
马铃薯块茎蛾幼虫对不同寄主植物的取食选择性
2013-04-13 07:43王浩元张立敏陈斌李正跃
中国马铃薯 2013年4期
王浩元,张立敏,陈斌*,李正跃*
(1.云南农业大学植物保护学院,农业生物多样性与病虫害控制教育部重点实验室,农业生物多样性应用技术国家工程研究中心,云南昆明650201;2.云南农业大学基础与信息工程学院,云南昆明650201)
病虫防治
马铃薯块茎蛾幼虫对不同寄主植物的取食选择性
王浩元1,张立敏2,陈斌1*,李正跃1*
(1.云南农业大学植物保护学院,农业生物多样性与病虫害控制教育部重点实验室,农业生物多样性应用技术国家工程研究中心,云南昆明650201;2.云南农业大学基础与信息工程学院,云南昆明650201)
采用叶盘法,室内测定了马铃薯块茎蛾[Phthorimaes operculella(Zeller)]初孵幼虫和3龄幼虫对马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆植物的选择性与非择性取食。结果表明,马铃薯块茎蛾初孵幼虫和3龄幼虫取食对供试植物具有明显的选择性,随着时间的延续,选择性逐渐降低。测定的1~48 h内马铃薯块茎蛾初孵幼虫和3龄幼虫对马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆的总体选择取食率间差异显著(初孵幼虫:F=60.94,P<0.01;3龄幼虫:F=66.34,P<0.01),对供试植物的总体选择顺序为马铃薯>烟草>番茄>酸浆>辣椒>龙葵>茄子>蓖麻>曼陀罗。马铃薯块茎蛾初孵幼虫和3龄幼虫对受其危害后的马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆叶片也现出明显的取食选择性,该选择性随着处理时间的延长而逐渐降低,处理后第48 h,初孵幼虫和3龄幼虫对供试验植物的选择性无显著差异(初孵幼虫:F= 2.34,P>0.05;3龄幼虫:F=1.76,P>0.05),在1~48 h内对受害植物的选择取食顺序为马铃薯>番茄>烟草>酸浆>龙葵>辣椒>茄子>蓖麻>曼陀罗。该研究结果将为利用植物多样性生态控制马铃薯块茎蛾提供参考。
马铃薯块茎蛾;幼虫;取食选择性;取食率
马铃薯块茎蛾[Phthorimaes operculella(Zeller)]隶属鳞翅目(Lepidotera),双孔亚目(Ditrysia),麦蛾总科(Gelechioidea),麦蛾科(Gelechiida),块茎蛾属(Phthirimaea),又叫马铃薯麦蛾,烟草潜叶蛾,是田间与储藏期马铃薯的主要害虫[1,2],田间危害可引起减产20%~30%,在马铃薯储藏期中危害率可达100%[3]。同时该虫还是烟草的主要害虫[4]。马铃薯作为重要的粮食、蔬菜、饲料和加工原料,被云南省列为重要产业。云南地理位置和气候特殊,栽培品种及栽培制度也不尽相同,这种生态环境,为马铃薯块茎蛾提供了适宜的生态条件和良好的食料条件,因而该虫是云南马铃薯上的主要害虫。
有些昆虫的幼虫对不同寄主植物的取食选择性不同[5],据报道,马铃薯块茎蛾可危害马铃薯、茄子、番茄、辣椒、曼陀罗、枸杞、龙葵、酸浆、刺蓟、颠茄、洋金花等茄科植物,目前,有关马铃薯块茎蛾成虫对不同寄主植物的产卵选择行为研究已有一些研究报道[6-12],而对于其幼虫取食习性及行为尚未见研究报道。
1 材料与方法
1.1 试验材料
马铃薯块茎蛾[Phthorimaes operculella(Zeller)]为室内饲养繁殖获得的种群。
供试植物:马铃薯(Solanum tuberosum L.)、烟草(Nicotiana tabacum)、番茄(Lycopersicon esculentum)、辣椒(Capsicum annuum)、茄子(Solanum melongena L.)、曼陀罗(Datura stramonium L.)、龙葵(Solanum nigrum L.)、蓖麻(Lonicera japonica Thunb)、酸浆(Physalis alkekengi)植物叶片。其中马铃薯品种为‘合作88’,番茄品种为‘拉比’,辣椒品种为‘云椒2号’,茄子品种为‘京茄1号’。
1.2 试验方法
1.2.1 非选择性试验
采用王冬妍等[13]培养皿叶盘非选择性试验方法,在直径15 cm的塑料培养皿底衬1张湿润滤纸,沿四周等间距分别放入9片供试植物长约2 cm,宽约1 cm的心叶,在滤纸中央分别接入饥饿24 h的20头初孵幼虫、3龄幼虫。接入虫后即盖上用1层黑棉布和1层红棉布制成的遮光布,移入(25±1)℃、RH 70%±10%、16 L:8 D的光照培养箱,重复10次。接虫后第1,2,4,6,8,10,12,24,36和 48 h检查取食不同植物叶片的幼虫数量,计算取食幼虫数量占处理虫量的百分率,用取食幼虫率表示。
1.2.2 选择性试验
采用王冬妍等[13]培养皿叶盘选择性试验方法,在直径15 cm的塑料培养皿底衬1张湿润的滤纸,沿四周等间距依次放入9种植物心叶各1片,在滤纸中央分别接入饥饿24 h的初孵幼虫、3龄幼虫20头。接入虫后即盖上用1层黑棉布和1层红棉布制成的遮光布,移入(25±1)℃、RH(70±10)%、16 L:8 D的光照培养箱,试验重复10次。按1.2.1观察记录方法检查取食不同植物叶片的幼虫数量,计算取食幼虫率。
1.2.3 对受虫害叶取食的选择性
按照1.2.1方法,在培养皿四周依次放置经马铃薯块茎蛾幼虫取食危害后的9种植物心叶,滤纸中央接入饥饿24 h的3龄幼虫20头。接虫后即盖上遮光布,移入(25±1)℃、RH(70±10)%、16 L:8 D光照培养箱,重复10次。数据观察记录方法同1.2.1。
1.3 数据统计分析
取食选择性数据均按完全随机设计的单因素试验统计分析中邓肯氏新复极差法进行比较各处理间的差异,所有统计分析均通过DPS9.5统计软件[14]完成。
2 结果与分析
2.1 马铃薯块茎蛾的选择性取食
马铃薯块茎蛾初孵幼虫对马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆叶片的选择性取食结果如图1。由图1可知,马铃薯块茎蛾初孵幼虫对不同植物叶片的取食选择性具有明显差异,且其选择性随着处理时间的延长而变化。如处理后1,2,4和6 h,仅对马铃薯、烟草、番茄叶表现出取食选择性。之后,随着处理时间的延长,选择性发生变化,到第48 h时,对马铃薯的选择性最高,取食率为(20.33±3.65)%,而对烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻和酸浆的取食率无显著差异(F=2.54,P>0.05),分别为(15.97±2.33)%、(4.33±0.27)%、(12.67±2.25)%、(8.40±2.28)%、(10.15±2.47)%、(9.73±3.26)%、(10.17±2.45)%和(8.25±2.45)%。综合1~48 h内马铃薯块茎蛾初孵幼虫对供试植物的取食选择情况,表现为对马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆的总体平均取食率分别为(68.66±23.10)%、(7.63±5.58)%、(7.43±3.39)%、(2.66±5.62)%、(2.53± 4.12)%、(1.02±3.21)%、(2.62±4.48)%、(2.04± 4.29)%、(5.42±4.74)%,各处理间差异显著(F= 60.94,P<0.01),总体选择顺序为马铃薯>烟草>番茄>酸浆>辣椒>龙葵>茄子>蓖麻>曼陀罗。
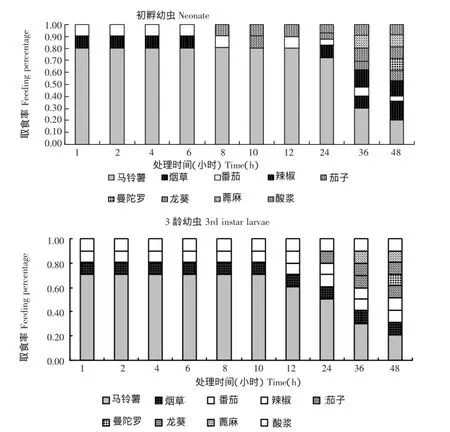
图1 马铃薯块茎蛾初孵幼虫和3龄幼虫对不同寄主植物的取食率(选择性试验,n=20)Figure 1 Feeding percent of neonates and 3rd instar larvae of Phthorimaea operculella on different plant leaves(choice test,n=20)
马铃薯块茎蛾3龄幼虫对马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆叶具有明显的取食选择性,并随着处理时间的延长而发生变化(图1)。处理后1,2,4,6,8,10和12 h,仅对马铃薯、烟草、番茄和酸浆叶具有选择性;从12 h开始,寄主选择性逐渐分化,对马铃薯叶片的选择性逐渐降低,对其他植物的选择性增加。到48 h时,对马铃薯的选择性最高,取食率为(20.67± 2.35)%,而对烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆叶上取食率分别为(10.46± 0.32)%、(10.17±0.48)%、(9.67±0.24)%、(10.35± 0.65)%、(9.67±0.54)%、(9.67±1.67)%、(9.67± 1.45)%、(9.67±1.26)%,各处理间无显著差异(F= 2.35,P>0.05)。综合1~48 h内马铃薯块茎蛾3龄幼虫对供试马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆的总体平均取食率分别表现为58.11%、10.03%、10.00%、3.97%、2.04%、0.97%、2.97%、1.97%、9.95%,各处理间差异显著(F=66.34,P<0.01),总体选择顺序为马铃薯>烟草>番茄>酸浆>辣椒>龙葵>茄子>蓖麻>曼陀罗。
2.2 马铃薯块茎蛾的非选择性取食

图2 马铃薯块茎蛾初孵幼虫和3龄幼虫对不同寄主植物的取食率(非选择性试验,n=20)Figure 2 Feeding percent of neonates and 3rd instar larvae of Phthorimaea operculella on different plant leaves(No-choice test,n=20)
在非选择性试验中,马铃薯块茎蛾初孵幼虫对供试马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆植物表现出明显的选择性(图2),其取食选择性随时间的延长呈下降趋势,处理后1 h时,对马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆叶片的取食率分别为(96.30±1. 23)%、(86.50±3.34)%、(80.00±4.54)%、(80.00± 0.87)%、(80.00±0.43)%、(70.00±0.54)%、(80.00± 0.43)%、(75.70±0.78)%和(80.00±0.35)%,差异显著(F=56.38,P<0.01)。处理2 h时幼虫对马铃薯、烟草、番茄取食率增加至(98.70±1.15)%、(90.00± 3.43)%和(84.00±3.6)%,而对辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆的取食率则降为(78.81±4.65)%、(77.30±3.32)%、(60.80±0.78)%、(75.20±0.67)%、(69.11±0.76)%和(80.04±1.76)%。之后随时间的延长呈缓慢下降,到48 h时,马铃薯块茎蛾初孵幼虫对供试的马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆叶片的取食率降至(90.60± 5.66)%、(73.60±4.65)%、(33.00±5.46)%、(16.00± 3.43)%、(4.00±3.33)%、(0.00±3.65)%、(7.70± 3.67)%、0和(24.70±2.67)%,差异显著(F=123.45,P<0.01)。由此表明,在处理后连续观察的48 h时内,马铃薯块茎蛾初孵幼虫对马铃薯的趋性最强,对烟草、番茄、茄子、蓖麻、曼陀罗的选择性较弱,对曼陀罗的选择性最弱,选择顺序为马铃薯>烟草>番茄>酸浆>辣椒>龙葵>茄子>蓖麻>曼陀罗。
由图2马铃薯块茎蛾3龄幼虫的非选择性取食试验结果可以看出,处理后1 h时时对供试的马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆的取食率分别为(94.40±3.25)%、(89.70± 4.56)%、(86.70±5.32)%、(80.00±6.25)%、(80.00± 4.35)%、(72.00±5.28)%、(80.00±3.54)%、(76.40± 4.65)%和(82.80±5.42)%,表现出明显的选择性差异(F=36.37,P<0.01)。之后随时间的延长,取食选择性缓慢下降。处理后48 h时,马铃薯块茎蛾3龄幼虫对供试马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆叶片上的取食率降至(93.60±3.54)%、(86.70±2.65)%、(82.80±2.36)%、(64.80±2.65)%、(49.80±2.98)%、(42.50±3.85)%、(56.70±3.56)%、(45.10±3.78)%和(72.00±3.98)%。综合1~48h内马铃薯块茎蛾3龄幼虫对供试的马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆平均取食率,马铃薯块茎蛾3龄幼虫对马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆具有明显的选择性(F=59.34,P<0.01),其选择顺序为马铃薯>烟草>番茄>酸浆>辣椒>龙葵>茄子>蓖麻>曼陀罗。
2.3 马铃薯块茎蛾对受害叶取食的选择性
马铃薯块茎蛾初孵幼虫对受其危害后植物叶片的取食选择性结果表明,释放初孵幼虫1~4 h,对受害马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆表现出了明显的选择性,其中对受害马铃薯、番茄和酸浆叶的选择性强于对其他植物(图3)。之后,随着处理时间的延长,选择性逐渐降低,至24 h时,其对马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆叶片选择率分别为(20.00±2.43)%、(14.16±3.43)%、(14.86±3.46)%、(10.00±2.34)%、(10.00±2.15)%、(5.00±2.34)%、(10.50±2.25)%、(5.00±1.16)%和(10.48±3.25)%,各处理间差异显著(F=21.56,P<0.01)。到48 h时,其对马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆叶片选择率分别为(15.76± 2.34)%、(14.75±3.24)%、(10.74±2.34)%、(9.56± 1.24)%、(9.67±1.24)%、(10.00±2.45)%、(9.76± 1.54)%、(10.00±2.45)%和(9.76±2.32)%,处理间无显著差异(F=2.34,P>0.05)。但从处理后1~48 h内,初孵幼虫对被马铃薯块茎蛾取食危害后的供试植物的取食选择性来看,其选择顺序为马铃薯>番茄>烟草>酸浆>龙葵>辣椒>茄子>蓖麻>曼陀罗。
马铃薯块茎蛾3龄幼虫对受马铃薯块茎蛾危害后的马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆叶片的取食选择性结果(图3)表明,处理后1,2,4 h,3龄幼虫对各受害植物叶片的取食率间差异显著(1 h:F=15.28,P<0.01;2 h:F =18.21,P<0.01;4 h:F=13.65,P<0.01)。处理后第8 h,表现出明显的选择性,对马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆的取食率分别为(30.00±2.15)%、(13.45±1.05)%、(25.00±2.21)%、(5.00±1.13)%、(5.00±0.25)%、(5.00±0.35)%、(5.00±0.48)%、(5.00±0.24)%、(10.00±1.35)%。之后随时间延长,马铃薯块茎蛾3龄幼虫对受害叶片的取食选择性降低,至48 h,对受害马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆的取食率分别为(15.00±3.26)%、(15.00±3.35)%、(10.00±2.64)%、(10.00±1.24)%、(10.00±0.26)%、(10.00±0.28)%、(10.00±0.12)%、(10.00±1.35)%和(10.00±0.26)%,处理间无明显的差异(F=1.76,P>0.05)。但处理1~48 h内,3龄幼虫对被马铃薯块茎蛾取食危害后的供试植物的取食选择性来看,其选择顺序为马铃薯>番茄>烟草>酸浆>龙葵>辣椒>茄子>蓖麻>曼陀罗。
3 讨论
昆虫在长期的演化过程中形成了特定的选择性,不同种类的昆虫,取食食物的种类和范围不同,本试验中选择的马铃薯、烟草、番茄、辣椒、茄子、曼陀罗、龙葵、蓖麻、酸浆是马铃薯种植区常见的植物,研究发现,马铃薯块茎蛾幼虫最嗜食的寄主为马铃薯,该结果与文献报道的马铃薯块茎蛾成虫产卵的最嗜好寄主相同[6,9,10],这也许是马铃薯田发生危害重于烟草田的原因之一。然而,本研究发现,马铃薯块茎蛾幼虫对马铃薯和烟草的取食选择性强于番茄,但据孙红艳等[10]研究发现,马铃薯成虫对马铃薯和番茄的产卵选择性间无显著差异,这种差异的原因还有待进一步研究。
利用作物多样性生态控制害虫的“推(Push)-拉(Pull)”策略就是利用对昆虫具有引诱或驱避作用的植物的配置来达到控制害虫[15],本研究发现,马铃薯块茎蛾幼虫对不同植物的取食选择性不同,高龄幼虫对不同寄主植物的选择性趋于弱化。同时,该虫也是烤烟的重要害虫[4],因此,在马铃薯或烤烟种植时应注意与周围环境中茄子、辣椒等作物的布局,以降低越冬场所及寄主的交替,从而减少马铃薯块茎蛾虫源。此外,马铃薯块茎蛾幼虫对不同寄主植物选择性的差异,也反映了该虫对不同植物的趋向或忌避效应,因而该研究结果对马铃薯块茎蛾的生态防治策略的制定具有重要的参考价值。
植物挥发物在植食性昆虫寄主选择行为中起着重要的作用[16-18],而植物遭受植食性昆虫的进攻时释放的一些挥发性化学物具有明显的间接防御作用[19-21]。然而,有些害虫对不同水稻品种取食选择性的差异与品种叶片物理性状差异有一定的相关性[22]。本研究中,马铃薯块茎蛾初孵幼虫和3龄幼虫对受马铃薯块茎蛾危害后的不同植物叶片的取食物选择性与未受虫害叶片的取食选择性间存在一定差异,这种差异是否与供试验植物受马铃薯块茎蛾危害后挥发的化学挥发物有关,也是值得进一步研究的重要内容。
[1]徐国淦.马铃薯块茎蛾[M].北京:中国农业科学院植物保护研究所.1985:10-55.
[2]王丽珍,孙茂林,Chujo E.云南马铃薯储藏害虫及其特征描述[J].云南大学学报:自然科学版,2002,24(5):398-400.
[3]段玉云,隋启君,孙茂林.云南省马铃薯的生产现状[J].云南农业科技,2003(2):29.
[4]郑朝政.龙山县烟田烟草潜叶蛾的发生为害和综防措施[J].中国植保导刊,2008,28(7):30-31.
[5]袁志华,郭井菲,王振营,等.亚洲玉米螟幼虫对不同寄主植物的取食选择性[J].植物保护学报,2013,40(3):205-210.
[6]任静涛,胡纯华,王旭,等.幼虫粪便提取物对马铃薯块茎蛾雌虫产卵的抑制作用[J].江西农业学报,2007,19(2):77-78.
[7]康敏,谭仲夏,任静涛,等.印楝素与桉叶油醇对马铃薯块茎蛾产卵选择性的影响[J].安徽农学通报,2007,13(3):134-135.
[8]康敏,任静涛,苏鹏娟,等.3种植物提取物对马铃薯块茎蛾产卵的抑制作用[J].安徽农业科学,2007,35(10):2858-2859.
[9]张丽芳,李正跃,王继华.马铃薯块茎蛾交配行为研究[J].江西农业学报,2008,20(9):77-78.
[10]孙红艳,Narayan S.Talekar,等.马铃薯块茎蛾的产卵特性[J].云南农业大学学报:自然科学版,2009,24(3):354-360.
[11]马艳粉,李正跃,任明佳,等.马铃薯块茎蛾对不同寄主植物的产卵选择性比较[J].农药,2010(5):380-382,389.
[12]Gómez Jiménez M I,Poveda K.Synergistic effects of repellents and attractants in potato tuber moth control[J].Basic and Applied Ecology,2009,10(8):763-769.
[13]王冬妍,王振营,何康来,等.转B基因抗虫玉米对亚洲玉米螟幼虫取食行为的影响[J].昆虫知识,2005,42(5):270-274.
[14]唐启义,冯明光.实用统计分析及其DPS数据处理系统.北京:科学出版社,2002.
[15]Samantha MC,ZeyaurRK,John AP.The use ofpush-pullstrategies in integrated pest management[J].Annual Review of Entomology, 2007,52:375-400.
[16]Pichersky E,Noel J P,Dudareva N.Biosynthesis of plant volatiles: Nature'sdiversity and ingenuity[J].Science,2006,311:808-811.
[17]Allmann S,Baldwin I T.Insects betray themselves in nature to predators by rapid isomerization of green leaf volatiles[J]. Science,2010,329:1075-1078.
[18]戴建青,韩诗畴,杜家纬.植物挥发性信息化学物质在昆虫寄主选择行为中的作用[J].环境昆虫学报,2010,32(3):407-414.
[19]蔡晓明,孙晓玲,董文霞,等.虫害诱导植物挥发物(HIPVs):从诱导到生态功能[J].生态学报,2008,28(8):3969-3980.
[20]Hare JD.Ecologicalroleofvolatilesproducedbyplantsin responseto damagebyherbivorousinsects[J].AnnualReviewofEntomology,2011, 56:161-180.
[21]KesslerA,Baldwin IT.Defensive function ofherbivore-induced plant volatile emissions in nature[J].Science,2001,292:2141-2144.
[22]Khan Z R,Saxena R C.Effect of steam distillate extracts of resistant and susceptible rice cultivars on behavior of Sogatella furcifera(Homoptera:Delphacidae)[J].Journal of Economic Entomology,1986,79:928-935.
Feeding Selectivity of Phthorimaes operculella(Zeller)on Different Host Plants
WANG Haoyuan1,ZHANG Limin2,CHEN Bin1*,LI Zhengyue1*
(1.College of Plant Protection,Yunnan Agricultural University,Key Laboratory of Agro-biodiversity and Pest Management of Education Ministry of China,Kunming,Yunnan 650201,China;2.College of Basic Science and Information Engineering, Yunnan Agricultural University,Kunming,Yunnan 650201,China)
The feeding selectivity of neonates and the third instar larvae of the potato tuber moth Phthorimaes operculella(Zeller)on Solanum tuberosum L.,Nicotiana tabacum,Lycopersicon esculentum,Capsicum annuum,Solanum melongena L., Datura stramonium L.,Solanum nigrum L.,Lonicera japonica,and Physalis alkekengi was studied using choice and no-choice feeding test under the leaf plate method in laboratory.Results showed that the neonate and 3rd instar larvae of P.operculella showed significant different feeding selectivity for the tested S.tuberosum,N.tabacum,L.esculentum,C.annuum,S. melongena,D.stramonium,S.nigrum,L.japonica,and P.alkekengi(Neonate:F=60.94,P<0.01;3rd instaralrvae:F=66.34,P<0.01)during 1-48 h,with a selection rank of S.tuberosum>N.tabacum>L.esculentum>P.alkekengi>C.annuum>S. nigrum>S.melongena>L.japonica>D.stramonium.The neonates and 3rd instar larvae of P.operculella were also showed feeding selectivity forthe differentleaves afterfeeding by the larvae of P.operculella.The feeding selectivity decreased with the increase ofthe treatmenttime,and nosignificantdifferencewas foundforthe feed selectivity forthe differentplants(Neonate:F= 2.34,P>0.05;3rd instaralrvae:F=1.76,P>0.05)at48 h aftertreatment.The average of selection rank was S.tuberosum>L. esculentum>N.tabacum>P.alkekengi>S.nigrum>C.annuum>S.melongena>L.japonica>D.stramonium during 1-48 h. The results willprovide a base fortheecologicalcontrolon the P.operculella by using the plantdiversity.
Phthorimaes operculella(Zeller);larvae;feeding selectivity;feeding percentage
S532
B
1672-3635(2013)04-0226-06
2013-07-21
国家基础研究计划(973)项目(2011CB100404);国家自然科学基金项目(31160363);云南省科技创新团队项目(2011HC005);云南省现代农业马铃薯产业技术体系项目[云农科(2009)53号]。
王浩元(1989-),男,硕士研究生,主要从事农业害虫综合防治研究。
陈斌,教授,从事昆虫生理与害虫综合治理研究,E-mail:chbins@163.com;李正跃,教授,从事昆虫生态与害虫综合治理研究,E-mail:lizhengyue@163.net。
猜你喜欢
TMR Modern Herbal Medicine(2022年3期)2022-09-06
天天爱科学(2022年4期)2022-05-23
食品工业科技(2021年19期)2021-10-29
现代农业研究(2021年4期)2021-04-11
老友(2020年11期)2020-11-28
中国诗歌(2018年3期)2018-11-14
中成药(2017年7期)2017-11-22
创新作文(3-4年级)(2015年8期)2015-09-15
绿色科技(2012年11期)2012-11-16
农家科技中旬版(2011年3期)2011-06-13
