
河西走廊不同红砂天然群体种子表型性状相关性研究
2013-04-10 06:13苏世平李毅种培芳高茜
草业学报 2013年1期
苏世平,李毅,种培芳,高茜
(甘肃农业大学林学院,甘肃 兰州730070)
植物的性状是由基因型和环境共同作用的结果,可以通过测定表型或进行遗传标记等多种手段进行多样性研究,其中用表型性状来测定是最现实和直接的方法[1],因为表型变异必然蕴藏着遗传变异,对植物表型性状进行研究不仅能初步了解种群遗传变异的大小,同时也有助于了解生物适应和进化的方式。
同一植物,由于长期处在不同的生境下,受到各种条件的限制,在形态和生理上出现不同程度的变化,这种变化与植物所处环境的生态因子、群体间的距离以及群体内的生殖方式有关,一般认为,生境越复杂、群体间距离越远、异花授粉程度越高则表型变异越大,可能存在的遗传变异也越大。利用形态性状来研究物种遗传变异是简便而易行的手段,可以直接利用野外采集的样本来进行分析[2]。
红砂(Reaumuriasoongorica)属柽柳科(Tamaricaceae)红砂属(Reaumuria)的落叶超旱生半灌木,主要分布在年降水量在60~300mm、海拔500~3 200m的荒漠、半荒漠的山麓洪积平原、山地丘陵、剥蚀残丘、山前砂砾质和砾质洪积扇、戈壁等地,是干旱荒漠区分布最广的植物种之一,具有固沙、固土的优良特性,以其为建群种的植被群落是干旱荒漠中的主要植被类型之一,是保护干旱荒漠化土地的重要生物屏障[3-5]。但是,由于干旱和人为破坏,导致以红砂为建群种的植被群落自然更新速度缓慢,使得群落的稳定性下降,迫切需要恢复与重建。通过对天然群体红砂种子性状的研究,揭示其种内变异规律,为今后开展种质资源收集与保存、遗传评价及种质创新利用以及人工辅助更新及群体遗传多样性研究提供理论依据[6]。
在红砂的研究方面,前人主要集中于繁殖特性[4,5]、群落更新[3]、群落稳定性[4-7]以及不同种群生理特性[8,9]等方面,而对于不同群体红砂遗传多样性方面的研究比较少见[10-12]。因此进行红砂群落种子表型多样性的研究,可揭示红砂表型变异程度和变异规律,可为今后该树种种质资源的保护与开发利用提供依据[4,12-14]。
1 材料与方法
1.1 群体的确立与试验材料的采集
按经度、纬度和海拔梯度设置样地,于2007年10月完成所有群体种子的采集工作。在河西走廊红砂天然分布区内,自西向东共选取10个群体(图1):酒泉1(JQ1)、酒泉2(JQ2)、酒泉3(JQ3)、武威1(WW1)、武威2(WW2)、武威3(WW3)、武威4(WW4)、张掖1(ZY1)、张掖2(ZY2)、张掖3(ZY3)。每群体选取15株生长正常、具有代表性的单株进行采种,株间距离5m以上;在每个单株上采收所有种子带回实验室进行试验。
样地中实测经纬度、海拔;年均温、1月均温、7月均温和年降水量等气象因子从当地气象部门查询(2007年气象数据)(表1)。
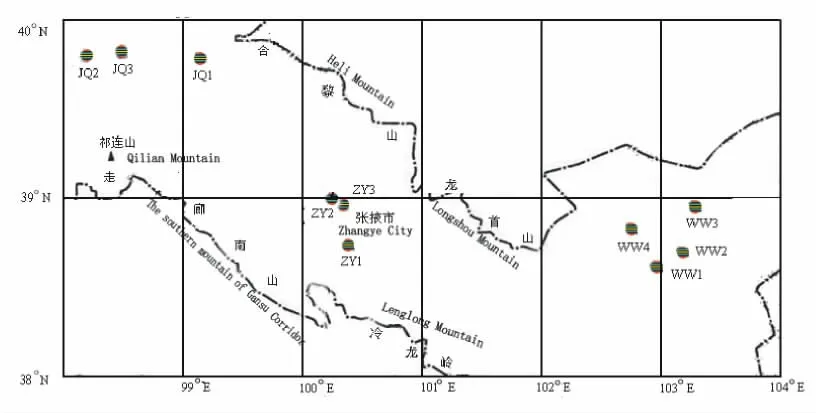
图1 红砂采种分布图Fig.1 The plots of R.soongoricaseed collecting
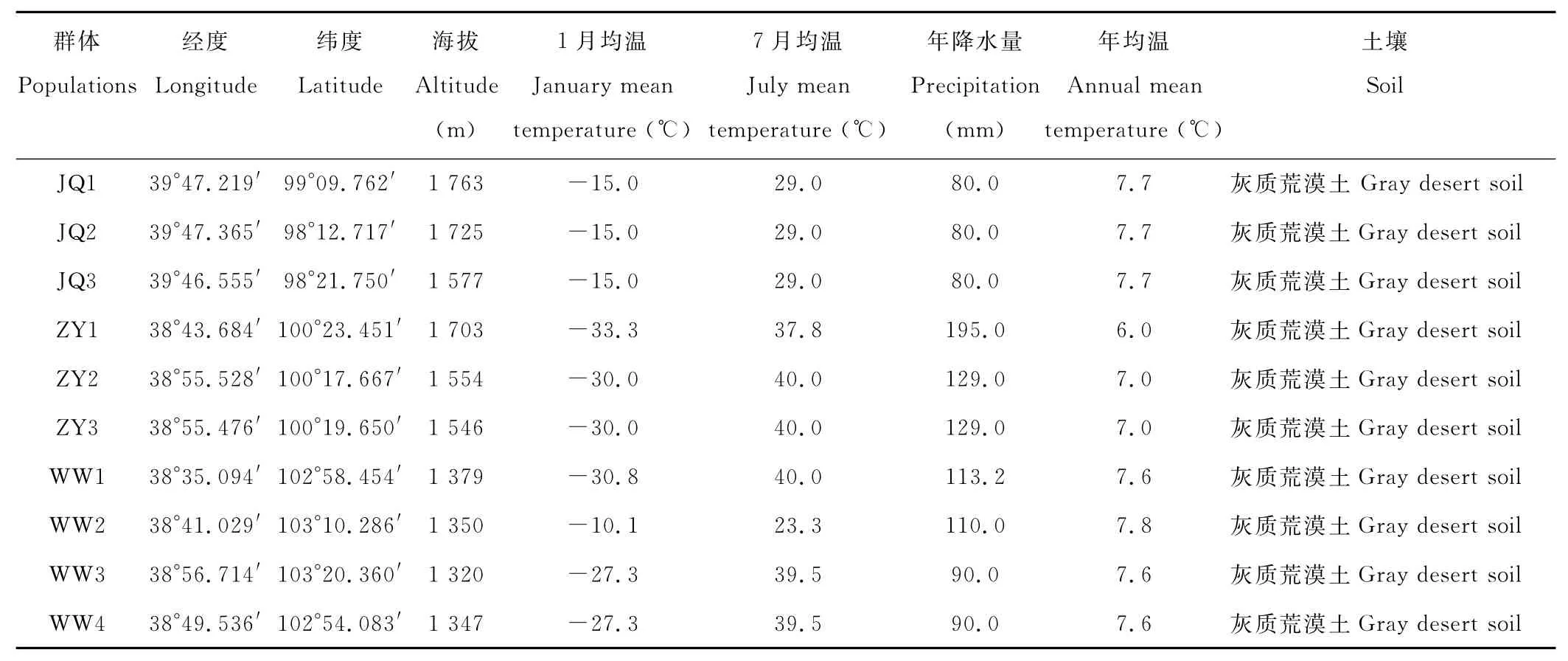
表1 红砂采样群体地理位置、生态因子及样地简介Table 1 Locations and releated ecological factors of the sampled populations
1.2 种子表型性状的测定方法
千粒重:在10个群体中进行每株测定。每群体测定15株,每株抽取3 000粒种子分成3等份测量千粒重。取平均值作为单株千粒重。
种子长(Ls)、种子宽(Ws):每群体15个单株,每单株随机取20粒种子,用浓硫酸腐蚀掉种毛,将去掉种毛后的种子置于坐标纸上进行数码拍照,种子长宽用CAD软件进行测量,按实际比例进行换算。

式中,y1为沿种子长方向坐标纸单位长度CAD测量值,y2为种子长CAD测量值,y3为沿种子长方向坐标纸单位长度。
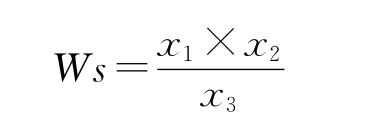
式中,x1为沿种子宽方向坐标纸单位长度CAD测量值,x2为种子宽CAD测量值,x3为沿种子宽方向坐标纸单位长度。
1.3 统计分析方法
对各性状采用巢式设计方差分析[15-19],线性模型为:
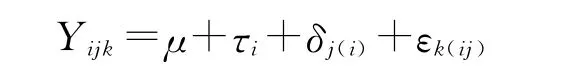
式中,Yijk为第i个群体第j个家系第k个观测值,μ为总平均值,τi为群体间效应值,δj(i)为群体内家系效应值,εk(ij)为随机误差。
表型分化系数(Vst)[18]反映群体间表型分化的值,即群体间变异占遗传总变异的百分比。

式中,σ2t/s为群体间的方差分量,σ2s为群体内的方差分量。
用变异系数CV表示表型性状的离散性特征,变异系数越大,性状值离散程度越大。
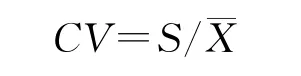
式中,S为标准差,珡X为平均值[19]。
为了消除量纲使性状间具有可比性,采用相对极差R′i表示各群体内的极端变异程度。
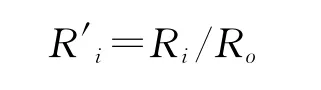
式中,Ri为群体内的极差,Ro为性状总极差。
采用双变量相关分析法对红砂各性状与采集点地理生态因子之间进行相关分析。利用欧氏平均距离法进行分层聚类分析。
上述的数据统计均利用Excel 2003,SPSS 11.5软件完成。
2 结果与分析
2.1 红砂群体间和群体内的形态变异特征
红砂种子4个表型性状在群体间都存在极显著差异,在群体内差异不显著(表2)。各性状平均值和多重比较(表3)表明,群体中4个性状间存在显著的差异。千粒重最大的群体是WW4,为2.42g,最小的是JQ3,为1.07g,种子长最大的群体是 WW4,为3.496mm,最小的群体是ZY1,为2.532mm,种子宽最大的群体是JQ3,为1.364mm,最小的群体是ZY1,为0.850mm,种子长宽比最大的群体是 WW4,为3.977,最小的群体是JQ3,为3.006。
2.2 红砂种子形态变异特征
各群体表型性状变异系数(表4)表明,4个种子性状平均变异系数为0.019 3,变异幅度0.001 8~0.068 5。各表型变异系数由大到小依次为:千粒重(0.068 5)>种子宽(0.004 0)>种子长宽比(0.003 1)>种子长(0.001 8),这表明千粒重在群体间变化最大,种子宽最小。
同一性状在不同群体内的变异幅度也有差异,说明不同地区的环境异质性导致表型产生差异。10个群体所有性状的平均变异系数从大到小的排序为:ZY1(0.048 4)>ZY2(0.037 1)>WW2(0.025 5)>WW4(0.018 1)>JQ3(0.017 7)>WW1(0.010 8)>JQ1(0.010 4)>ZY3(0.010 3)>WW3(0.008 3)>JQ2(0.006 6)。变异系数最大的群体(ZY1)要比最小的群体(JQ2)大近7倍多。说明ZY1的表型多样性较高,而JQ2的表型多样性程度较低。
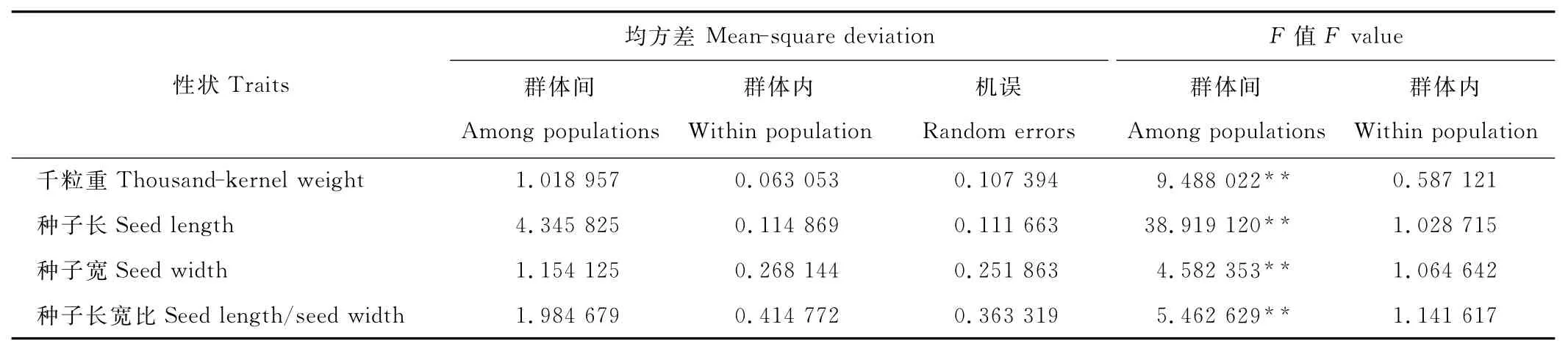
表2 红砂群体间群体内表型性状的方差分析结果Table 2 Variance analysis of phenotypic traits among population/within population
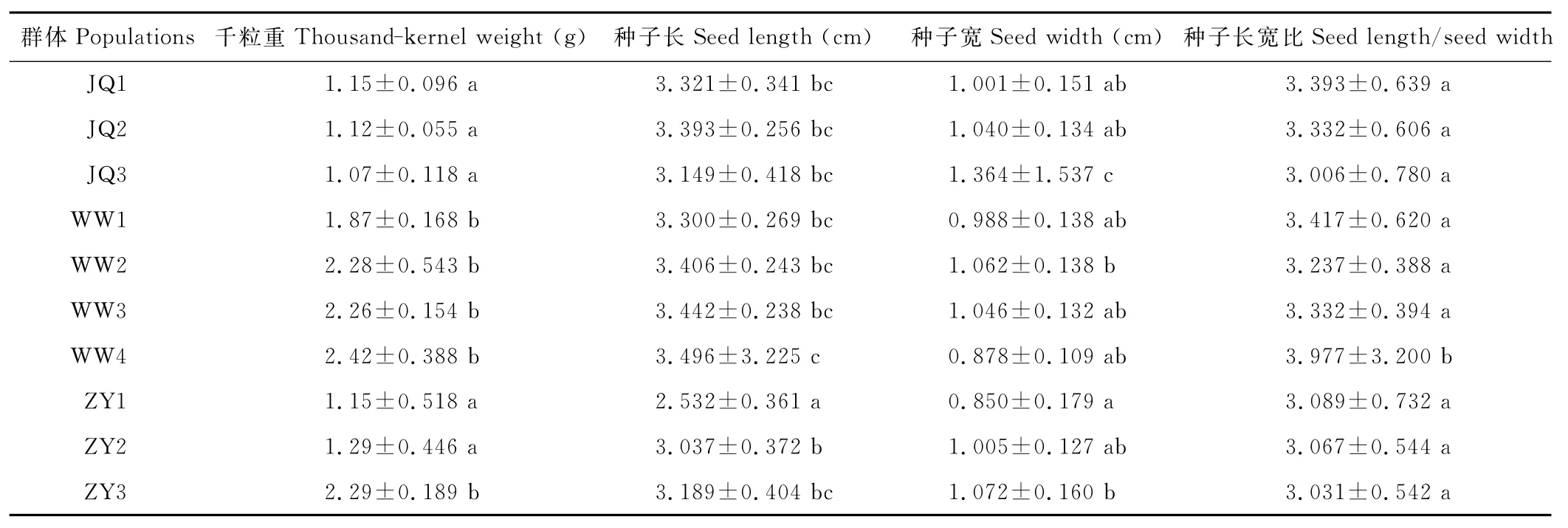
表3 10个天然群体表型性状的平均值和标准差Table 3 The mean value and standard deviation of phenotypic traits of 10populations
2.3 各群体表型性状的相对极差
4个表型性状的平均相对极差(表5)为40.369 0,相对极差最大的性状是种子长(R′i=65.541 5),相对极差最小的是种子宽(R′i=17.228 9),说明种子长在群体内的极端变异程度最大,而种子宽在群体内变异程度比较稳定。4个表型性状的相对极差排序为:种子长>种子长宽比>千粒重>种子宽。10个群体之间相对极差的变化比较大,按相对极差从大到小的顺序为:JQ3>ZY1>ZY2>JQ1>ZY3>WW2>WW4>WW1>JQ2>WW3。
2.4 红砂天然群体间的表型分化及各形态特征变异来源
表型性状的表型分化系数(Vst)变异幅度为68.312 458%~99.247 950%(表6),其中最大的是种子长,Vst=99.247 950%,最小的是千粒重,Vst=68.312 458%,说明4个性状在群体间的变异大于群体内的变异,群体间的变异占绝对优势。

表4 红砂天然群体表型性状的变异系数Table 4 Variation coefficient of phenotypic traits of R.soongorica populations
2.5 红砂表型性状和采集地的地理生态因子的相关关系
对4个性状与其分布地地理生态因子偏相关分析与检验结果表明(表7),千粒重、种子长、种子长宽比与经度呈极显著正相关关系,种子宽与经度呈极显著负相关。千粒重与纬度呈显著负相关,种子长、种子宽、种子长宽比与纬度呈极显著的正相关关系。千粒重、种子长、种子宽与海拔呈极显著负相关,种子长宽比与海拔呈显著正相关。种子长与1月均温、7月均温呈显著正相关,与年均温、年降水量呈极显著正相关。
这说明千粒重、种子长和种子长宽比3个表型性状随着经度的升高而升高,种子宽随经度的减小而增加;种子长、种子宽和种子长宽比随纬度升高而升高。即由西到东,千粒重、种子长、种子长宽比都有所增大,由北到南,红砂种子长、种子宽、种子长宽比都有所降低,千粒重有所升高;从高海拔到低海拔,除种子长宽比外,其他性状均有升高的趋势。
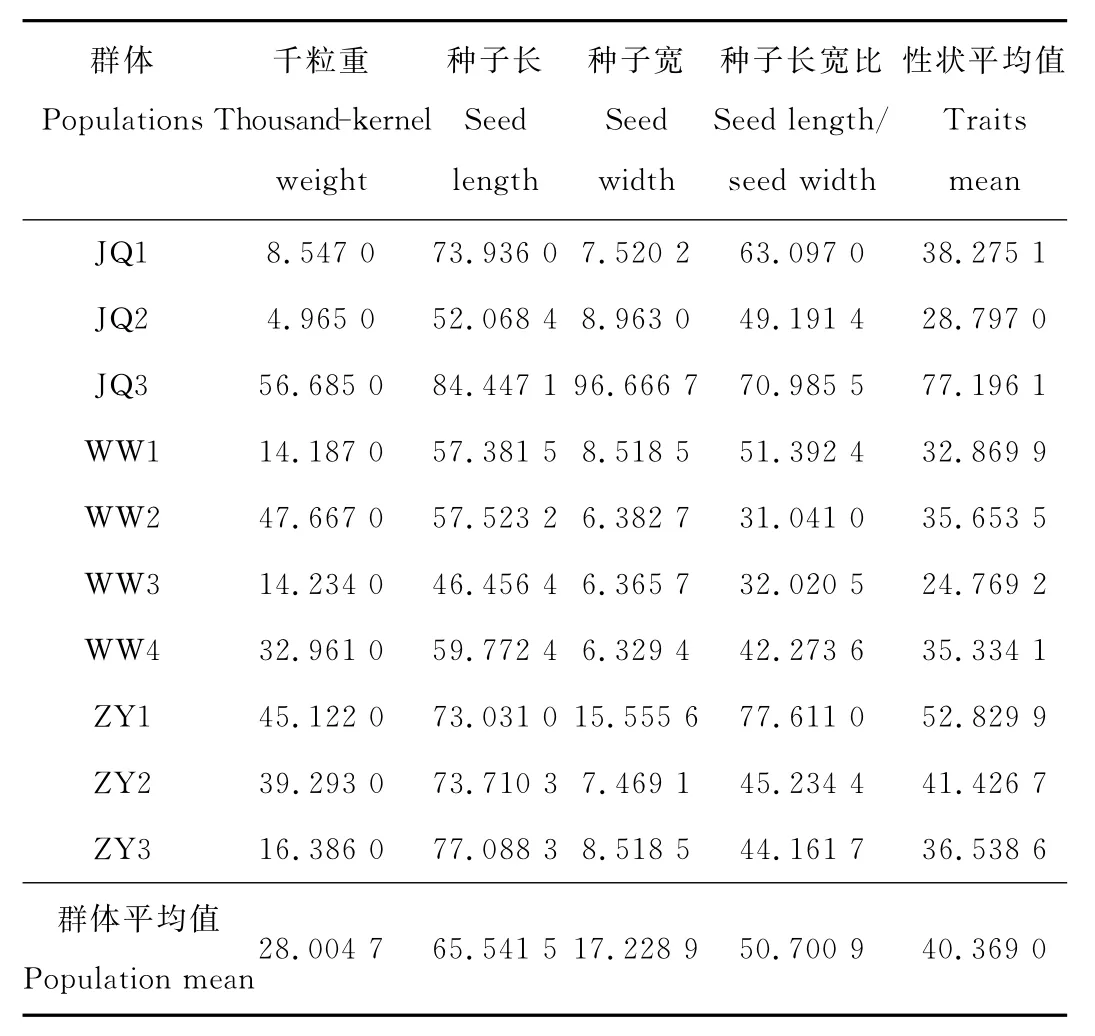
表5 红砂天然群体表型性状的相对极差(R′i)Table 5 Relative extreme value differences of phenotypic traits in R.soongorica populations %
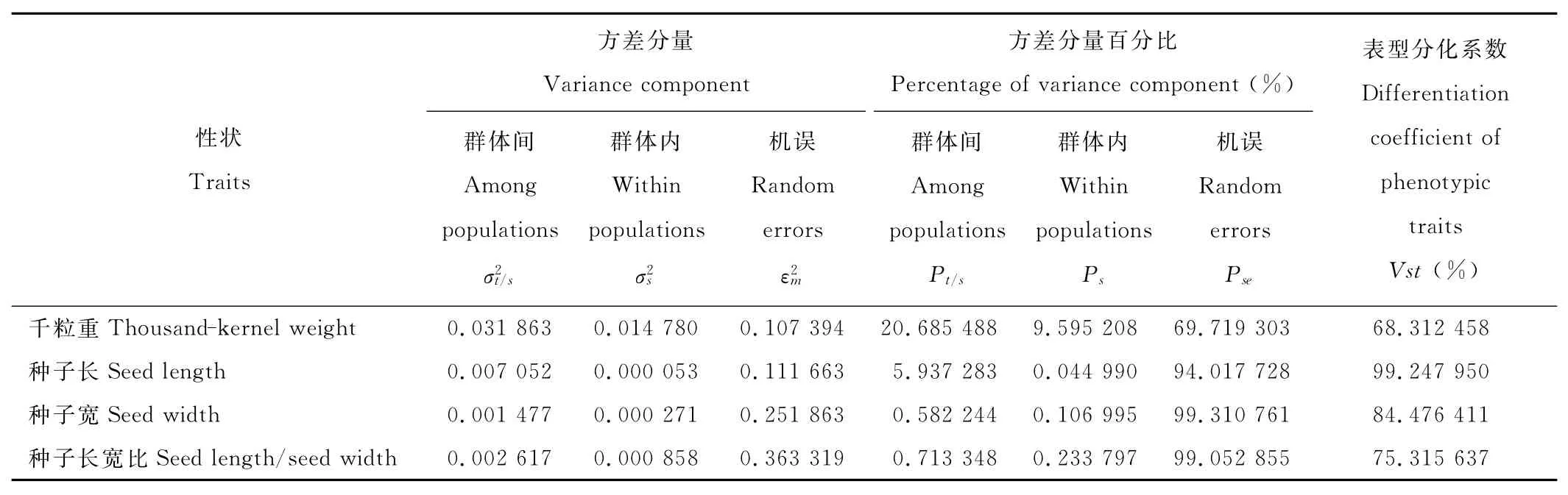
表6 表型性状的方差分量及群体间表型分化系数Table 6 Variance portions and differentiation coefficients of phenotypic traits among/within populations

表7 红砂表型性状与地理生态因子间的相关关系Table 7 Correlation coefficients matrix between the phenotypic traits and geo-ecological factors in R.soongorica
2.6 红砂天然群体表型聚类分析
利用欧氏平均距离法,对各群体4个表型性状数据进行聚类分析(图2)。根据聚类结果将10个群体划分为4个类群。将JQ1、JQ2、ZY2聚为一类,WW1、WW2、WW3、WW4和 ZY3聚为一类,JQ3、ZY1单独归为2类。
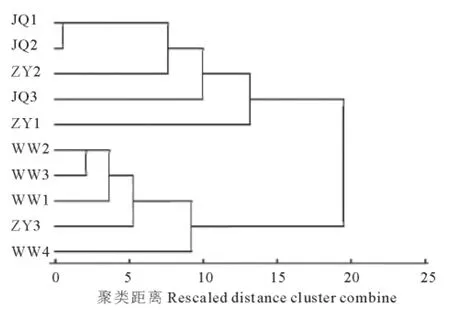
图2 红砂天然群体表型性状聚类Fig.2 UPGMA cluster based on the phenotypic traits of 10 populations
3 结论与讨论
3.1 群体间表型变异产生的原因
红砂不同群体间同一性状都存在着广泛差异,说明群体表型的差异是由不同地区的环境异质性所导致,这表明其性状稳定性较差并受到一定环境压力的影响[20,21],导致群落内差异不显著的结果有可能与其繁殖方式有关,也可能与群落所处立地环境有关,也可能与群体之间的地理距离有关。曾彦军等[5]报道红砂的繁殖方式主要受其生长环境和群落的年龄结构影响,生长于干旱、沙化生境的成熟红砂群落以无性繁殖为主,无性繁殖占种群比例的72.4%,处于相对湿润的幼龄红砂种群以有性繁殖为主,这说明,在河西走廊这种干旱环境下,无性繁殖导致种群性状高度一致,遗传基础趋于一致化;黄培祐等[22]对荒漠地区红砂繁殖方式的研究表明,红砂种子和幼苗建植需要较湿润的环境,这也说明,干旱少雨导致红砂种子难以萌发和幼苗难以成活,从而降低了通过有性繁殖所产生的表型分化,因此导致群体的遗传基础变窄,本研究与此结果相似,从降水量看,张掖1的年降水量为195mm,为10个群体中最高,该群落的变异系数均比其他群体高,这也表明,湿润环境可提高种子和幼苗的建植;表型的这种变异模式也可能与生殖的地域隔离有关,由于各群体之间间隔距离比较大,导致群落之间没有或者很少有基因交流,经过长期的繁衍,从而导致这种群落分布格局。
3.2 表型变异的梯度规律性
树种分布范围的大小是影响树木地理变异的主要因子。一般而言,分布区较大,则变异大;分布范围小则变异小。另外,树种自然分布区内环境因素多样性也是影响林木地理变异的因子。分布区的环境条件越复杂,则种内群体的遗传变异越大。研究结果表明红砂种子性状变异呈现以水平地带性和垂直地带性变异为主的梯度规律性,该研究结果与前人在云杉(Piceaasperata)[23]、青海云杉(Piceacrassifolia)[19]、白皮松(Pinusbungeana)[15]等的研究结果相似,导致这种结果发生的原因可能与河西走廊不同区域年降水量、1月和7月均温的不同均有关。所抽样群体在降水上整体趋势从酒泉、武威到张掖呈陡减趋势,7月和1月均温绝对值都呈升高趋势,这也可能导致种子体内内含物、营养物质的积累由于环境因素不同而差异悬殊,所以才有这种变异趋势。
尽管群体内表型的变异不显著,但群体间变异的意义却大于群体内的变异,因为存在于群体间的变异反映了地理、生殖隔离上的差异,群体间的多样性变异是种内多样性的重要组成部分。分布在群体间的变异真正反映了群体在不同环境中的适应状况,其大小在某种程度上说明了该生物对不同环境适应的广泛程度,值越大适应的环境越广[20]。
3.3 种质资源保存的艰巨性
根据红砂丰富的自然变异,可以初步推断其改良前景是广阔的。红砂表型性状68% 以上的变异来自群体间,32%的变异存在于群体内,群体间的分化明显高于群体内,造成这种变异的原因可能与植物自身的繁殖对策和环境因素有关。群落间的这种变异为生物多样性保护和优良种质资源的利用提供了物质基础,但是群落内部这种极大的相似性使群落稳定性和抗御自然灾害的能力降低,群落内部的生物多样性指数下降,这种多层次的变异为种质资源保存和生物多样性保护提供了物质基础,同时也为遗传改良工作提出了新的要求,即在遗传改良工作中进行优良种源、优良群体选择和利用的同时,应加大优良个体选择和利用的力度[24,25],同时也表明了生物多样性保护任务的艰巨性,这就要求如何在保持和发展群体间这种广泛变异的基础上发掘和扩大群体内的变异,使得群体内和群体间均有丰富的遗传多样性。
[1] 李文英,顾万春.蒙古栎天然群体表型多样性研究[J].林业科学,2005,41(1):49-56.
[2] 张恒庆,安利佳,祖元刚.天然红松种群形态特征地理变异的研究[J].生态学报,1999,19(6):932-938.
[3] 刘家琼,邱明新,蒲锦春,等.我国荒漠典型超旱生植物——红砂[J].植物学报,1982,24(5):485-488.
[4] 曾彦军,王彦荣,张宝林,等.红砂种群繁殖特性的研究[J].草业学报,2002,(2):66-71.
[5] 曾彦军,王彦荣,庄光辉,等.红砂和霸王种子萌发对干旱与播深条件的响应[J].生态学报,2004,24(8):1629-1634.
[6] 周资行,李毅,焦健.兰州市南北两山不同生境红砂种群数量动态研究[J].自然资源学报,2011,26(10):1726-1737.
[7] 周资行,李毅,焦健.兰州市郊红砂种群数量动态与分布格局研究[J].生态与农村环境学报,2011,27(8):24-31.
[8] 高暝,李毅,种培芳,等.渗透胁迫下不同地理种源白刺的生理响应[J].草业学报,2011,20(3):99-107.
[9] 左利萍,李毅,焦健.水分梯度下不同地理种群红砂叶片生理特征比较研究[J].中国沙漠,2009,29(3):514-518.
[10] 冯亮亮,唐红,李毅,等.甘肃红砂不同种群遗传多样性的ISSR分析[J].草业学报,2011,20(1):125-130.
[11] 苏世平,李毅,种培芳.河西走廊不同红砂天然群体种子活性相关性[J].生态学报,2012,32(1):135-141.
[12] 李秀玲,陈健,王刚.西北地区红砂种群ISSR遗传变异的空间自相关分析[J].中国沙漠,2008,28(3):468-472.
[13] 朱恭,王万鹏.红砂种群自然更新与人工辅助恢复机理的初步研究[J].甘肃农业大学学报,2004,39(4):427-433.
[14] Ye M S,Guan W B,Wu B,etal.Plant community complexity in the arid valley of Minjiang River,southwestern China[J].Acta Ecologica Sinica,2006,26(10):3159-3165.
[15] 李斌,顾万春,卢宝明.白皮松天然群体种实性状表型多样性研究[J].生物多样性,2002,10(2):181-188.
[16] 李梅,韩海荣,康峰峰,等.山西灵空山辽东栎种群叶性状表型变异研究[J].北京林业大学学报,2005,27(5):10-16.
[17] 李文英,顾万春.蒙古栎天然群体表型多样性研究[J].林业科学,2005,41(1):49-56.
[18] 葛颂.同工酶与林木群体遗传变异研究[J].南京林业大学学报,1988,1:68-72.
[19] 王娅丽,李毅,陈晓阳.祁连山青海云杉天然群体表型性状遗传多样性分析[J].林业科学,2008,44(2):70-77.
[20] 周连第,兰彦平,曹庆昌,等.板栗叶片性状表型多样性研究[J].林业科学,2005,21(9):136-139.
[21] 李梅.辽东栎天然群体表型多样性研究[D].北京:北京林业大学,2005.
[22] 黄培祐,聂湘萍,周建民,等.准噶尔盆地中部琵琶柴群落的生境研究[J].新疆大学学报:自然科学版,1988,5(3):66-71.
[23] 罗建勋,顾万春.云杉表型与同功酶遗传多样性研究进展[J].林业科学研究,2004,17(2):255-262.
[24] 梁绪振,鄢家俊,白史且,等.斑茅种质资源形态性状的变异研究[J].草业科学,2011,28(7):1307-1314.
[25] 张妙青,王彦荣,张吉宇,等.垂穗披碱草种质资源繁殖相关特性遗传多样性研究[J].草业学报,2011,20(3):182-191.
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
河南科学(2020年3期)2020-06-02
现代园艺(2017年21期)2018-01-03
中国康复理论与实践(2015年10期)2015-12-24
草业科学(2015年9期)2015-12-11
百科知识(2015年18期)2015-09-10
医学研究杂志(2015年5期)2015-06-10
