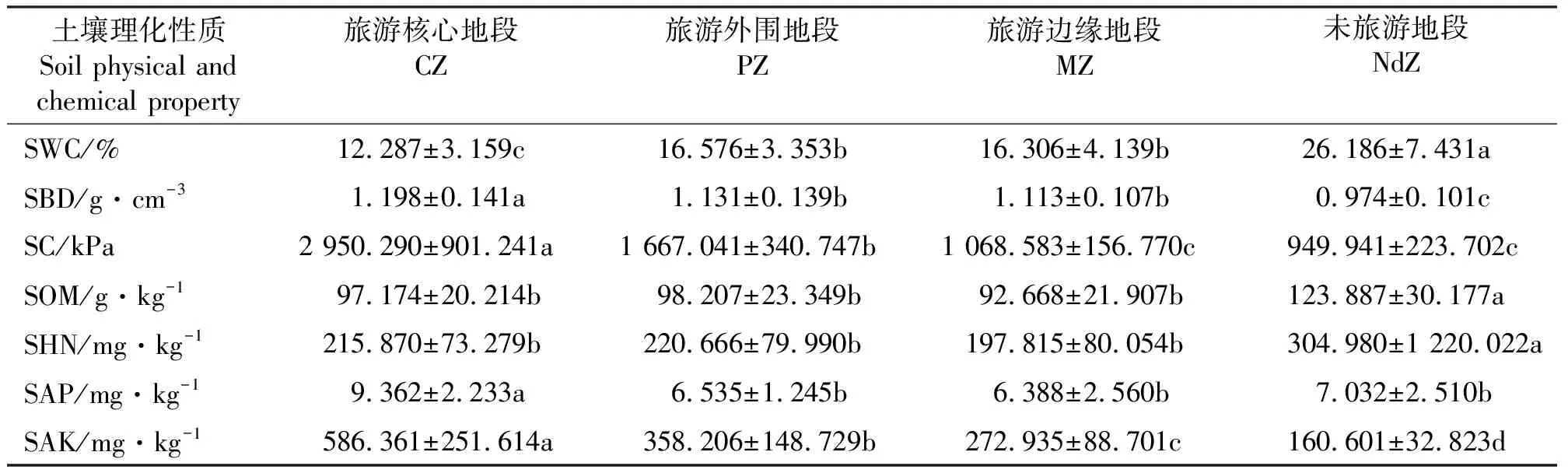
表4 不同旅游地段0-30 cm土壤理化性质Table 4 Soil physical and chemical properties of soil depth 0-30 cm among different tourism zones
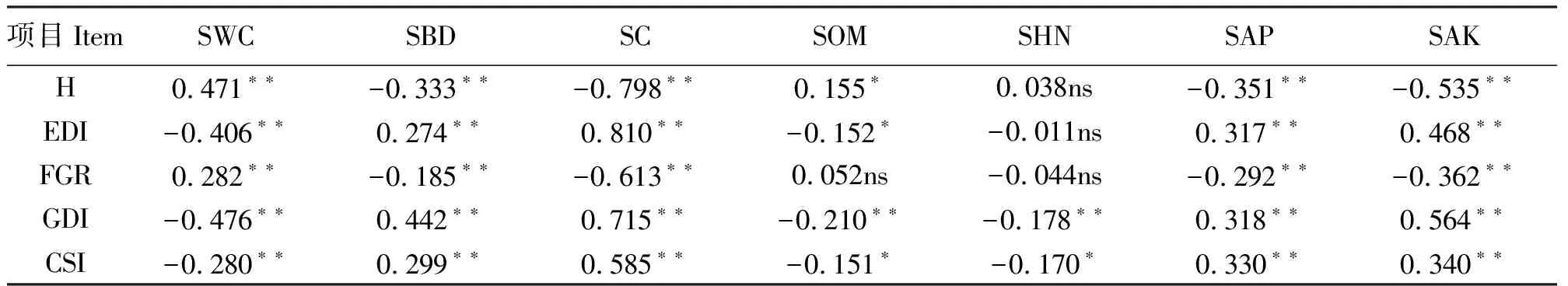
表5 植物多样性指数与土壤理化性质的相关系数Table 5 Correlation coefficient among plant diversity indexes and soil physical and chemical properties
2.2.2功能群多样性及其生产力与土壤理化性质的关系 FGR与土壤含水量呈极显著正相关(P<0.01),与土壤容重、紧实度、速效钾及有效磷呈极显著负相关,与土壤有机质、碱解氮相关不显著(P>0.05)(表5)。而GDI与土壤含水量、有机质、碱解氮呈极显著负相关,与土壤容重、紧实度、速效钾及有效磷呈极显著正相关。曲线拟合得出(表7),FGR与土壤含水量可拟合为幂函数,与有效磷呈极显著线性负相关;GDI与土壤含水量、容重、速效钾及有效磷均可拟合为二项式,与土壤紧实度可拟合为对数函数。
土壤含水量与Sh、PrG、PS、PF及PL呈极显著正相关(P<0.01),与PbG呈显著正相关(P<0.05),与AB呈极显著负相关(表6)。土壤容重、紧实度、速效钾及有效磷与Sh、PrG、PS、PF呈极显著负相关,与AB呈极显著正相关;土壤紧实度与PbG呈极显著正相关,而土壤容重、碱解氮、速效钾及有效磷与PbG相关性不显著(P>0.05);土壤速效钾与PL呈极显著负相关,土壤容重、紧实度、碱解氮及有效磷与PL相关不显著。土壤有机质与PbG、PrG、PF及PL呈极显著正相关或显著正相关,与AB呈极显著负相关,与Sh、PS相关性不显著。碱解氮仅与PF及AB呈显著负相关或极显著负相关。
2.2.3群落多样性及其生产力与土壤理化性质的关系 CSI与土壤含水量呈极显著负相关(P<0.01),与土壤有机质、碱解氮呈显著负相关(P<0.05),与土壤容重、紧实度、速效钾及有效磷呈极显著正相关(表5)。群落生物量与土壤含水量、有机质呈极显著正相关,与土壤容重、紧实度、速效钾呈极显著负相关,与土壤碱解氮及有效磷相关不显著(表6)。曲线拟合分析得出(表7),CSI与土壤含水量、紧实度、速效钾及有效磷均可拟合为二项式,与土壤容重可拟合为指数函数(P<0.01)。群落生产力(生物量)与土壤含水量呈显著线性正相关,与土壤速效钾可拟合为二项式。
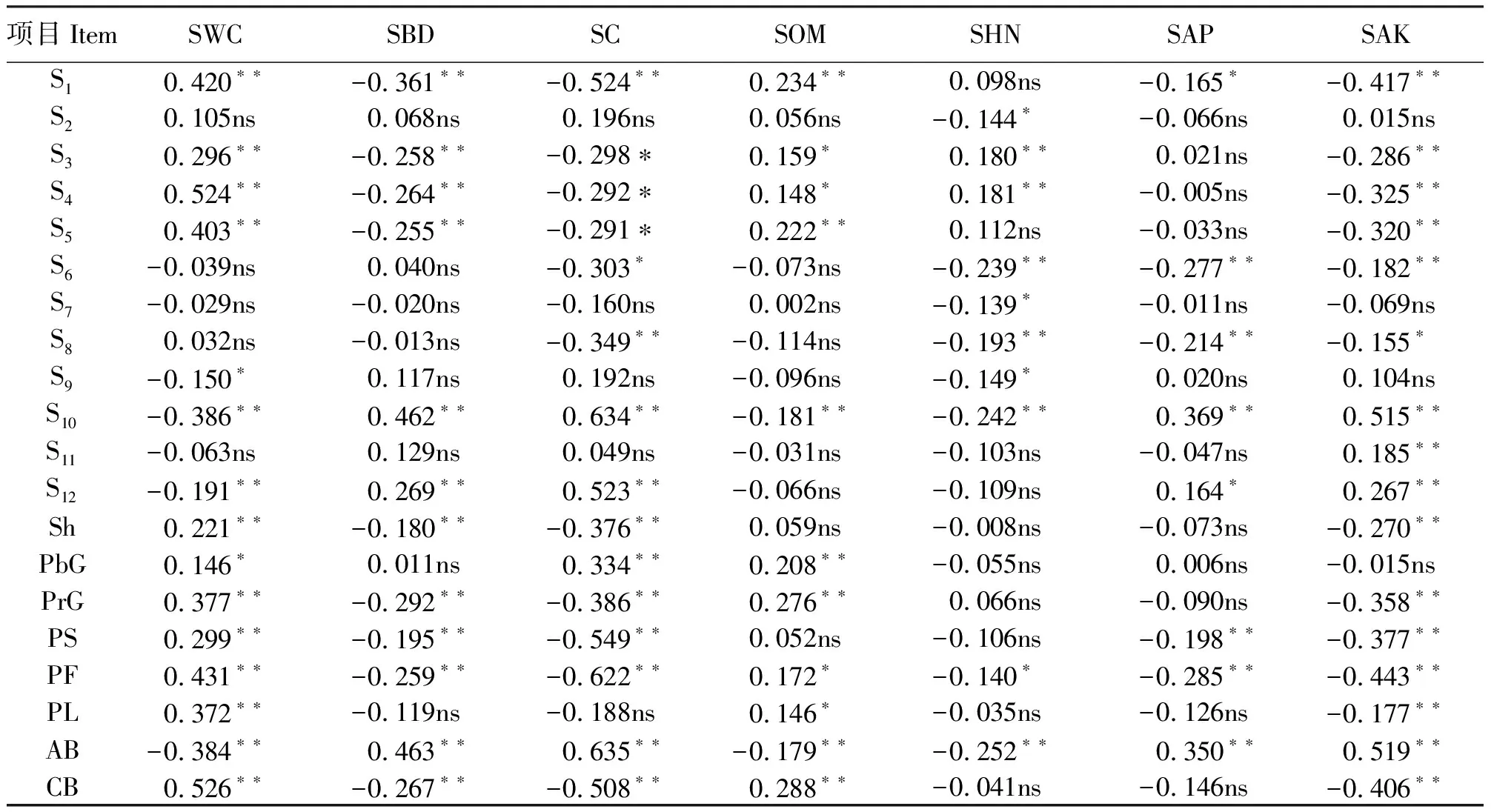
表6 物种、功能群及群落各尺度生物量与土壤理化性质的相关系数Table 6 Correlation coefficient among biomass of different scales and soil physical and chemical properties
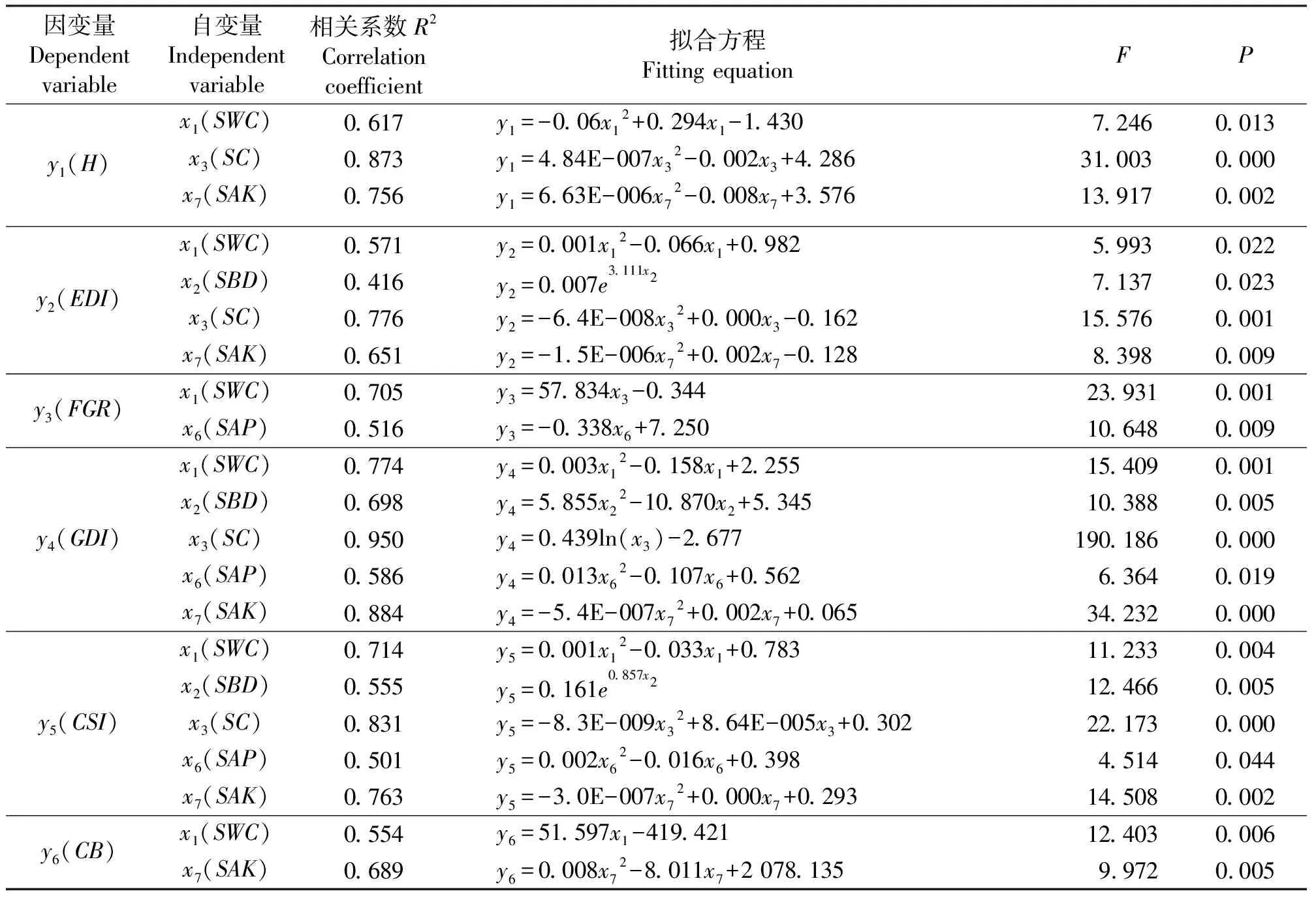
表7 土壤理化性质与植物多样性指数及群落生物量的曲线拟合Table 7 Curve estimation between plant diversity indexes, community biomass and soil physical and chemical properties
3 讨论与结论
3.1旅游活动对不同尺度植物多样性及生产力的影响 在物种多样性尺度,旅游活动的增加往往伴随着伴人植物侵入,原有顶级群落的优势种地位和作用发生转变[38]。在喀纳斯景区,旅游活动首先使物种多样性的分布特征发生改变,距离旅游景点越近,属、种的数量显著减少。其次,山地草甸植物种类组成发生改变,在旅游景点附近,扁蓄、灰藜等伴人植物的作用和地位逐渐加强,甚至成为优势种;而山地草甸原有的短柄苔草、地榆、狭颖鹅观草、白花砧草等优势种逐渐变为伴生种甚至消失。另外,山地草甸群落结构也因种类组成的变化而改变,在旅游活动的干扰下,纤细、高大的上层植物消失,逐渐被低矮、营养繁殖能力强、位于群落下层的短柄苔草、草地早熟禾所取代,群落结构简化。从物种多样性指数及主要物种生产力(生物量)来分析,距离旅游景点越近,α多样性越低,群落被少数生长点低、营养器官柔软具弹性、营养繁殖能力强的物种所控制[39],物种多样性及群落稳定性降低。
在功能群多样性尺度,功能群多样性指数的变化规律与物种多样性指数相似,但出现了功能群丰富度在旅游边缘地段高于未旅游地段的情形。Walker[40]和Naeem[41]认为这是功能群具有的功能冗余对外界干扰的缓冲。从不同功能群生物量的变化规律来看,不同功能群对旅游活动干扰的敏感性不同,多年生莎草、多年生豆科植物和多年生根茎禾草受干扰的影响较大,而一、二年生草本适宜在频繁干扰的生境中生存,并且这种功能群结构的变化对群落功能也产生了明显影响。在群落多样性尺度, 多样性变化不明显,而草地品质、群落生物量随旅游活动的增加呈下降趋势,但下降趋势较前两个尺度有所减缓,这表明生态系统内的中小尺度干扰可以被大尺度下的系统有所消化[32]。
3.2旅游活动影响下不同尺度植物多样性及其生产力与环境因子的关系 植被与环境的关系是生态学和环境科学研究的热点问题和重要内容之一[42-43]。土壤作为植物生长的重要物质基础,其理化性质的不同,可能影响植物的生长,使包括植物多样性、生产力在内的群落特征发生变化[26,34]。喀纳斯景区不同旅游地段0-30 cm土壤理化性质分析结果表明,土壤含水量、有机质和碱解氮含量越高,土壤容重、紧实度及速效钾越低,有利于植被生长,有效磷的影响则不明显。在物种多样性尺度,短柄苔草、地榆、狭颖鹅观草、白花砧草等山地草甸原有的优势种受土壤理化性质的影响较大,土壤理化性质变劣(土壤容重、紧实度及速效钾含量升高,土壤含水量、有机质含量下降),不利于植物生长;草地早熟禾、千叶蓍、药用蒲公英等物种受土壤理化性质的影响较小;而扁蓄、灰藜则受益于这种变化。在功能群多样性尺度,土壤理化性质变劣,灌木和半灌木、多年生根茎禾草、多年生莎草、多年生豆科牧草及多年生杂类草生长受到抑制,多年生丛生禾草受到的影响较小,而一、二年生草本则有利于其生长。因此,不同植物功能群对环境变化的敏感性不同[44]。在群落多样性尺度,土壤理化性质变劣,也使群落生物量与草地品质下降。通过曲线拟合分析可知,3个尺度的植物多样指数大部分可与土壤理化性质拟合为二项式,表现出低的植物多样性出现在土壤物理性质及速效养分梯度的中间位置,土壤有机质、碱解氮则对植物多样性指数的影响不显著[26]。群落生产力可与土壤含水量拟合为线性正相关,与有效磷则可拟合为二项式。综合来看,喀纳斯景区山地草甸土壤含水量、有机质高,土壤容重、紧实度与土壤速效钾含量低,而土壤碱解氮与有效磷含量适中,会使3个尺度的植物多样性及生产力提高。因此,外界的旅游干扰因素和土壤因素均可影响物种、功能群及群落3个尺度的植物多样性分布格局及生产力水平。但是如何分离干扰因素与环境因素对不同尺度植物多样性的作用效应,是下一步研究工作的重点及难点。
[1] White P S.Pattern,process and natural disturbance in vegetation[J].Botanical Review,1979,45:229-299.
[2] McGrady-Steed J,Harris P M,Morin P.Biodiversity regulates ecosystem predictability[J].Nature,1997,390:162-164.
[3] McCann K,Hastings A,Huxel G.Weak trophic interactions and the balance of nature[J].Nature,1999,385:794-798.
[4] Hector A,Beale A J,Minns A.Consequences of the reduction of plant diversity for litter decomposition:Effects through litter quality and microenvironment[J].Oikos,2000,90(2):357-371.
[5] Loreau M,Naeem S,Inchausti P,etal.Biodiversity and ecosystem function:Current knowledge and future challenges[J].Science,2001,294:804-808.
[6] Wardle D,Zackrisson O.Effects of species and functional group loss on island ecosystem properties[J].Nature,2005,435:806-810.
[7] Díaz S,Lavorel S,de Bello F,etal.Incorporating plant functional diversity effects in ecosystem service assessments[J].PNAS,2007,104(52):20684-20689.
[8] Schulze E D,Mooney H A.Biodiversity and Ecosystem Function[M].New York:Springer-Verlag,1993.
[9] Spehn E M,Joshi J,Schmid B,etal.Aboveground resource use increases with plant species richness in experimental grassland ecosystems[J].Functional Ecology,2000,14(3):326-337.
[10] Tilman D,Knops J,Wedin D,etal.The influence of functional diversity and domposition on ecosystem rocesses[J].Science,1997,277:1300-1302.
[11] Lanta V,LepšJ.Effect of functional group richness and species richness in manipulated productivity-diversity studies:A glasshouse pot experiment[J].Acta Oecologica,2006,29:85-96.
[12] Tilman D,Reich P B,Knops J.Biodiversity and ecosystem stability in a decade long grassland experiment[J].Nature,2006,441:629-633.
[13] Bai Y F,Han X G,Wu J G,etal.Ecosystem stability and compensatory effects in the Inner Mongolia grassland[J].Nature,2004,431:181-184.
[14] Hughes A R,Byrnes J E,Kimbro D L,etal.Reciprocal relationships and potential feedbacks between biodiversity and disturbance[J].Ecology Letters,2007,10:849-864.
[15] Vitousek P M,Matson P A,Van Cleve K.Nitrogen availability and nitrification during succession,primary,secondary and old field series[J].Plant and Soil,1989,115:229-239.
[16] Dybzinski R,Fargione J E,Zak D R,etal.Soil fertility increases with plant species diversity in a long-term biodiversity experiment[J].Oecologia,2008,158:85-93.
[17] Reich P B,Knops J,Tilman D,etal.Plant diversity enhances ecosystem responses to elevated CO2and nitrogen deposition[J].Nature,2001,410:809-812.
[18] Dijkstra F A,Hobbie S E,Reich P B,etal.Divergent effects of elevated CO2,N fertilization,and plant diversity on soil C and N dynamics in a grassland field experiment[J].Plant and Soil,2005,272:41-52.
[19] Ewel J J.Designing agricultural ecosystems for the humid tropics[J].Annual Review on Ecological System,1986,17:245-254.
[20] Ewel J J,Mazzarino M J,Berish C W.Tropical soil fertility changes under monoculture and successional communities of different structure[J].Ecological Applications,1991,1:289-302.
[21] William J R,Renard K G,Dyke P T.EPIC:A new method for assessing erosion,effect on soil productivity[J].Journal of Soil and Water Conservation,1983,36:381-383.
[22] Tilman D,Wedin D,Knops J.Productivity and sustainability influenced by biodiversity in grassland ecosystems[J].Nature,1996,379:718-720.
[23] 王震洪,段昌群,侯永平,等.植物多样性与生态系统土壤保持功能关系及其生态学意义[J].植物生态学报,2006,30(3):392-403.
[24] 郑淑华,赵萌莉,韩国栋,等.不同放牧压力下典型草原土壤物理性质与植被关系的研究[J].干旱区资源与环境,2005,19(7):199-203.
[25] 顾峰雪,张远东,潘晓玲,等.阜康绿洲土壤盐渍化与植物群落多样性的相关性分析[J].资源科学,2002,24(3):42-48.
[26] 左小安,赵学勇,赵哈林,等.科尔沁沙质草地群落物种多样性、生产力与土壤特性的关系[J].环境科学,2007,28(5):945-951.
[27] 郑伟,朱进忠,潘存德.喀纳斯草地群落和土壤理化特征对放牧干扰的响应[J].草业科学,2008,25(8):103-108.
[28] 鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000:23-35.
[29] 吴甘霖,黄敏毅,段仁燕,等.不同强度旅游干扰对黄山松群落物种多样性的影响[J].生态学报,2006,26(12):3924-3930.
[30] 许鹏.新疆草地资源及其利用[M].乌鲁木齐:新疆科技卫生出版社,1993:51-79.
[31] 高伟,鲍雅静,李政海,等.不同保护和利用方式下羊草草原群落生物量及能量功能群构成的比较[J].干旱区资源与环境,2010,24(12):132-136.
[32] 郑伟,朱进忠,潘存德.草地植物多样性对人类干扰的多尺度响应[J].草业科学,2009,26(8):72-80.
[33] 张大勇,王刚,杜国祯.甘南山地草原人工草场的演替[J].植物生态学与地植物学学报,1990,14:103-109.
[34] 马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究Ⅱ.丰富度、均匀度和物种多样性指数[J].生态学报,1995,15(3):268-277.
[35] 陈生云,刘文杰,叶柏生,等.疏勒河上游地区植被物种多样性和生物量及其与环境因子的关系[J].草业学报,2011,20(3):70-83.
[36] 沈振西,陈佐忠,周兴民,等.高施氮量对高寒矮嵩草甸主要类群和多样性及质量的影响[J].草地学报,2002,10(1):7-17.
[37] 张桂萍,张峰,茹文明.旅游干扰对历山亚高山草甸优势种群种间相关性的影响[J].生态学报,2005,25(11):2868-2874.
[38] 高贤明,马克平,黄建辉,等.北京东灵山地区植物群落多样性的研究Ⅺ.山地草甸多样性[J].生态学报,1998,18(1):24-32.
[39] 鲁庆彬,游卫云,赵昌杰,等.旅游干扰对青山湖风景区植物多样性的影响[J].应用生态学报,2011,22(2):295-302.
[40] Walker B H.Conserving biological diversity through ecosystem resilience[J]. Conservation Biology,1995,9:747-752.
[41] Naeem S.Species redundancy and ecosystem reliability[J].Conservation Biology,1998,12:39-45.
[42] Schaffers A P.Soil,biomass and management of semi-natural vegetation part Ⅱ.Factors controlling species diversity[J].Plant Ecology,2002,158:247-268.
[43] Kahmen A,Perner J,Audorff V,etal.Effects of plant diversity,community composition and environmental parameters on productivity in montane European grasslands[J].Oecologia,2005,142:606-615.
[44] 李禄军,曾德慧,于占源,等.氮素添加对科尔沁沙质草地物种多样性和生产力的影响[J].应用生态学报,2009,20(8):1838-1844.
猜你喜欢
竹子学报(2019年1期)2019-09-12
中国水土保持科学(2019年6期)2019-04-26
现代园艺(2017年21期)2018-01-03
农业机械学报(2017年4期)2017-06-05
红土地(2016年3期)2017-01-15
绿色科技(2016年16期)2016-10-11
环球市场信息导报(2015年15期)2015-12-03
中国水土保持(2015年2期)2015-03-22
海峡姐妹(2015年9期)2015-02-27
亚热带资源与环境学报(2015年1期)2015-01-22
