
杜仲绿原酸生物合成途径相关基因的差异表达
2013-04-03 09:16李铁柱杜红岩朱高浦
经济林研究 2013年4期
李铁柱,杜红岩,朱高浦
(1.中国林业科学研究院 经济林研究开发中心,河南 郑州 450003;2.国家林业局 杜仲工程技术研究中心,河南 郑州 450003)
杜仲Eucommia ulmoidesOliv.是我国特有名贵药用树种[1],果实、叶片、树皮、雄蕊中含有苯丙素类、不饱和脂肪酸、黄酮类、环烯醚萜类、木质素类等活性成分,具有明显的补肝肾、强筋骨、安胎作用、抗菌消炎、降血压降血脂等作用,广泛应用于医药保健、食品和化妆品行业[2]。杜仲叶片中的主要苯丙素类物质是绿原酸(Chlorogenic acid)[3-7]。杜仲绿原酸抗菌作用较强,具有降压、抗菌、消炎、止血、抗氧化等作用,对急性咽喉炎及皮肤病有明显疗效[8]。
绿原酸(Chlorogenic acid)是由咖啡酸(Caffeic acid)与奎尼酸(Quinic acid,QA)组成的缩酚酸[9-10],异名咖啡单宁酸,又咖啡鞣酸,化学名为5-O-咖啡酰奎尼酸(5-O-caffeoylqulnic acid),分子式为C16H18O9,相对分子质量为354.30。它是植物在有氧呼吸过程中经磷酸戊糖途径中间产物合成的一种苯丙素类物质[11]。绿原酸生物合成途径见图1。

图1 绿原酸合成途径Fig.1 The pathway of chlorogenic acid biosynthesis
咖啡酰基在奎尼酸上的结合部位和数目有所不同,从理论上讲,单咖啡酰奎尼酸和二咖啡酰奎尼酸所行形成的绿原酸共有10种异构体,但到目前为止,从植物中发现的绿原酸异构体有7种,即绿原酸(5-O-咖啡酰奎尼酸)、隐绿原酸(4-咖啡酰奎尼酸)、新绿原酸(3-咖啡酰奎尼酸)、异绿原酸A(4,5-二咖啡酰奎尼酸)、异绿原酸B(3,5-二咖啡酰奎尼酸)、异绿原酸C(3,4-二咖啡酰奎尼酸)、莱蓟素(l,3-二咖啡酰奎尼酸)。
1 材料与方法
分别于2010年5月28日和10月28日,在国家林业局泡桐研究开发中心采集“华仲6号”杜仲优良品种叶片和果实,提取RNA,送至深圳华大基因公司进行转录组测序和基因功能注释[12-13]。
转录组测序得到的Unigene,利用NCBI网站上的BLAST工具,和其它物种已知的CDS序列进行在线比较,用相似性得分比较高的已知基因确定并命名杜仲转录组中的待分析基因。确定基因后,分析该基因(或基因家族)在杜仲叶片和果实中的表达量,推断该基因在杜仲活性成分生物合成中的地位和作用。
2 结果与分析
2.1 杜仲转录组中绿原酸生物合成途径中被注释的Unigene基因
在杜仲叶片和果实转录组数据中共3个步骤、4个基因被KEGG数据库绿原酸合成途径注释(见图2)。苯丙氨酸脱氨酶(phenylalanin ammonia-lyase,PAL)注释Unigene9151等3条Unigene,其中果实和叶片差异表达的基因有1条;肉桂酸-4-羟化酶(cinnamate 4-hydroxylase,C4H)注释Unigene9197等9条Unigene,其中差异表达基因有4条;香豆酸-CoA连接酶(4CL)注释Unigene6686等24条Unigene,其中差异表达基因有13条;莽草酸羟基肉桂酰转移酶(shikimate hydroxycinnamoyl transferase,HCT) 注 释Unigene24848等43条Unigene,其中差异表达基因有20条。
2.2 杜仲叶片和果实绿原酸合成途径系列基因的确定
经过进一步BLAST分析发现,Unigene9151、Unigene3306、Unigene8123等13条Unigene与长春花、咖啡等植物种的PAL基因相似性达到77%~99%(见表1)。其中Unigene9151为PAL基因全长cDNA;Unigene9197、Unigene27913等2条Unigene与烟草等植物种的肉桂酸-4-羟化酶相似性达到77%~90%,为C4H基因全长cDNA;Unigene6686、Unigene14128等11条Unigene与杨树、白桦等植物种的4CL基因相似性达到66%~92%;Unigene24848、Unigene6511与咖啡等植物种的HCT基因相似性分别达到56%和54%,其中Unigene24848为全长cDNA。
2.3 杜仲绿原酸相关基因多样性及表达差异
2.3.1 杜仲EuPAL基因的多样性及表达差异
苯丙氨酸脱氢酶(PAL)是绿原酸合成途径中的关键酶[14-15],杜仲的果实和叶片组织中PAL的多态性非常高,PAL基因家族高达至少12个成员,分别命名为EuPAL1~EuPAL12,其中Unigene9151即EuPAL1为全长cDNA(见表2)。
杜仲果实和叶片的转录组分析表明,杜仲苯丙氨酸脱氢酶EuPAL1~EuPAL12(Unigene15537~Unigene81458)12个基因在叶片和果实中均有表达,但EuPAL3~EuPAL12(Unigene15537~Unigene8123)的表达量存在显著差异(见表3)。其中表达量最高的EuPAL3(Unigene46841)和 最 低 的EuPAL12(Unigene38418)在果实的表达量分别比叶片高29.22倍和19.02倍(见图3),而EuPAL1(Unigene9151) 和EuPAL12(Unigene18458),在果实和叶片的表达量未达到显著水平。

图2 KEGG数据库中的绿原酸生物合成Fig.2 The KEGG pathway of chlorogenic acid biosynthesis

表1 绿原酸合成途径中相关酶基因和对应Unigene相似度信息Table 1 Information of similarity between gene and Unigene in chlorogenic acid synthesis pathway

表2 杜仲叶片和果实EuPAL基因多样性Table 2 Genetic diversity of EuPAL
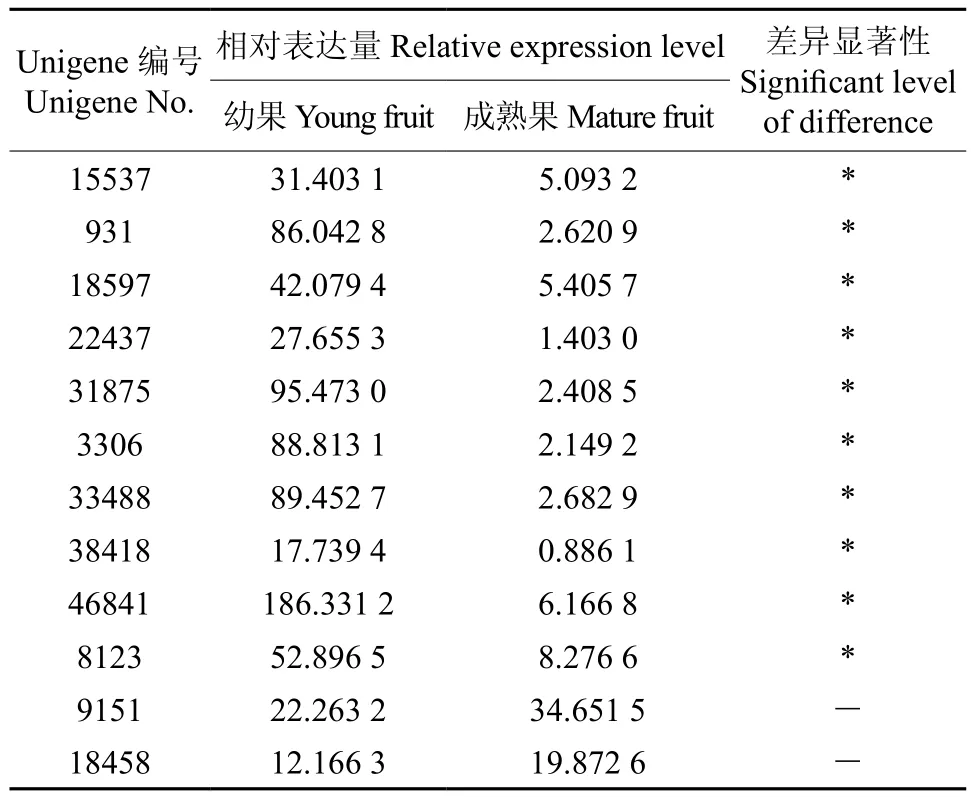
表3 苯丙氨酸脱氢酶相关基因在杜仲叶片和果实中的表达多样性Table 3 Expression of Unigene related in EuPAL in leaves and young fruits
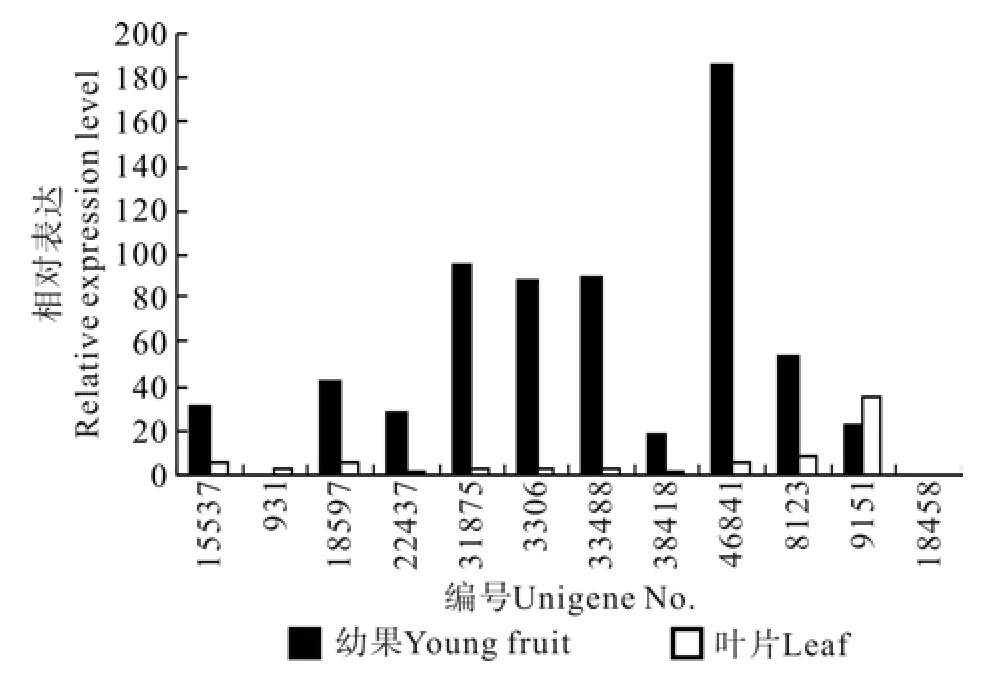
图3 苯丙氨酸脱氢酶相关Unigene在杜仲叶片和果实中的表达差异Fig.3 Expression difference of related EuPAL Unigene related in leaf and fruit
叶片的PAL基因的表达量最高为EuPAL1(Unigene9151),其次为EuPAL12(Unigene81458),推测杜仲叶片中绿原酸合成的主要的苯丙氨酸脱氢酶为EuPAL1和EuPAL2。果实PAL基因按照表达量由大到小的顺序排列依次为EuPAL3、EuPAL4、EuPAL5、EuPAL6、EuPAL7、EuPAL8、EuPAL9、EuPAL10、EuPAL11、EuPAL1、EuPAL12、EuPAL2。杜仲叶片以EuPAL1和EuPAL2为主,而果实中PAL基因的遗传多样性比叶子高很多,基因表达量也高很多,表达调控模式极其复杂。
2.3.2 EuC4H基因的多样性和表达差异
肉桂酸-4-羟化酶(C4H)基因拷贝数在不同植物种是不同的,如苜蓿有2个,豌豆有1个,绿豆和长春花中多个拷贝的小基因家族形式存在[15-17]。从表4可以看出,杜仲的果实和叶片组织中获得2个全长C4H基因(Unigene9197和Unigene27913),分别命名为EuC4H1和EuC4H2。

表4 杜仲肉桂酸-4-羟化酶基因的多样性Table 4 Diversity of EuC4H genes
欧芹种子C4H表达模式研究结果显示,该酶在微管束发达的花梗中表达丰富,在幼叶和老叶中不表达,拟南芥种C4H基因于正在木质化的细胞中表达最强[18-19]。杜仲的果实和叶片中EuC4H1(Unigene9197)和EuC4H2(Unigene27913)基因均有表达,且表达量存在着显著差异(见表5),EuC4H1和EuC4H2在果实中的表达量分别比叶片高出105.25倍和6.32倍(见图4)。叶片中EuC4H1的表达量较高,但EuC4H2表达量接近零,推测叶片绿原酸合成途径中主要起作用的是EuC4H1基因。无论在果实还是在叶片中EuC4H1的表达量均比EuC4H2高,故果实也以EuC4H1为主。
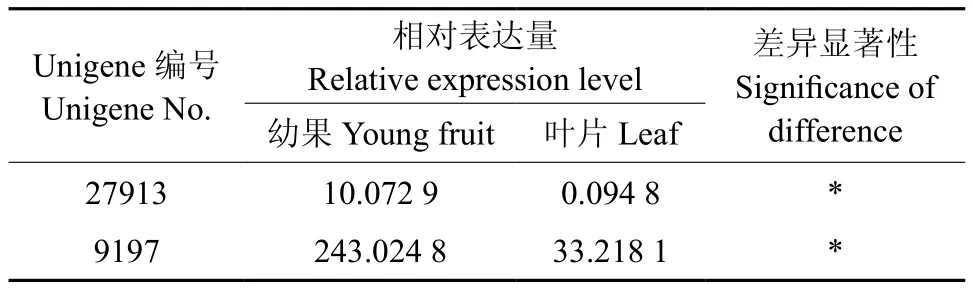
表5 EuC4H基因在杜仲叶片和果实中的表达多样性Table 5 Expression diversity of EuC4H in leaf and fruit

图4 EuC4H基因在杜仲叶片和果实中的表达差异Fig.4 Expression difference of EuC4H in leaf and fruit
2.3.3 Eu4CL基因的多样性及表达差异
拟南芥中4CL基因家族有4个成员,烟草有2个成员,大豆有4个成员,颤杨有2个成员,复盆子有3个成员[20-22]。杜仲的果实和叶片中获得了11个4CL家族成员,分别命名为Eu4CL1~Eu4CL11(见表6)。
11个杜仲香豆酰-CoA连接酶中Eu4CL2(Unigene14857)、Eu4CL3(Unigene6686)、Eu4CL7(Unigene18613)等3个Eu4CL基因在果实和叶片的表达未达到显著差异(见表7);而Eu4CL1(Unigene14128)、Eu4CL4(Unigene48688)、Eu4CL5(Unigene29944)、Eu4CL6(Unigene45667)、Eu4CL8(Unigene28080) 等 5个Eu4CL在 果 实和叶片中的表达具有显著差异,而且在幼果中均比叶片中表达量高,表达量最大的Eu4CL4基因比叶片高出35.04倍(见表7和图5);Eu4CL9(Unigene40135)、Eu4CL10(Unigene23597)、Eu4CL11(Unigene22350)在果实中特异表达,但表达量较低。按照Eu4CL在果实中表达量由大到小的顺序排列依次为:Eu4CL4、Eu4CL6、Eu4CL5、Eu4CL1、Eu4CL2、Eu4CL3、Eu4CL7、Eu4CL8、Eu4CL11、Eu4CL9、Eu4CL10; 按 照Eu4CL在叶片中表达量由大到小的顺序排列依次 为:Eu4CL2、Eu4CL3、Eu4CL6、Eu4CL4、Eu4CL5、Eu4CL1、Eu4CL7、Eu4CL8。幼果和叶片中Eu4CL~Eu4CL6等基因大量表达,表达调控规律极其复杂。

表6 杜仲4CL基因多样性Table 6 Diversity of Eu4CL genes

表7 香豆酰-CoA连接酶相关Unigene在杜仲叶片和果实中的表达多样性Table 7 Expression diversity of related Eu4CL Unigene in leaf and fruit

图5 香豆酰-CoA连接酶相关Unigene在杜仲叶片和果实中的表达差异Fig.5 Expression difference of related Eu4CL Unigene in leaf and fruit
2.3.4 EuHCT基因的多样性和表达差异
莽草酸羟基肉桂酸酰转移酶是绿原酸合成途径中的限速酶[23-24]。杜仲转录组数据中获得了3个莽草酸羟基肉桂酸酰转移酶基因家族3个成员,分别命名为EuHCT1~EuHCT3(见表8)。
3个EuHCT基因家族成员在叶片和果实中均有表达,但仅有EuHCT1在果实和叶片中表达量有显著差异,且在果实中的表达量比叶片高出25.44倍(见表9和图6);而EuHCT2和EuHCT3在果实和叶片中的表达量无显著差异。按照在果实中表达量由大到小的顺序排列依次为EuHCT1、EuHCT2、EuHCT3; 按 照 叶 片 中的表达量由大到小的顺序排列依次为EuHCT2、EuHCT1、EuHCT3。杜仲幼果以EuHCT1、叶片以EuHCT2为主。

表8 杜仲莽草酸羟基肉桂酸酰转移酶基因多样性Table 8 Diversity of EuHCT genes

表9 莽草酸羟基肉桂酸酰转移酶相关Unigene在杜仲叶片和果实中的表达量Table 9 Expression of related HCT Unigene in leaf and fruit
3 结论与讨论
杜仲苯丙素类物质广泛存在于杜仲根皮、茎皮、绿叶、果实中,包括绿原酸、咖啡酸、松柏酸、松柏苷、丁香苷、香草酸等11种化合物。本研究中发现的各相关基因及基因家族,其中哪些基因家族成员专营合成绿原酸,或者哪些基因在绿原酸合成中起最关键作用,这些问题都有待进一步分析。
苯丙氨酸解氨酶是杜仲绿原酸合成途径中的上游的关键酶,本研究从杜仲叶片和果实的转录组中发现了12个PAL基因,说明杜仲该基因的遗传多样性非常高,超过一般植物。杜仲器官组织中的绿原酸含量,还随着时间季节的变化而变化[25],本研究采集的样品是5月份和10月份的,杜仲PAL基因及其它基因是否也有时间上的多态性。
另外,在不同的杜仲品种之间,绿原酸等活性成分含量也不一样[26],在其它品种中,PAL等基因的表达规律是否和“华仲6号”一致还有待进一步探讨。不同栽培模式对杜仲叶片及枝、皮等器官中的活性成分的含量也有较大影响[27],从分子层次来看,是否对PAL等基因的表达调控也会造成影响,还需要进一步分析。
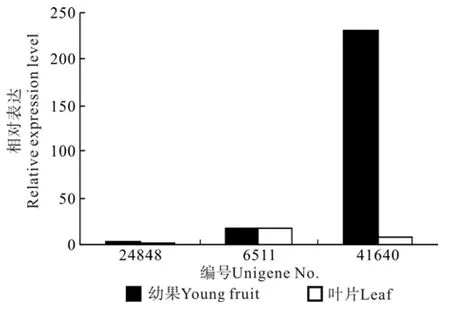
图6 HCT相关Unigene在杜仲叶片和果实中的表达差异Fig.6 Expression difference of related HCT Unigene in leaf and fruit
参考文献:
[1] 李时珍.本草纲目[M].北京:北京出版社,1995:1590-1596.
[2] 薛程远,曲范仙,刘 辉,等.杜仲叶乙醇提取物对小鼠免疫功能的影响[J].甘肃中医学院学报,1998,15(3):50-52.
[3] 吴龙奇,朱文学,张玉先,等.杜仲中绿原酸含量及提取检测方法的分析[J].食品科学,2005,(S1):187-192.
[4] 陈乃炽,汪洪武,刘艳清,等.杜仲叶中绿原酸的提取与含量测定[J].经济林研究,2001,19(2):59-61.
[5] 刘昌勇,李 钦,杜红岩,等.配方施肥对杜仲雄花主要活性成分含量的影响[J].中南林业科技大学学报,2010,30(8):29-34.
[6] 涂 佳,欧阳昶,吴建平,等.金银花氨基酸有机无机专用肥的增产示范研究[J].中南林业科技大学学报, 2012, 32(12):191-194.
[7] 马晓乾,刘 欣,邓 勋,等.根部损伤及外源物质对红皮云杉幼苗酚酸含量的影响[J].中南林业科技大学学报,2013,33(3): 51-56.
[8] 杜红岩.杜仲活性成分与药理研究的新进展[J].经济林研究,2003,21(2):58-61.
[9] 彭新辉,易建华,周清明,等.烟草绿原酸的研究进展[J].中国烟草学报,2006,(4):52-57.
[10] 邓 良,袁 华,喻宗沅.绿原酸的研究进展[J].化学与生物工程,2005,(7):4-7.
[11] Amrita Kumar, Brian E Ellis.The Phenylalanie Ammonia-Lyase Gene Family in Raspberry.Structure, Expession, and Evolution[J].Plant Physiology, 2001,127:230-239.
[12] 李铁柱,杜红岩,刘慧敏,等.杜仲果实和叶片转录组数据组装及基因功能注释[J].中南林业科技大学学报,2012,32(11):122-130.
[13] 李铁柱,杜红岩,刘慧敏,等.杜仲幼果和成熟果实转录组数据组装及基因功能注释[J].中南林业科技大学学报,2012,32(10):9-17.
[14] Koukol J, Coan E E.The metabolism of aromatic compounds in higher Plants IV Puri fi cation and properties of the phenylaianine deaminase of Herdeum vulagare [J].Journal of Biology and Chemistry,1961,236(10):2692-2698.
[15] Whetten R W, Sederof R R.Phenylalanine Ammonia-Lyase from Loblolly Pine(Puri fi cation of the EnZyme and Isolation of Complementary DNA Clones[J].Plant Physiol, 1992, 98:380-386.
[16] Bell-Lelong D A, Cusumano J C, Meyer K,et al.Cinnamate-4-hydroxylase expression inArabidopsis(Regulation in response to development and the environment)[J].Plant physiol, 2007,113(6): 729-738.
[17] Chapple C.Molecular-genetican alysis oplant cytochromep450-depen dentmono oxygenases[J].Annu Rev plant physiol Plant Mol Biol,1998,49(5):311-343.
[18] Koopman E, Logemann E, Hahlbrock K.Regulation and functional expression of cinnamate 4-hydroxylase from parsley [J].plant physiol, 1999,119(11):49-55.
[19] Nedelkina S, Jupe S C, Blee K,et al.Novel characteristics and regulation of a divergent cinnamate 4-hydroxylase (CYP73A15)from French bean: engineering expression in yeast[J].Plant Mol Biol,1999,39:1079-1090.
[20] 杨 婷,潘 翔,饶国栋,等.植物4CL基因家族结构功能与表达特性研究进展[J].成都大学学报:自然科学版,2011, (1):4-7.
[21] 毕咏梅,欧阳光察.水稻4-香豆酸CoA连接酶的基本性质[J].植物生理学通讯,1990,(6):18-20.
[22] 李欢欢,饶国栋,范丙友,等.重组毛白杨4-香豆酸:辅酶A连接酶催化不同肉桂酸衍生物的酶促动力学研究[J].成都大学学报:自然科学版,2009,(1):1-4.
[23] 薛永常,李金花,卢孟柱,等.木质素单体生物合成途径及其修订[J].林业科学,2003,39(6):146-153.
[24] 耿 飒,徐存拴,李玉昌.木质素的生物合成及其调控研究进展[J].西北植物学报, 2003, 23(1): 171-181.
[25] 杜红岩,刘昌勇,李 钦,等.杜仲叶中3种主要活性成分含量的季节变化[J].中南林业科技大学学报,2011,31(8):6-9.
[26] 杜红岩,李 钦,刘攀峰,等.不同变异类型杜仲皮及再生皮中木脂素类成分含量比较[J].中南林业科技大学学报,2010,30(7): 38-42.
[27] 吕 强,彭密军,彭 胜,等.不同栽培模式对杜仲叶及枝皮中多种活性成分含量的影响[J].经济林研究,2012,30(1):73-76.
猜你喜欢
世界科学技术-中医药现代化(2021年10期)2021-03-02
天然产物研究与开发(2019年1期)2019-03-01
中成药(2018年11期)2018-11-24
中成药(2018年8期)2018-08-29
中成药(2017年10期)2017-11-16
材料科学与工程学报(2016年4期)2017-01-15
陕西画报(2016年1期)2016-12-01
小雪花·小学生快乐作文(2015年9期)2015-09-29
小雪花·小学生快乐作文(2015年9期)2015-09-29
华东理工大学学报(自然科学版)(2014年1期)2014-02-27
