
塔里木河下游漫溢干扰频次和持续时间对河岸植被和土壤的影响差异
2013-04-10 06:13傅荩仪徐海量赵新风白元王希义
草业学报 2013年6期
傅荩仪,徐海量,赵新风,白元,王希义
(1.中国科学院新疆生态与地理研究所 绿洲生态与荒漠环境重点实验室,新疆 乌鲁木齐830011;2.中国科学院大学,北京100049)
漫溢干扰(flooding disturbance)与“水淹(waterlogging)”[1],“洪水干扰(flood disturbance)”[2-3]以及“洪水脉冲(flood pulse)”[4]等概念相近。作为影响河流水文生态系统的重要因素之一,其对河岸植物群落的构建和分布亦具有决定性作用。漫溢干扰及其生态影响已成为国内外生态学、水文学和土壤学等多学科交叉的热点问题之一。近年来学者们就漫溢干扰在植物群落结构[5-7]、物种多样性[8-9]、土壤种子库[10]等方面上的生态影响开展了大量野外实地调查与模拟实验研究,其中一些研究结果已表明漫溢干扰在不同频次、持续时间、水量和深度等条件下对植物群落的影响亦存在一定的差异性[11-13]。然而,鉴于天然漫溢干扰的波动性与多变性,一些人为控制实验在实验区内较难开展,以至于很多研究对漫溢强度、频度等指标的定义不够定量化;其次漫溢干扰的影响程度和范围受气候、地势、土壤等众多环境条件的制约,致使国内外不同研究区的漫溢研究结果之间无法建立很好的关联性。此外,涉及干旱区、半干旱区内漫溢干扰对荒漠植被的研究相对较少,缺乏对荒漠植被生态响应的长期监测与综合分析。因此,深入开展和完善漫溢干扰研究不仅有利于推进河流水文过程的保护和调控,对漫溢区植物群落恢复重建以及生物多样性保护更具有十分重要的参考价值。
塔里木河下游地区是我国乃至世界范围内生态退化最为严重的地区之一,其极度干旱恶劣的环境条件和独特严峻的生态问题已引起国内外学者们的广泛关注。以生态恢复为目的的塔里木河下游生态输水工程自2000年实施以来,使得下游干涸已久的河道内及河道两旁出现了漫溢现象,漫溢区内的环境条件发生改变,河岸衰败植被得到一定程度的恢复。由此,本研究将下游研究区内所经历的漫溢频次和持续时间划分为较为具体的不同梯度,基于野外样地调查结果分析探讨不同漫溢频次和持续时间分别对河岸荒漠植被的群落结构、物种多样性以及土壤性质的影响,提出最适宜植被恢复的漫溢频次和持续时间,以期为塔里木河下游水资源调控和植被恢复提供理论依据和实践指导。
1 材料与方法
1.1 研究区概况
研究区位于塔里木河下游段(大西海子—台特玛湖)(39°30′~40°34′N,87°33′~88°27′E),属暖温带大陆性荒漠气候,气候干燥,多风沙天气,区域多年平均气温10.6~11.5℃,日照时数约3000h。年均降水量为17.4~42.0mm,蒸发量为2500~3000mm,是我国极端干旱地区之一[14]。区域代表性植被有胡杨(Populuseuphratica)、柽柳属(Tamarixspp.)、芦苇(Phragmitesaustralis)、罗布麻(Apocynumvenetum)和花花柴(Kareliniacaspia)等[15]。1972年大西海子水库的建成导致下游河道长期断流。2000年起实施的塔里木河下游生态输水工程使得河道沿岸分布大片的漫溢区域,区内土壤条件和植物群落发生显著变化。
1.2 调查样地选取
在塔里木河下游选取9个典型断面进行地表植被调查(图1)。表1显示的是塔里木河下游近12次生态输水的基本资料。由于第3次、第5次、第7次和第11次输水均分两个阶段进行,因而共统计为16次输水。其中16次输水均流经(或止于)英苏,15次输水流经(或止于)喀尔达依,12次输水流经(或止于)阿拉干,10次输水流经(或止于)考干。因此累计过水次数表现为:英苏>喀尔达依>阿拉干>考干。
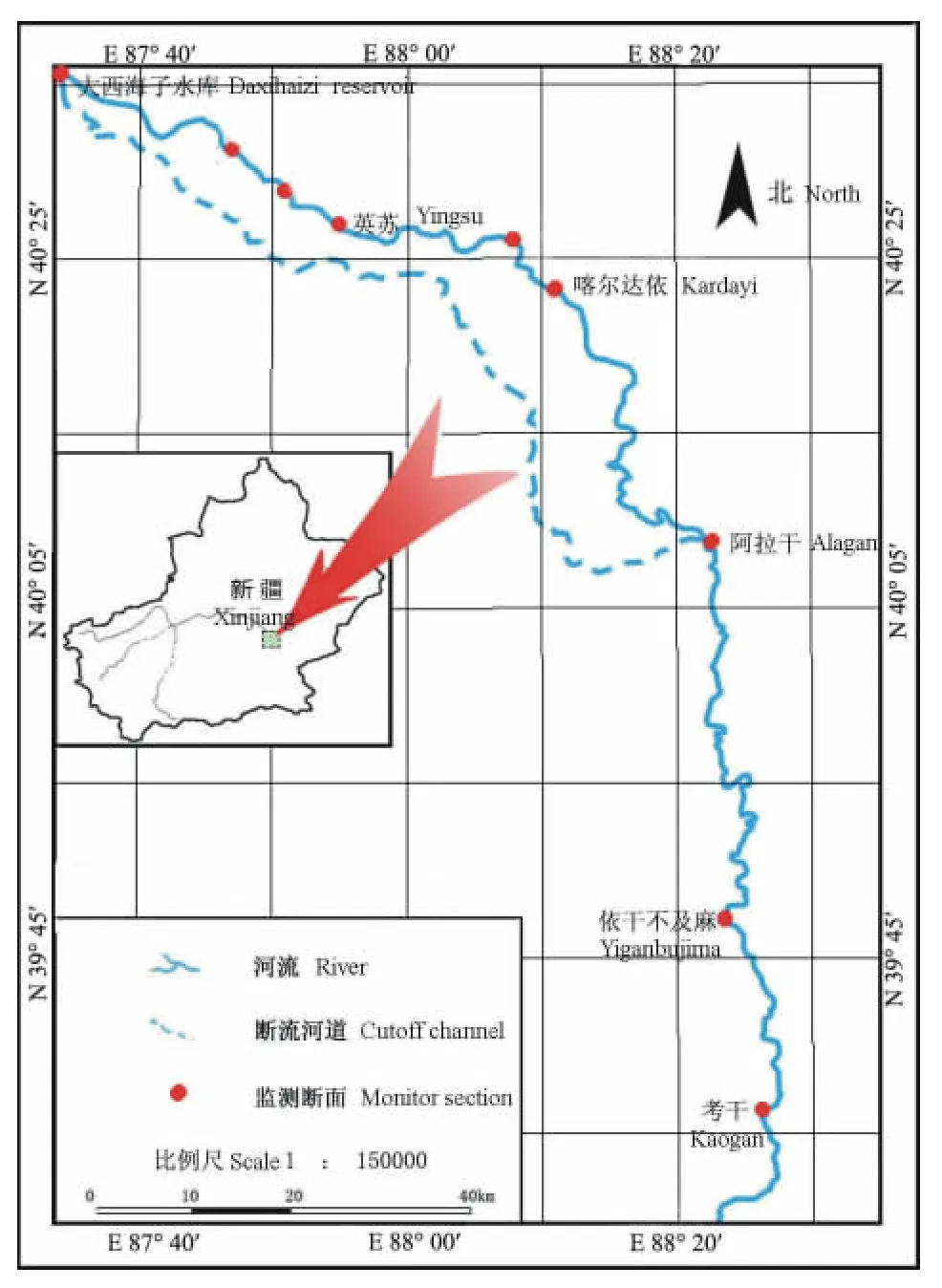
图1 研究区分布图Fig.1 Sketch map of study region
同时在野外实地调查中对各个监测断面及附近几个典型漫溢样地的实际漫溢情况进行记录和整理(表2)。由此,本研究对以上监测断面及3个另设样地的漫溢干扰进行频次和持续时间梯度的划分(表3),其中持续时间梯度按照平均1年内的漫溢天数进行划分。
基于漫溢干扰频次和持续时间梯度的划分,在监测断面和另设样地内进行相应漫溢频次或持续时间梯度的样地设置,且在无漫溢干扰条件下的临近地区设置对照样地。漫溢样地和对照样地均设置为50m×50m。总共设置55个调查样地,其中漫溢样地35个,对照样地20个。
1.3 野外植被调查
2012年5月中旬和7月下旬在所设置的样地内进行地表植被和土壤调查。将每个样地(包括漫溢样地和对照样地)划分成4个25m×25m的植被样方,调查样方内所出现的乔灌木的物种数、个体数、株高、胸径和冠幅;同时在每个样方内任意选取3个1m×1m的草本样方,调查草本植物的物种数、个体数和株高。此外,在每个监测样地内随机挖取0~10cm地表土壤样品3个,取样面积为10cm×10cm。将土样装袋带回实验室进行电导率和有机质的测定。
1.4 数据统计分析
基于野外调查结果,计算漫溢样地和对照样地中一年生草本、多年生草本和乔灌木的重要值,并求出同一漫溢频次或持续时间梯度下3类植被重要值的算术平均值,以作为分析植物群落结构变化的依据;同时选取Simpson指数、Shannon-Wiener指数和Margalef指数来反映植被物种多样性在不同漫溢频次和持续时间梯度下的变化情况。计算公式如下[16-18]:

式中,Pi表明第i个物种的出现频率,Pi=Ni/N;N为样方的总个体数,Ni为第i种的个体数;S为样方的总物种数。利用SPSS 16.0软件对不同监测样地间的植被重要值、多样性指数、土壤电导率和有机质进行ANOVA方差分析和LSD多重比较,以分析不同漫溢干扰频次和持续时间梯度对植被特征和土壤指标的影响差异。
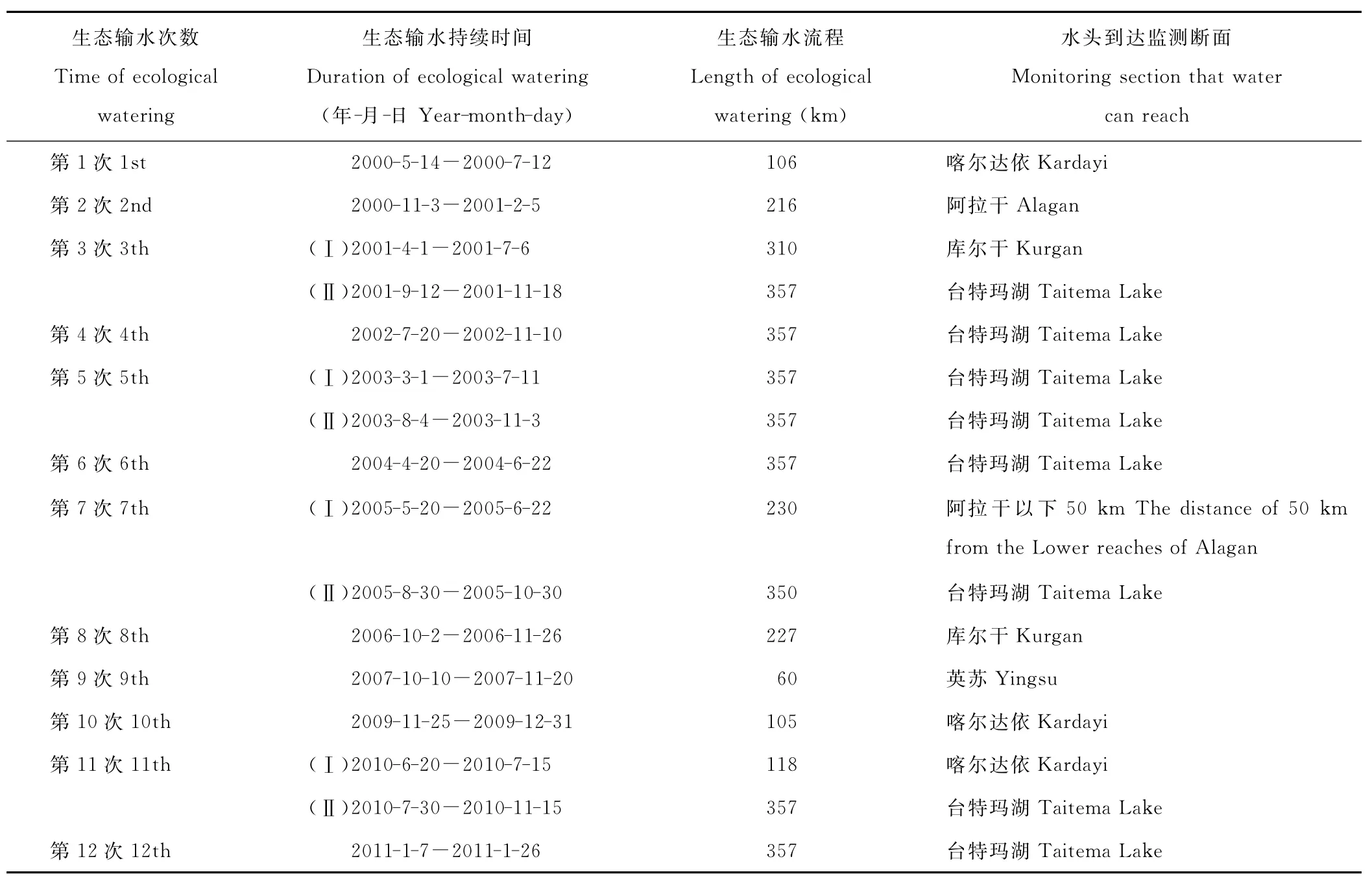
表1 塔里木河下游12次生态输水统计表Table 1 Statistical table of ecological watering to the Lower Tarim River
2 结果与分析
2.1 漫溢干扰频次和持续时间对植物群落结构的影响
无漫溢条件下,乔灌木在群落组成中占据优势,其重要值为0.879(表4)。对照样地内没有出现一年生草本,多年生草本的重要值也仅为0.121。方差分析结果显示,除“多年一次”漫溢样地外,其余3个漫溢频次样地的一年生草本重要值与对照样地相比均具有显著差异(P<0.05)。特别是“多年多次”漫溢样地与对照样地之间的差异达到极显著水平(P<0.01),且此样地中一年生草本重要值为5个漫溢频次样地的最大值(IV=0.415)。与对照样地相比,多年生草本重要值在4个漫溢频次样地中均表现为显著增加(P<0.05),且最大值出现在“多年一次”漫溢样地(IV=0.581)。与对照样地相比,乔灌木重要值在4个漫溢频次下均表现为极显著减少(P<0.01),其最小值出现在“多年多次”漫溢样地,为0.184。受“一年多次”频次的漫溢干扰影响,乔灌木的重要值有所减小但仍大于其他2种植被的重要值。因而乔灌木在“一年多次”漫溢样地的群落结构中占据优势。一年生草本为“多年多次”漫溢样地的主要物种。在其余2个漫溢频次样地(“多年一次”和“一年1~2次”)中,多年生草本的重要值均大于一年生草本和乔灌木的重要值,因而多年生草本是这两个漫溢频次样地的主要物种。

表2 塔里木河下游典型监测断面和样地的漫溢情况Table 2 Flooding conditions of typical monitoring sections and sample plots in the Lower Tarim River
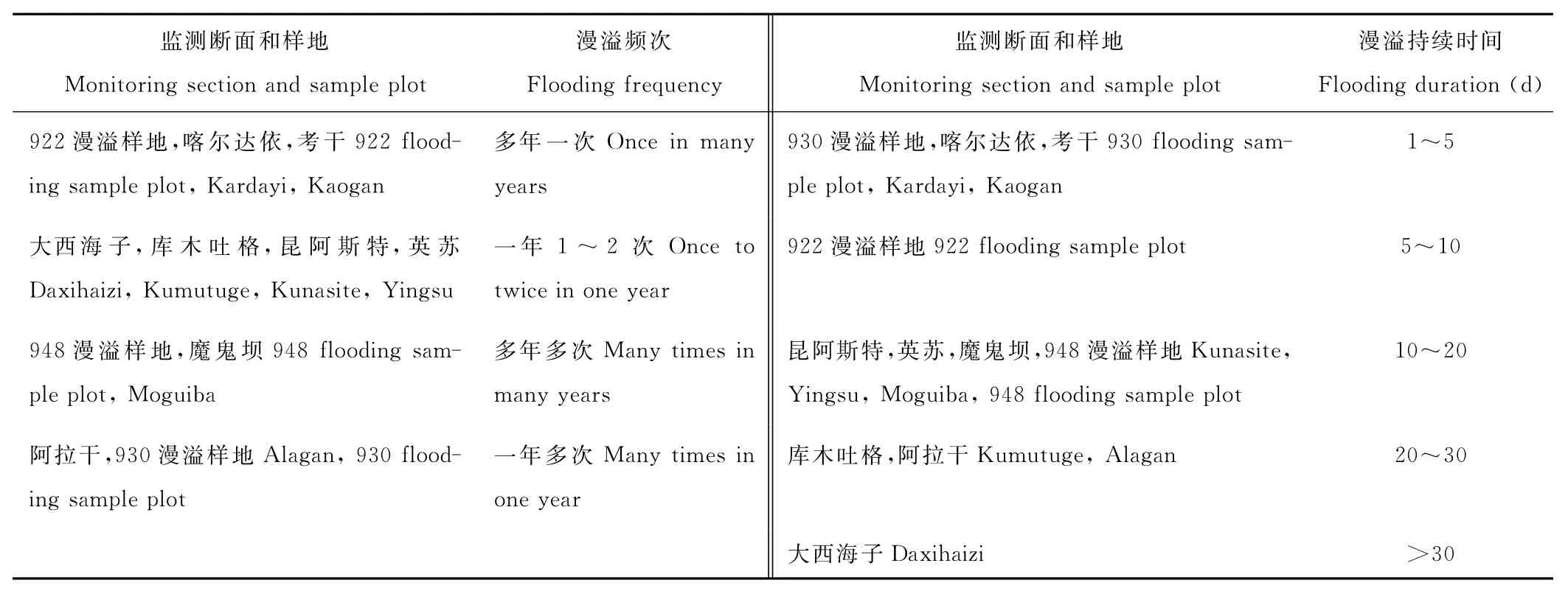
表3 塔里木河下游监测断面漫溢频次和持续时间的梯度划分Table 3 Gradient divisions of flooding frequency and duration in monitoring sections of the Lower Tarim River
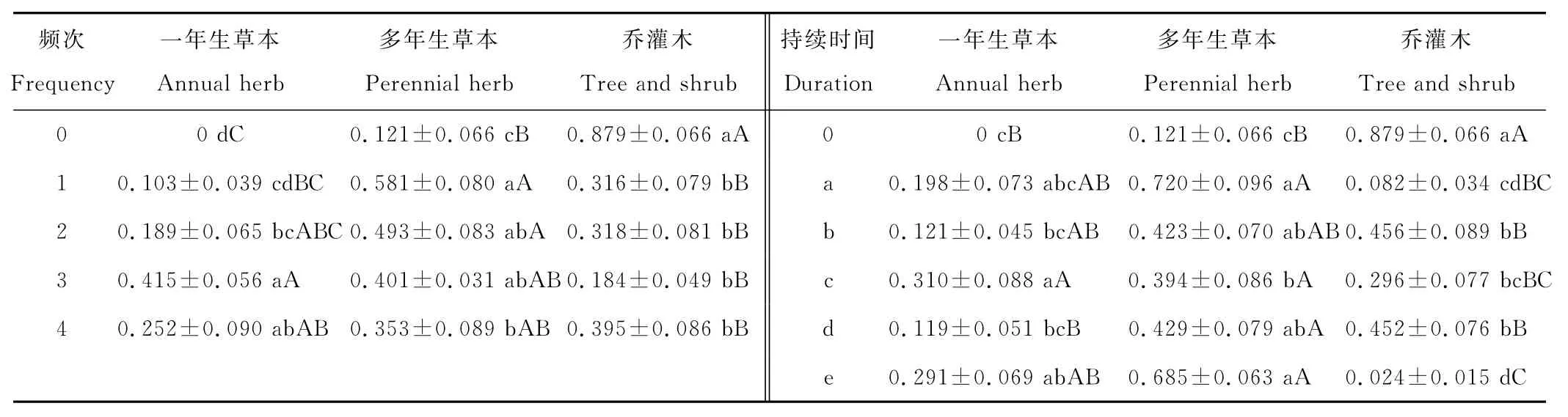
表4 不同漫溢干扰频次、不同漫溢持续时间下植被重要值(均值±SD)Table 4 Importance values of vegetation under different flooding frequencies and durations
5个持续时间梯度的漫溢干扰使得一年生草本和多年生草本的重要值均有所增加(表4)。其中一年生草本的重要值在持续时间为“10~20d”漫溢样地表现为极显著增加(P<0.01),为0.310。该漫溢样地中一年生草本的重要值位居5个漫溢持续时间样地中的首位,说明10~20d的漫溢持续时间有利于一年生草本的生长。对于多年生草本的重要值而言,除了持续时间为“5~10d”漫溢样地之外,其余4个漫溢持续时间样地与对照样地之间的差异均达到极显著水平(P<0.01),且漫溢持续时间最短的样地—“1~5d”漫溢样地中出现其最大值(IV=0.720)。此外,多年生草本的重要值在“1~5d”,“10~20d”和“>30d”漫溢样地中均排名第一位,说明多年生草本为这3个漫溢持续时间样地的主要物种。乔灌木在经历最长持续时间——“>30d”漫溢样地中分布最少,其重要值仅为0.024。而乔灌木的重要值在持续时间为“5~10d”和“20~30d”的漫溢样地中仍大于其余2个植被,因而乔灌木是这两个漫溢持续时间样地的主要物种。
2.2 漫溢干扰频次和持续时间对植被物种多样性的影响
由图2可知,漫溢频次样地与对照样地的Simpson指数之间具有显著性差异(P<0.05),除“多年一次”漫溢样地之外,其余3个漫溢频次样地与对照样地之间的差异均达到了极显著水平(P<0.01)。其中,“多年多次”漫溢样地的Simpson指数值最大(D=0.523)(图2A),其次为“一年1~2次”漫溢样地(D=0.447)。方差分析结果显示,4个漫溢频次样地的Shannon-Wiener指数与对照样地相比差异显著(P<0.05)(图2B),其中“多年多次”漫溢样地与对照样地之间的差异达到了极显著水平(P<0.01),指数值为0.979,比对照样地增加了154.02%。Margalef指数的差异情况表现为:除“多年一次”漫溢样地差异不显著外,其余3个漫溢频次样地与对照样地之间的差异均表现为极显著(P<0.01)(图2C),据多重比较结果显示,“一年1~2次”和“多年多次”漫溢样地分别与“多年一次”漫溢样地相比均具有极显著差异(P<0.01),且“多年多次”漫溢样地的Margalef指数值为1.023,是4个漫溢频次样地中的最大值。
方差结果显示,除了“1~5d”和“>30d”漫溢样地外,其余3个漫溢持续时间样地的Simpson指数与对照样地之间均具有显著性差异(P<0.05)(图3A),特别是“20~30d”漫溢样地与对照样地之间的差异达到了极显著水平(P<0.01),比对照样地增加了97.37%。同时“20~30d”漫溢样地的Simpson指数值为5个漫溢持续样地中的最大值,为0.451。就Shannon-Wiener指数而言,据方差分析结果显示,除“1~5d”和“>30d”漫溢样地外,其余漫溢持续时间样地与对照样地相比均具有显著差异(P<0.05)(图3B)。尤其是“10~20d”及“20~30 d”漫溢样地与对照样地之间的差异达到了极显著水平(P<0.01)。其中“20~30d”漫溢样地的Shannon-Wiener指数值最大(H=0.777),与对照样地相比增加了101.56%。与前2个多样性指数相同的是,除“1~5d”和“>30d”漫溢样地之外,其余3个漫溢样地的Margalef指数与对照样地之间的差异均表现显著(P<0.05),其中“10~20d”和“20~30d”漫溢样地与对照样地之间的差异达到极显著水平(P<0.01)(图3C)。Margalef指数的最大值出现在“10~20d”漫溢样地(D=0.863),比对照样地增加了162.75%。
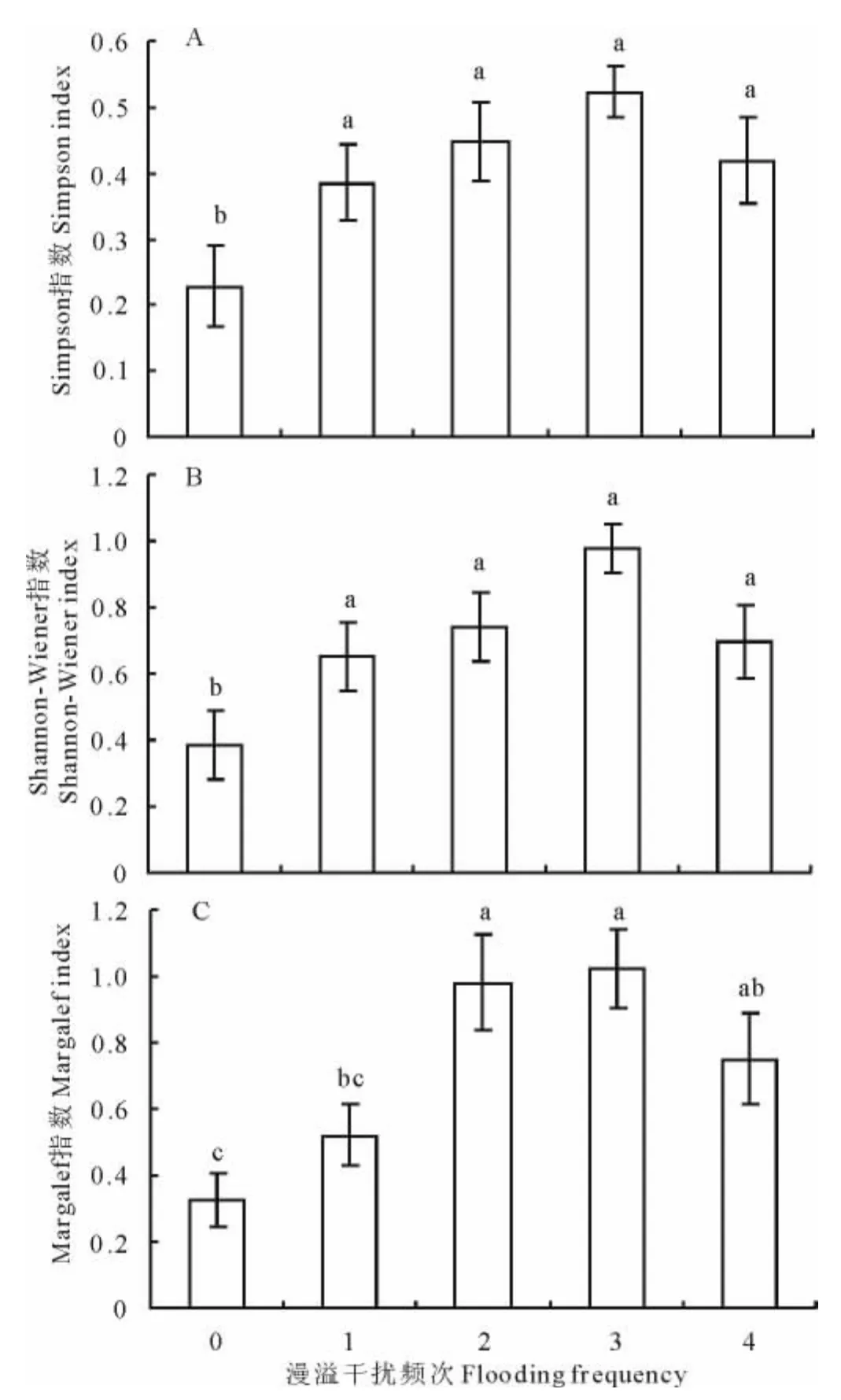
图2 不同漫溢频次下植被多样性指数差异Fig.2 Differences of diversity indexes of vegetation under different flooding frequencies
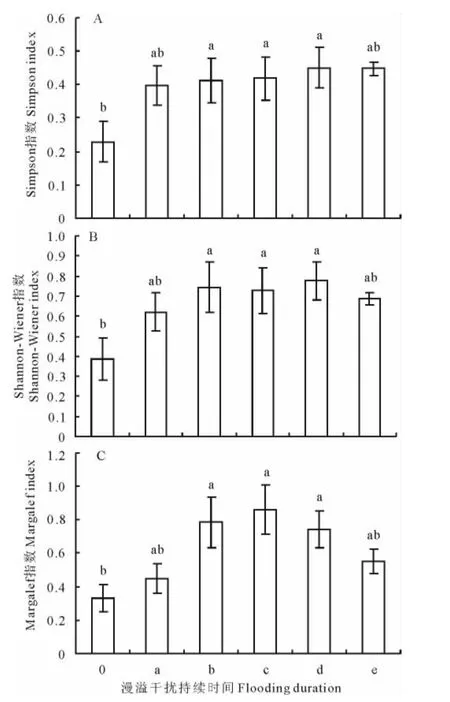
图3 不同漫溢持续时间下植被多样性指数差异Fig.3 Differences of diversity indexes of vegetation under different flooding durations
2.3 漫溢干扰频次和持续时间对土壤性质的影响
4个漫溢频次样地中的土壤电导率均小于对照样地(图4A),且在方差分析中表现为极显著差异(P<0.01)。土壤电导率大致呈“U”型变化,处于中等频次——“一年1~2次”漫溢样地的土壤电导率最小(1.148ms/cm),其次为“多年多次”漫溢样地,其土壤电导率为1.229ms/cm。在图4C中,5个漫溢持续时间样地中的土壤电导率与对照样地相比均表现为极显著减少(P<0.01),其中“10~20d”漫溢样地的土壤电导率最小,其值为1.155 ms/cm,其次为“5~10d”漫溢样地(4.325ms/cm)。频次为“多年多次”的漫溢干扰使得土壤有机质极显著增加(P<0.01),表现为4个漫溢样地中的最大值(11.577g/kg),与无漫溢样地相比增加了73.46%(图4B)。其余漫溢干扰频次样地的土壤有机质均小于无漫溢样地,且最小值出现在“一年1~2次”漫溢样地(3.818g/kg)。在漫溢持续时间样地中,较短漫溢时间——“1~5d”和“5~10d”的漫溢干扰下的土壤有机质与无漫溢条件相比有所增加,其中“5~10d”漫溢样地与对照样地之间的差异达到显著水平(P<0.05),其值为8.770g/kg,比对照样地增加了69.55%(图4D)。而其余3个漫溢样地内土壤有机质均小于对照样地,最小值出现在“20~30d”漫溢样地,其值为4.079g/kg。

图4 不同频次和持续时间的漫溢干扰下土壤特征变化Fig.4 Variations of soil characteristics after flooding with different frequencies and durations
3 讨论
漫溢干扰使得植物群落结构和演替发生明显变化。本研究在野外调查中发现,无漫溢区内长期缺水无幼苗更新,以胡杨、柽柳、黑刺(Lyciumruthenicum)等乔灌木的成年植株为主,一些退化严重的地区甚至只有枯枝。而漫溢区样地内明显有幼苗萌发,特别是一年生草本和多年生草本的幼苗萌发,说明漫溢干扰带来的湿润条件有利于浅根系植物的萌发。在不同频次和持续时间的漫溢样地中,一年生草本和多年生草本的重要值呈波动变化,而在漫溢干扰持续时间最长(30d以上)的地区,则以耐涝的多年生植物(如芦苇)占据优势。本研究得出漫溢频次中的 “多年多次”以及漫溢持续时间中的“10~20d”和“20~30d”对研究区植被物种多样性指数的促进作用最为显著,与一些学者提出的“中度干扰假说(Intermediate Disturbance Hypothesis)”理论[19-20]较为一致。王正文等[8]在松嫩平原的研究,及Barrett等[9]在澳大利亚墨累河的研究等均充分证实了这一假说。此外,Ward等[21]的研究认为,保持漫溢干扰(本研究主要指频次和持续时间)的多样性也是一种使得植物群落物种多样性最大化的方法。由于不同植被物种适应于不同强度的漫溢干扰,多样化的漫溢干扰有利于不同植被物种的生存与繁殖。
漫溢干扰可通过改变土壤性质从而影响植物的存活和分布。有研究表明,短期洪水冲刷会降低表层土壤的盐分累积,促使土壤中的有机碳、总磷和氮含量增加[22-24];但长期漫溢会导致土壤处于缺氧条件,厌氧细菌活动活跃,从而土壤中磷、氮等含量减少,植被生长受到胁迫[25]。本研究得出中等频次(“多年多次”),短期(“5~10d”)的漫溢干扰相对更有利于土壤表层盐分降低和有机质积累,与前人研究结果较为符合。至于在本研究结果中,促进植物多样性的最佳漫溢持续时间与维持土壤性质的最佳漫溢持续时间较为一致,但在研究中还应考虑其他环境因素(例如地势)的影响作用。随着野外观测数据的不断累积,后期研究可对此进行深入分析探讨。
漫溢干扰为干旱区半干旱区的荒漠植被带来水分条件,是影响荒漠植被萌发和繁殖的重要环境因子。生态输水工程的实施使得塔里木河下游出现大片漫溢区,是开展干旱区漫溢干扰研究的典型区域。基于本研究的初步分析成果,可认为今后在下游地区进行人工生态放水的频次和持续时间不宜过多过长,长期频繁的漫溢条件并不是适宜于荒漠河岸植被萌发和生长的最佳条件。综合分析认为,频次为多年多次且持续时间在10~30d的漫溢干扰对下游植被恢复最为有利。此外,鉴于下游不同断面的生态退化程度不尽相同,输水频次和持续时间也需根据不同断面植被实际生长情况而定。总之,今后研究还需在长期连续野外观测的基础上对于塔里木河下游最适宜漫溢频次和持续时间进行更深入的研究,以便科学定量地指导下游河流管理和植被恢复工作。
4 结论
乔灌木为对照样地即无漫溢条件下的主要物种,而漫溢干扰后多年生草本在植物群落中的地位有了明显提升,其中多年生草本是频次为“多年一次”和“一年1~2次”漫溢样地和持续时间为“1~5d”,“10~20d”和“>30 d”漫溢样地的主要物种。说明漫溢干扰使得植物群落结构组成发生了改变,多年生草本逐渐替代乔灌木成为漫溢样地内的主要物种。
漫溢干扰使得植被多样性指数显著增加。其中频次为“多年多次”的漫溢样地及持续时间为“10~20d”和“20~30d”漫溢样地内的多样性指数与对照样地之间的差异均达到极显著水平(P<0.01),说明连续多年多次实施且持续时间在10~30d之间的漫溢条件对植被多样性恢复较为有利。
漫溢干扰使得表层土壤电导率有所减少,最小值出现在频次为“多年多次”漫溢样地和持续时间为“10~20 d”漫溢样地。漫溢干扰在“多年多次”的频次梯度下和“5~10d”的持续时间梯度下能够显著促进表层土壤有机质的积累。说明多年多次且较短持续时间的漫溢条件有助于土壤盐分减少和有机质的积累,为植物恢复生长和新物种萌发提供了良好的条件。
[1] Mcfarlane N M,Ciavarella T A,Smith K F.The effects of waterlogging on growth,photosynthesis and biomass allocation in perennial ryegrass(LoliumperenneL.)genotypes with contrasting root development[J].Journal of Agricultural Science,2003,141:241-248.
[2] Tiegs S D,O’Leary J F,Pohl M M,etal.Flood disturbance and riparian species diversity on the Colorado River Delta[J].Biodiversity and Conservation,2005,14:1175-1194.
[3] Marks J C,Power M E,Parker M S.Flood disturbance,algal productivity,and interannual variation in food chain length[J].OIKOS,2000,90:20-27.
[4] Benke A C,Chaubey I,Ward G M,etal.Flood pulse dynamics of an unregulated river-floodplain in the southeastern U.S.coastal plain[J].Ecology,2000,81(10):2730-2741.
[5] Bischoff A.Dispersal and establishment of floodplain grassland species as limiting factors in restoration[J].Biological Conservation,2002,104:25-33.
[6] Auble G T,Shafroth P B,Scott M L,etal.Early vegetation development on an exposed reservoir:implications for dam removal[J].Environmental Management,2007,39(6):806-818.
[7] Van Etten E J B,Vellekoop S E.Response of fringing vegetation to flooding and discharge of hypersaline water at Lake Austin,Western Australia[J].Hydrobiologia,2009,626:67-77.
[8] 王正文,邢福,祝廷成,等.松嫩平原羊草草地植物功能群组成及多样性特征对水淹干扰的响应[J].植物生态学报,2002,26(6):708-716.
[9] Barrett R,Nielsen D L,Croome R.Associations between the plant communities of floodplain wetlands,water regime and wetland type[J].River Research and Applications,2010,26(7):866-876.
[10] 王晓荣,程瑞梅,封晓辉,等.三峡库区消落带回水区水淹初期土壤种子库特征[J].应用生态学报,2009,20(12):2891-2897.
[11] Casanova M T,Brock M A.How do depth,duration and frequency of flooding influence the establishment of wetland plant[J].Plant Ecology,2000,147:237-250.
[12] Blom C W P M,Voesenek L A C J.Flooding:the survival strategies of plants[J].Tree,1996,11(7):290-295.
[13] Orr C H,Stanley E H.Vegetation development and restoration potential of drained reservoirs following dam removal in Wisconsin[J].River Research and Applications,2006,22(3):281-295.
[14] 李卫红,周洪华,杨晓明,等.干旱荒漠区草地植物群落地上生物量时空分布对地下水的响应[J].草业学报,2010,19(5):186-195.
[15] 陈永金,陈亚宁,刘加珍.塔里木河下游植被覆盖度变化与地下水质关系[J].环境科学,2010,31(3):612-617.
[16] 马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数[J].生态学报,1995,15(3):268-277.
[17] 王增如,杨国靖,何晓波,等.物种多样性和生物量分布规律对冻土活动层埋深变化的响应[J].草业学报,2012,21(1):10-17.
[18] 鲁庆彬,游卫云,赵昌杰,等.旅游干扰对青山湖风景区植物多样性的影响[J].应用生态学报,2011,22(2):295-302.
[19] Connell J H.Diversity in tropical rain forests and coral reefs[J].Science,1978,199:1302-1310.
[20] Huston M A.A general hypothesis of species diversity[J].American Naturalist,1979,113:81-101.
[21] Ward J V,Tockner K,Schiemer F.Biodiversity of floodplain river ecosystems:ecotones and connectivity[J].Regulated Rivers:Research & Management,1999,15:125-139.
[22] 王正文,祝廷成.水淹干扰对羊草草地地上生物量影响的初步研究[J].应用生态学报,2003,14(12):2162-2166.
[23] Barrett-Lennard E G.The interaction between waterlogging and salinity in higher plants:causes,consequences and implications[J].Plant and Soil,2003,253:35-54.
[24] 周斌,杨红梅,胡顺军,等.河水漫溢对塔里木河下游土壤及植被的影响[J].干旱区地理,2010,33(3):442-448.
[25] Vercoutere B,Honnay O,Hermy M.Vegetation response after restoring the connectivity between a river channel and its floodplain[J].Applied Vegetation Science,2007,10:271-278.
猜你喜欢
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
意林绘阅读(2019年2期)2019-03-05
下一代英才(2018年12期)2018-02-20
现代园艺(2017年13期)2018-01-19
第二课堂(课外活动版)(2015年5期)2015-10-21
电测与仪表(2015年12期)2015-04-09
小说月刊(2014年1期)2014-04-23
吐鲁番(2014年2期)2014-02-28
城市道桥与防洪(2014年1期)2014-02-27
