
青藏高原高山嵩草草甸优势植物营养成分对放牧的响应
2013-03-14 01:50:26付娟娟益西措姆苗彦军呼天明许岳飞
草业科学 2013年4期
付娟娟,益西措姆,陈 浩,苗彦军,呼天明,许岳飞
(1.西北农林科技大学动物科技学院,陕西 杨凌 712100; 2.西藏农牧学院植物科学学院,西藏 林芝 860000)
高寒草甸作为草地生态系统的重要组成部分,其独特的气候环境使得生态系统非常脆弱,且环境异质性很大[1]。我国高寒草甸主要分布在青藏高原地区,可利用面积为5.88×104hm2,居于各类草地面积之首。高寒草甸具有多种功能和多重效益,不仅是农牧民的基本生产资料,更是重要的生态屏障[2-3]。最近研究发现,西藏的高寒草甸退化严重,草地载畜量是影响草地退化的主要因素之一[3-4],因此,研究西藏高寒草甸野生牧草的营养价值进而确定合理载畜量迫在眉睫。
牧草的营养成分是形成畜产品的基础,也是评定牧草营养价值的重要指标之一。牧草在不同生产阶段或季节的营养成分变化,会直接影响到家畜的健康及生产性能[5]。评定牧草的营养物质含量时,多以牧草的粗蛋白质、粗脂肪、矿物质及粗纤维素含量等为主要指标。粗蛋白质和矿物质含量越高,纤维素含量越低,牧草营养价值就越高,反之,牧草营养价值就越低[6]。
放牧家畜通过采食植物吸收养分来满足自身生长的需要,家畜通过排泄使一部分营养物质和矿物质又回归到土壤,草地植物又可以吸收利用这些养分来满足自身的生长繁殖。放牧家畜的选择性采食、践踏等行为,不仅会改变草地植物的群落结构,同时对植物的营养结构及养分含量也会产生影响[7-9]。然而,不同放牧强度对草地植物养分变化的影响不同,而植物自身也表现出不同的适应机制。牧草贮藏养分及其变化趋势是确定草地放牧利用时间的最佳指标[10-11]。许多优质牧草从草地上消失,主要是因为利用过度引起的养分耗竭所致[12-13],因此,本研究对青藏高原放牧区主要野生牧草的营养成分变化进行了分析和总结,旨在探明西藏高寒草甸的最佳放牧方式,达到草地-土壤-家畜的优化配置,以便为西藏地区确定合理的草地放牧和管理措施,为草地的持续、稳定和高产提供科学的理论依据,为实现高寒草地生态系统的可持续发展提供一定的科学依据。
1 材料与方法
1.1研究区自然概况 试验地位于西藏林芝地区工布江达县加兴乡松多三村邦杰塘野外试验站(29°87′ N,93°38′ E),海拔4 450 m,最冷月(1月)平均气温-10.5 ℃,最热月(7月)平均气温9.3 ℃,年平均气温-3.8 ℃。雨水主要集中在夏季,年均降水量为550 mm,年均日照时数为2 016 h,无绝对无霜期,野生牧草生长期为120 d左右。草地类型为高山嵩草草甸。主要植被有高山嵩草(Kobresiapygmaea)、紫花针茅(Stipapurpurea)、西藏羊茅(Festucawallichanica)、西藏粉报春(Primulapumilio)、高山点地梅(Androsacegmelinii)、高山委陵菜(Potentillapolyschista)、高山大戟(Euphorbiastracheyi)、高山红景天(Rhodiolarosea)、高原唐松草(Thalictrumcultratum)、条裂银莲花(Anemonetrullifolia)、马先蒿(Pedicularisreaupinanta)、独一味(Lamiophlomisrotata)、肉果草(Lanceatibetica)、火绒草(Leontopodiumalpinum)等。
1.2试验方法
1.2.1试验设计 于2010年在西藏邦杰塘野外试验站建站,放牧家畜为48月龄的雌性牦牛,质量(100±5.0) kg。本试验采用经济型试验设计方法——以畜定草的方式,根据试验地地上多年平均生物量、家畜理论采食量、草地面积及放牧时间确定放牧强度,即重度放牧(HG)、适度放牧(MG)和零放牧(CK)3个处理。重度放牧小区面积为5.4 hm2,适度放牧小区面积为8.0 hm2,对照小区面积为2.0 hm2。重度与适度放牧小区均放牧3头牦牛,每个处理设置3次重复,共9个小区。完全随机排列,各小区之间用刺围栏分割,研究不同放牧强度对高寒草甸植物养分的影响。
1.2.2野外调查与取样 在每个试验小区内按照对角线选定3个具有代表性的固定样点,分别在2011年6月20日(放牧前)和8月24日(放牧后)在每个样点上随机选取5个样方(50 cm×50 cm)进行植物群落调查。
本试验以青藏高原高山嵩草草甸优势种莎草科的高山嵩草和禾本科的紫花针茅为试验样品,采集不同牧压(零放牧、适度放牧和重度放牧)下的以上两种优势植物,对其粗蛋白(CP)、粗脂肪(EE)、粗纤维(CF)、钙(Ca)、磷(P)营养成分进行测定,进而研究放牧对高山嵩草草甸优势种植物养分的影响。粗蛋白采用凯氏定氮法测定、粗脂肪采用索氏提取法测定、粗纤维使用粗纤维测定仪测定、植物全钙采用乙二胺四乙二酸钠(EDTA)络合滴定法测定、全磷采用比色法测定、干物质采用烘干法测定[4]。
2 结果与分析
2.1放牧对高山嵩草草甸优势植物粗蛋白含量的影响 不同放牧强度下,高山嵩草草甸的优势种高山嵩草粗蛋白的含量无显著差异(P>0.05),但放牧前各处理植物粗蛋白含量差异显著(P<0.05),且以适度放牧为最高;紫花针茅放牧前各处理间粗蛋白含量差异显著(P<0.05),适度放牧区粗蛋白含量比对照高43.7%,而重度放牧比对照高14.5%;放牧结束后适度放牧和重度放牧紫花针茅粗蛋白含量显著高于对照(P<0.05),分别比对照高71.0%与70.2%,而适度放牧与重度放牧差异不显著(P>0.05)。放牧能促进高山嵩草草甸优势植物高山嵩草与紫花针茅对氮的吸收,进而增加牧草蛋白质含量,改善野生牧草的品质(图1)。
2.2放牧对高山嵩草草甸优势植物粗脂肪的影响 放牧对高山嵩草与紫花针茅粗脂肪含量的影响不显著(图1)。高山嵩草放牧前与放牧结束后各处理间差异均不显著(P>0.05);紫花针茅重度放牧后粗脂肪含量显著高于对照与适度放牧(P<0.05),适度放牧与对照相比差异不显著(P>0.05)。但是,放牧前各处理间无显著差异(P>0.05)。同一处理水平下,适度放牧处理紫花针茅粗脂肪含量,放牧结束后比放牧前减少25.4%,重度放牧处理,放牧结束后比放牧前减少6.8%。放牧导致高山嵩草草甸优势植物高山嵩草与紫花针茅粗脂肪含量减少,而且这种影响在不同植物种上表现出不同的效应。
2.3放牧对高山嵩草草甸优势植物粗纤维的影响 高山嵩草和紫花针茅粗纤维含量变化在不同牧压下表现出一定的差异(图2)。放牧前高山嵩草粗纤维含量表现为,适度和重度放牧显著高于对照(P<0.05);放牧结束后,对照的植物粗纤维含量显著高于适度放牧和重度放牧(P<0.05),适度放牧显著低于重度放牧(P<0.05)。从试验结果可适度放牧和重度放牧处理高山嵩草,放牧后粗纤知,维含量比放牧前增加5.0%和8.2%。紫花针茅放牧后粗纤维含量与高山嵩草变化趋势一致,适度放牧与重度放牧处理,放牧后比放牧前分别增加4.2%与6.8%。因此,放牧可以降低高山嵩草和紫花针茅粗纤维含量,提高牧草品质。

图1 不同放牧强度下高山嵩草草甸优势植物粗蛋白和粗脂肪含量的变化Fig.1 Crude protein and fat contents of dominant plants in Kobresia pygmaea meadow responding to different grazing intensities
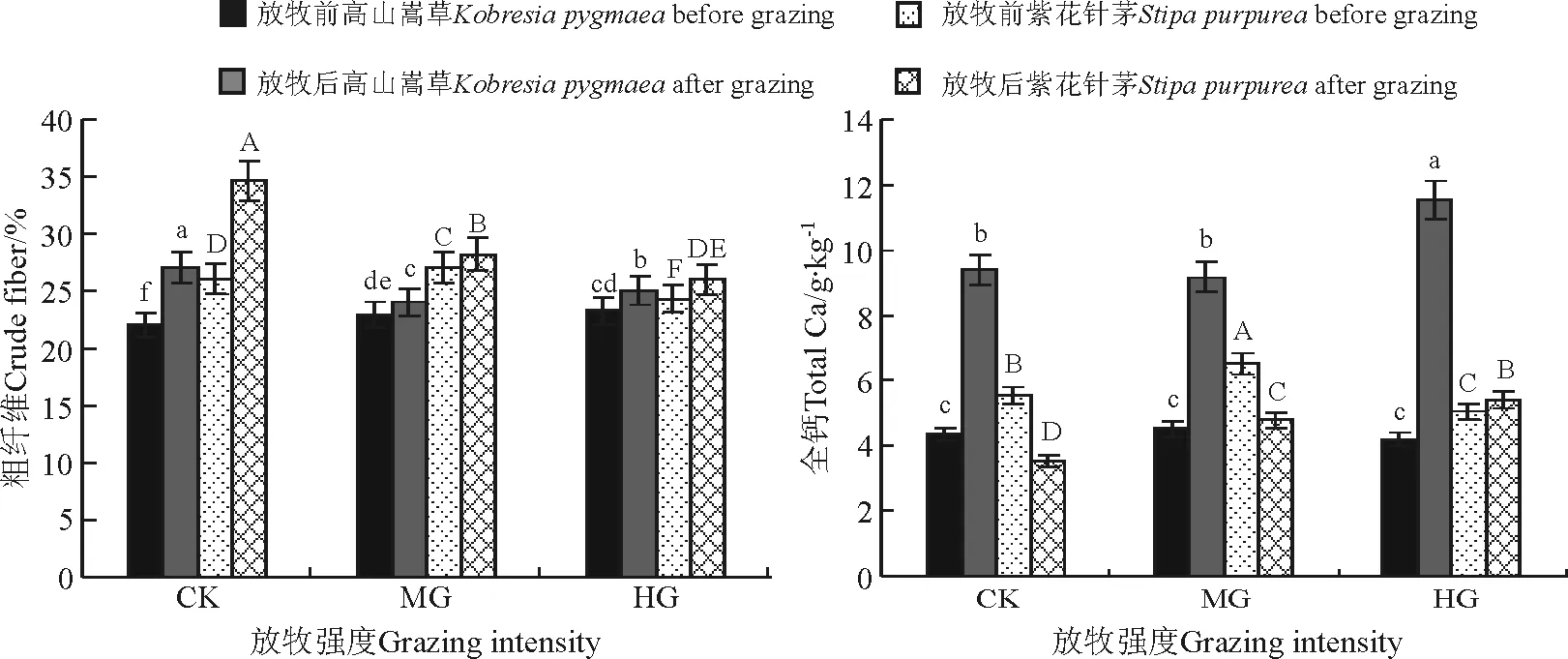
图2 不同放牧强度下高山嵩草草甸优势植物粗纤维和全钙含量的变化Fig.2 Crude fiber and total calcum contents of dominant plants in Kobresia pygmaea meadow responding to different grazing intensities
2.4放牧对高山嵩草草甸优势植物全钙含量的影响 随着放牧强度的增加,高山嵩草草甸植物全钙含量发生了显著的变化(图2)。高山嵩草放牧前不同处理间无显著差异(P>0.05);随着放牧强度的增加,放牧后的高山嵩草全钙含量显著增加(P<0.05);同一处理下,放牧前后高山嵩草全钙含量增加不同,对照、适度和重度放牧处理放牧结束后分别比放牧前分别增加116.6%、103.1%与175.4%。随着放牧强度的增加,紫花针茅放牧结束后不同处理差异显著(P<0.05),其全钙含量变化为重度放牧>适度放牧>对照。同一处理水平下,放牧前后紫花针茅全钙含量增加不同,对照与适度放牧处理,放牧结束后比放牧前分别减少了36.4%和26.7%,而重度放牧处理,放牧后比放牧前增加6.7%。以上结果表明,放牧,尤其是重度放牧有利于高山嵩草草甸优势植物高山嵩草与紫花针茅对钙的吸收。
2.5放牧对高山嵩草草甸优势植物全磷的影响 不同放牧强度下,高山嵩草放牧前不同处理全磷含量差异显著(P<0.05)。然而,放牧后对照和适度放牧处理下的高山嵩草全磷含量差异不显著(P>0.05),重度放牧显著高于对照和适度放牧。同一处理水平下,高山嵩草全磷含量与全钙含量变化相反,对照、适度放牧与重度放牧处理下放牧后分别比放牧前降低65.6%、36.0%与26.0%。紫花针茅全磷含量变化规律性较强,放牧前后各处理变化趋势基本一致,随着放牧强度的增加,出现先升高后下降的变化趋势,并且对照全磷含量显著低于放牧处理,且适度放牧全磷含量最高(图3)。
2.6放牧对高山嵩草草甸优势植物干物质含量的影响 随着放牧强度的增加,高山嵩草与紫花针茅干物质含量变化趋势基本一致(图3)。放牧前高山嵩草与紫花针茅干物质含量变化为适度放牧>重度放牧>对照,且适度放牧与重度放牧干物质含量差异不显著(P>0.05)。适度放牧后,高山嵩草的干物质含量最高,分别比重度放牧与对照增加了85.2%和106.9%。紫花针茅放牧后干物质含量变化与高山嵩草一致,适度放牧干物质含量分别比重度放牧与对照增加25.9%和61.6%。以上结果表明,放牧影响高山嵩草草甸优势植物高山嵩草与紫花针茅干物质含量的积累。
3 讨论
通过对不同放牧强度下西藏高山嵩草草甸优势植物粗蛋白、粗纤维、粗脂肪、钙和磷等含量变化的研究发现,在这5个营养指标中,只有粗纤维含量变化趋势在放牧后表现出一致性,粗纤维含量变化趋势为对照显著高于适度放牧和重度放牧。与高山嵩草不同的是,适度放牧和重度放牧紫花针茅粗纤维含量差异显著。从同一处理水平下,放牧前后结果可知,放牧能够减缓高寒草甸优势植物粗纤维含量随着生育期的推进而增加的现象,从而提高牧草品质,且适度放牧的这一效应更明显。可能是由于放牧家畜的啃食促进了植物的补偿性生长,使幼嫩枝条比例增加,故放牧后粗纤维含量显著低于对照。
粗蛋白是评价牧草营养价值的重要指标之一。研究发现,放牧前紫花针茅各处理间粗蛋白含量差异显著;放牧后,适度放牧和重度放牧粗蛋白含量显著高于对照,但适度放牧与重度放牧之间差异不显著;而高山嵩草放牧前以适度放牧粗蛋白含量最高,对照与重度放牧差异不显著,放牧后各处理间粗蛋白含量无显著差异,这与李春莉[14]、闫凯等[15]、董全民等[16]、Garcia 等[17]和江小雷等[18]的研究结果一致。通过比较分析同一处理下放牧前后高山嵩草与紫花针茅粗蛋白含量变化,发现放牧能够减缓高寒草甸优势种紫花针茅粗蛋白含量随着生育期的推进而降低的现象。可能的原因是,一定的放牧强度刺激了一些优势牧草的补偿性生长,使得放牧后牧草再生性比对照组强[19],且牧草新生组织中的氮在重度放牧和适度放牧下显著高于对照,而老组织中的氮则相反[20];其原因也可能是,放牧加速了土壤有机氮的矿化率,为植物的生长提供了充足的有效养分[21],这使放牧区内牧草氮和磷的含量高于对照[22]。
本研究还发现,紫花针茅与高山嵩草放牧前后粗脂肪的含量变化不同。粗脂肪含量不同的主要原因可能是不同牧草抽穗时间不一致,而放牧强度的大小是影响牧草抽穗的一个主要因素[18],故放牧家畜的采食促进牧草的补偿性生长,使幼嫩枝条比例上升进而使粗脂肪含量增加。但适度放牧的这种效应并不明显。对钙和磷的研究发现,紫花针茅在不同牧压下钙和磷含量变化不同,各处理间差异达到显著水平,放牧前后紫花针茅磷的含量显著高于对照,且以适度放牧为最高。与紫花针茅相比较而言,对照放牧前后高山嵩草钙含量与适度放牧差异不显著,而重度放牧与适度差异达到显著水平。放牧前高山嵩草各处理间磷含量差异显著,且以对照为最高;放牧后对照与适度放牧磷含量差异不显著,但重度放牧磷含量显著高于对照和适度放牧。同一处理水平下,放牧前后的研究结果表明,放牧能够减缓高寒草甸优势种高山嵩草与紫花针茅对钙和磷的吸收随着生育期的推进而下降的现象。不同植物种对相同牧压响应机制不同,高山嵩草耐牧性高于紫花针茅,放牧有利于牧草钙和磷的吸收,且重度放牧对钙和磷的吸收更有利。放牧家畜的采食、践踏等作用加速了牧草-土壤间钙和磷的循环,牧草补偿性生长对钙和磷吸收量增加,而这种趋势与放牧强度变化相一致。
4 结论
1)青藏高原高山嵩草草甸不同优势种对放牧强度响应不同,放牧能增加高山嵩草草甸优势种高山嵩草与紫花针茅粗蛋白的含量。
2)放牧能增加高山嵩草草甸优势种高山嵩草与紫花针茅粗脂肪的含量,可能是因为放牧家畜的采食促进牧草的补偿性生长,使幼嫩枝条比例上升进而使粗脂肪含量增加。但适度放牧的这种效应并不明显。
3)放牧有利于优势种高山嵩草与紫花针茅对钙和磷的吸收,且重度放牧对钙和磷的吸收更有利。
[1] 高永恒,陈槐,罗鹏,等.放牧强度对四川高寒草甸植物生物量及其分配的影响[J].生态与农村环境学报, 2008,24(3):26-32.
[2] 于格,鲁春霞,谢高地.青藏高原北缘地区高寒草甸土壤保持功能及其价值的实验研究[J].北京林业大学学报,2006,28(4):57-61.
[3] 许岳飞,益西措姆,付娟娟,等.青藏高原高山嵩草草甸植物多样性和土壤养分对放牧的响应机制[J].草地学报,2012,20(6):1026-1032.
[4] 郝力壮,刘书杰,吴克选,等.玛多县高山嵩草草地天然牧草营养评定与载畜量研究[J].中国草地学报,2011,33(1):84-88.
[5] 吴海艳,马玉寿,董全民,等.黄河源区藏嵩草沼泽化草甸地上生物量及营养季节动态研究[J].草业科学,2009,26(1):8-12.
[6] 赵彦光,洪琼花,谢萍,等.云贵高原石漠化地区人工草场营养价值评价研究[J].草业学报,2012,21(1):1-9.
[7] 侯扶江,杨中艺.放牧对草地的作用[J].生态学报,2006,26(1):244-263.
[8] 仁青吉,崔现亮,赵彬彬.放牧对高寒草甸植物群落结构及生产力的影响[J].草业学报,2008,17(6):134-140.
[9] 范国艳,张静妮,张永生,等.放牧对贝加尔针茅草原植被根系分布和土壤理化特征的影响[J].生态学杂志, 2010,29(9):1715-1721.
[10] Tilman D,Downing J A.Biodiversity and stability in grasslands[J].Nature,1994,367:363-365.
[11] Fahnestock J T,Knapp A K.Plant responses to selective grazing by bison:Interaction between light,herbivore and water stress[J].Vegetation,1994,115:123-131.
[12] 高英志,韩兴国,汪诗平.放牧对草原土壤的影响[J].生态学报,2004,24(4):790-797.
[13] Grime J P.Control of species diversity in herbaceous vegetation[J].Journal of Environmental Management,1973,1:151-167.
[14] 李春莉.放牧对短花针茅草原及糙羊茅草原植被和土壤影响的研究[D].呼和浩特:内蒙古农业大学,2008.
[15] 闫凯,张仁平,李德祥,等.新源县山地草原植被特征及植物营养对放牧强度的响应[J].草业科学,2011,23(8):1507-1511.
[16] 董全民,马玉寿,李青云,等.牦牛放牧率对小嵩草高寒草甸植物群落的影响[J].中国草地,2004,26(3):24-32.
[17] Garcia F,Carrere P,Soussana J F,etal.How do severity and frequency of grazing affect sward characteristics and the choices of sheep during the grazing season[J].Grass and Forage Science,2003,58:138-150.
[18] 江小雷,张卫国,杨振宇,等.不同干扰类型对高寒草甸群落结构和植物多样性的影响[J].西北植物学报,2003,23(9):1479-1485.
[19] 刘亚军.不同放牧强度对坝上草地植被和土壤理化性质的影响[D].保定:河北农业大学,2009.
[20] Risser P G,Patton W J.Ecosystem analysis of the tall grass prairie:Nitrogen cycle[J].Ecology,1982,63:1342-1351.
[21] Olofsson J,Stark S,Oksanen L.Reindeer influence on ecosystem processes in the tundra[J].Oikos,2004,105(2):386-396.
[22] Holland E A,Detling J K.Plant response to herbivore and below ground nitrogen cycling[J].Ecology,1990,71:1040-1049.
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
草业科学(2021年9期)2021-11-03 02:42:16
草业科学(2021年6期)2021-07-28 07:19:50
草地学报(2021年6期)2021-07-05 03:50:08
畜牧与饲料科学(2021年3期)2021-06-23 09:38:20
食品工业科技(2020年24期)2020-12-09 07:36:18
活力(2019年21期)2019-04-01 12:17:10
新课程·下旬(2017年10期)2018-01-02 08:05:46
