
芬太尼、咪达唑仑复合麻醉对大鼠认知功能及海马NR1、NR2B 表达的影响
2013-03-11 08:19:48赵艳梅闫玉仙常留栓
中风与神经疾病杂志 2013年1期
赵艳梅,闫玉仙,常留栓,陈 宁
多项研究发现,有些药物麻醉及术后常可发生认知功能障碍(postoperative cognitive dysfunction,POCD)[1],表现为认知能力异常、记忆受损、焦虑、人格改变、精神错乱等,严重者可出现痴呆,其确切机制迄今不明。研究认为,大脑神经元数量、与认知有关的神经递质谷氨酸、乙酰胆碱等及相应受体不足,引起神经元突触可塑性的改变可能是POCD 产生的原因之一[2]。海马富含谷氨酸能神经元,一直认为是与学习记忆有关的重要脑区。为了阐明海马神经元中谷氨酸受体与麻醉产生的认知障碍之间的关系,本研究测定了芬太尼、咪达唑仑复合麻醉对大鼠认知功能的影响,并检测了与学习记忆相关的谷氨酸受体——N-甲基-D-天冬氨酸(NMDA)受体亚单位NR1 和NR2B 在大鼠海马组织中的基因表达变化,探讨NMDA 受体与认知障碍形成的相关机制。
1 材料与方法
1.1 试剂与仪器
芬太尼(湖北宜昌人福药业有限责任公司,批号060208)、咪达唑仑(江苏恩华药业股份有限公司,批号20111219)、盐酸纳洛酮(美国Sigma 公司);氟马西尼(海南灵康制药有限公司,批号:091103);BP-2 大鼠无创血压计(成都泰盟科技有限公司)、Y 型电迷宫(张家港市生物医学仪器厂);DY-89 Ⅱ型电动玻璃匀浆器(宁波新芝生物科技股份有限公司);RNA 提取试剂盒(北京全式金生物技术有限公司);TaKaRa RNA PCR Kit AMV3.0(大连宝生物工程有限公司);100bp DNA MarkerⅠ、GoldView 核酸染料(北京博迈德科技发展有限公司);琼脂糖(Sigma),引物合成由北京奥科生物技术有限公司完成。
1.2 实验动物及分组
雄性Wistar 大鼠18 只,SPF 级,体重180~220g,由军事医学科学院实验动物中心提供,动物许可证号SCXK-(军)2007-004。雄性Wistar 大鼠随机分为3 组,分别为对照组、复合麻醉组、催醒组,每组6 只,复合麻醉组和催醒组均腹腔注射芬太尼0.5mg/kg、咪达唑仑50mg/kg 进行麻醉诱导,催醒组大鼠在麻醉起效30min 后腹腔注射纳络酮0.5mg/kg、氟马西尼0.8mg/kg 进行动物催醒。对照组腹腔注射等体积的生理盐水。
1.3 麻醉诱导与复苏
1.3.1 一般生理指标测定 复合麻醉组及催醒组动物在整个麻醉诱导与复苏过程中,每隔5~10min监测大鼠体温、呼吸频率、心率和血压。其中心率和血压的变化采用大鼠无创血压计观察,温度计记录肛温,通过观察大鼠胸廓运动来记录呼吸频率。
1.3.2 麻醉起效及复苏时间测定 麻醉诱导效应以翻正反射消失为标准。以大鼠前爪不能自行翻转为俯卧位,且维持时间≥10s 作为大鼠前爪翻正反射消失的标准。催醒组大鼠于诱导起效30min后注射拮抗剂纳络酮0.5mg/kg 和氟马西尼0.8mg/kg 催醒动物,记录动物复苏时间。复合麻醉组记录自然复苏时间。
1.4 复合麻醉对大鼠认知功能的影响(Y 迷宫实验)
动物麻醉复苏后继续饲养7d,1~7d 每日进行Y 迷宫实验,观察麻醉诱导及催醒对大鼠认知功能的影响。具体方法如下:在安静的暗室内用Y 迷宫测定。Y 迷宫箱底为铜栅间隔,每臂末端有信号灯,灯亮时箱底的铜栅无电流,提示为安全区。每次测试均在同一时间段进行,测试时调节刺激电压以保证大鼠在10s 内逃避跑动,大鼠受电击后直接跑到安全区为正确反应,否则为错误反应。每次大鼠跑至安全区后持续光亮15s,继而熄灯45s 后行下一次测试,灯光出现的方位按照随机次序变换,每日连续测量10 次,观察其正确反应次数,以正确反应率作为认知功能的评价指标。
1.5 麻醉诱导与药物催醒对NR1、NR2B 基因表达的影响(RT-PCR)
Y 迷宫实验结束后立即断头处死大鼠,于冰台上分离双侧海马,立即装入EP 管中置于液氮保存备用。根据NR1(GenBank NM_017010.1)和NR2B cDNA 序列(GenBank M91562.1),采用Primer Premier 5.0 软件自行设计引物,NR1:Sense:5’-TCGGTATCAGGAATGCG-3’;Anti-sense:5’-GGTGCTCGTGTCTTTGG-3 ’,328bp;NR2B:5 ’-GCTCAGCGACCTGTATGG-3’,Anti-sense:5’-TCGTCACTGCGTTCTTTGT-3’,381bp;β-actin:Sense:5’-CTGAGAGGGAAATCGTGCGT-3’,Anti-sense:5’-TGGAAGGTGGACAGTGAGGC-3’,448bp。PCR 反应条件见表1。

表1 PCR 反应条件
RNA 提取方法按照试剂盒说明书进行。扩增产物进行1.8%琼脂糖凝胶电泳,紫外透射仪下观察结果,图像采用Image J 分析软件进行处理。以目的基因条带的综合光密度与β-actin 基因条带综合光密度的比值作为目的基因的相对含量。
1.6 统计学处理
采用SPSS 10.0 软件进行统计分析,不同组别间的基因表达含量及Y 迷宫实验数据比较采用单因素方差分析。
2 结果
2.1 大鼠麻醉诱导与复苏时间及一般生理指标的变化
大鼠麻醉及复苏期间状态平稳,体温、呼吸频率、心率和血压均无明显变化。复合麻醉组麻醉起效时间(翻正反射消失时间)为3.12±0.25min,自然复苏时间为72.25±12.15min;催醒组大鼠麻醉起效时间为3.28±0.29min,给予拮抗剂后复苏时间为23.25±5.78min,较复合麻醉组复苏时间明显缩短(P<0.01)。
2.2 麻醉诱导对大鼠认知功能的影响
大鼠Y 迷宫认知功能的测定见表1。由表1 可以看出,复合麻醉组与正常对照组相比,正确反应率有所下降(P<0.01),拮抗剂催醒组与复合麻醉组相比,正确反应率明显升高(P<0.01)。提示芬太尼、咪达唑仑复合麻醉对大鼠短期内(1w)学习记忆能力有一定损害作用,麻醉后给予拮抗剂催醒对麻醉导致的学习记忆能力下降有明显的改善作用。
2.3 麻醉诱导与药物催醒对NR1、NR2B 基因表达的影响
麻醉诱导与拮抗剂催醒对NR1、NR2B 基因表达的影响结果见表2。由表2 可以看出,复合麻醉组与正常对照组相比,大鼠海马组织内NR2B mRNA 表达量显著降低(P<0.01),催醒组NR2B mRNA 表达量与复合麻醉组相比显著升高(P<0.01),与正常对照组相比无明显差异。NR1 mRNA 表达量在各组间均无明显变化。说明芬太尼、咪达唑仑复合麻醉可降低海马部位的NR2B 的mRNA 表达,使用拮抗剂催醒可显著增加海马部位的NR2B 的mRNA 表达。
表1 各组大鼠Y 迷宫实验结果(±s)
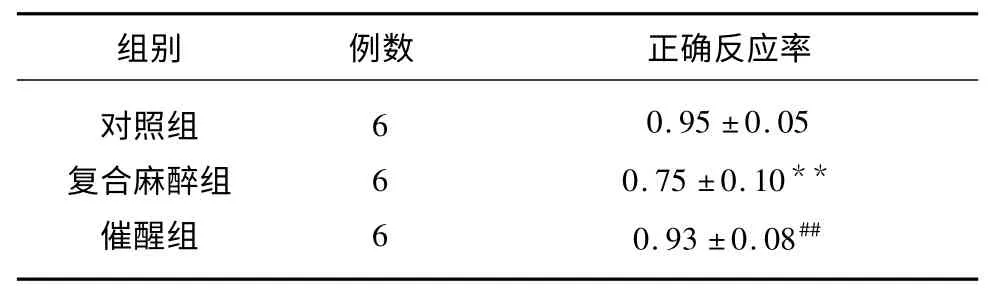
表1 各组大鼠Y 迷宫实验结果(±s)
与对照组比较**P<0.01;与复合麻醉组比较##P<0.01
表2 麻醉诱导与拮抗剂催醒对大鼠海马NR1、NR2B mRNA 表达的影响(±s)
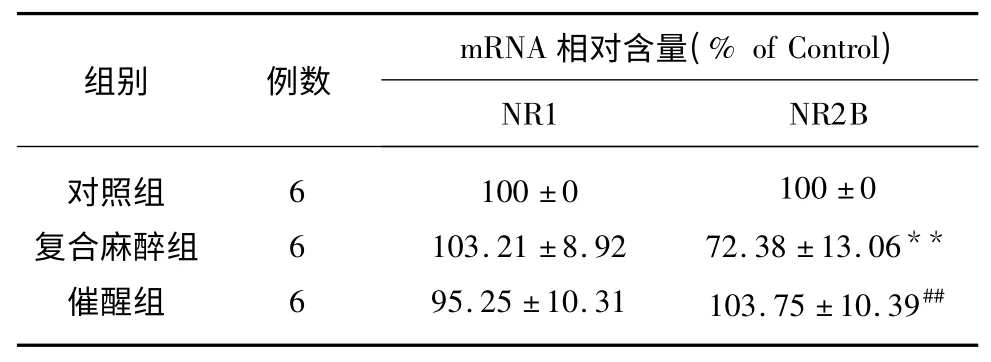
表2 麻醉诱导与拮抗剂催醒对大鼠海马NR1、NR2B mRNA 表达的影响(±s)
与对照组比较**P<0.01;与复合麻醉组比较##P<0.01
3 讨论
越来越多的证据表明,全身麻醉后大脑发生了基因表达改变[3]、β 淀粉样沉积[4]、神经元凋亡[5,6]等广泛而持久的变化,单纯的短时间麻醉即可造成实验动物长时间的认知功能障碍[7~9],临床上常常表现为术后认知功能障碍(POCD)。Moller 等[10]在一项临床调查中发现,10%~14%的老年患者全身麻醉或手术后发生了长期的认知功能障碍。而中年和青年患者术后1 至2w 的认知功能障碍也有很高的发生率[11]。咪达唑仑是一种常见静脉麻醉药物,是当前临床应用的唯一的水溶性苯二氮艹卓类药物,临床上常与芬太尼类化合物配伍使用。研究发现,镇静剂量的咪达唑仑可以抑制大鼠的记忆获得和巩固并维持较长一段时间[12],大鼠腹腔注射咪达唑仑可出现东莨菪碱所致的认知功能障碍加重的现象[13],因此认为咪达唑仑可能是导致POCD 的一个重要因素。芬太尼是一种强效麻醉性镇痛药,属于阿片受体激动剂,与咪达唑仑在镇痛、催眠作用上具有一定的协同作用,可减少单一药物的用药剂量,减轻不良反应[14]。本研究采用Y 迷宫实验观察了芬太尼和咪达唑仑麻醉后对大鼠认知功能的影响,结果发现,芬太尼、咪达唑仑复合麻醉后7d 内大鼠逃避伤害刺激的正确反应率有所下降(P<0.01);拮抗剂催醒组与复合麻醉组相比,正确反应率明显升高(P<0.01)。提示芬太尼、咪达唑仑复合麻醉短期内对大鼠学习记忆能力即认知功能有一定损害作用,麻醉后快速给予纳洛酮(阿片受体拮抗剂)和氟马西尼(苯二氮艹卓类受体拮抗剂)催醒对麻醉导致的学习记忆能力下降有明显的改善作用。
谷氨酸是哺乳动物脑内最重要的一种兴奋性氨基酸,其参与长时程增强(LTP)的产生和大脑的兴奋性传递,对大脑的学习记忆能力起重要作用。而谷氨酸主要是通过其离子型受体N-甲基-D-天冬氨酸受体(NMDAR)介导一系列高级神经活动,它被认为是突触可塑性和皮质及海马神经元LTP 的主要调控者,是构成中枢神经系统学习记忆功能的重要基础。NR1 是NMDA 受体复合物的功能性亚单位,是NMDA 受体通道的必要组成部分,它的含量代表了NMDA 受体的总量,NR2B 是主要的调节亚单位,可以反映NR 受体的功能变化。有研究证明:选择性敲除小鼠海马CA1区锥体细胞的NR1 亚单位后,其NMDAR 诱导的LTP 被破坏,且小鼠表现为空间记忆障碍[15]。慢性复合应激可通过使NR2B表达增多增强大鼠的学习记忆[16]。铅中毒可导致海马神经元NR2B 表达下降并引起不同程度记忆受损[17]。敲除NR2B 基因后小鼠记忆能力下降,而移植转染NR2B 基因可使大鼠记忆明显增高[18]。以上说明,NR1 和NR2B 受体的功能与学习记忆密切相关。因此,本研究选取了NR1 和NR2B 两种亚基进行研究,通过观察2 种亚基的mRNA 表达变化,间接反映NR 受体的功能,探讨麻醉后认知障碍与NMDA 受体的关系。
RT-PCR 实验结果显示,复合麻醉组与正常对照组相比,大鼠海马组织内NR2B mRNA 表达量显著降低(P<0.01),催醒组NR2B mRNA 表达量与复合麻醉组相比显著升高(P<0.01),与正常对照组相比无明显差异。NR1 mRNA 表达量在各组间均无明显变化。说明芬太尼、咪达唑仑复合麻醉可降低海马部位的NR2B 的mRNA 表达,使用拮抗剂催醒可显著增加海马部位的NR2B 的mRNA 表达。该结果与大鼠Y 迷宫实验结果相一致,提示海马组织内NR2B 表达下降可能与认知障碍的产生关系密切,该结果与以往研究相一致[19,20]。NR1 基因表达在认知障碍大鼠并未出现表达改变,研究推测NR1基因在认知过程的形成中起着必不可少的作用,但是认知障碍的形成可能主要由调节亚单位NR2B 的变化引起,NR2B 基因可能起着调节学习记忆功能的重要作用。具体NMDA 受体在认知障碍的形成过程中行使何种功能,各亚型之间存在何种相互作用,NMDA 受体与其他受体蛋白是否存在交互作用,具体机制如何将是我们下一步研究的方向。
[1]李 琪,江 伟.术后认知功能障碍研究进展[J].医学综述,2010,16(2):231-234.
[2]Akashi K,Kakizaki T,Kamiya H,et al.NMDA receptor GluN2B(GluR epsilon 2/NR2B)subunit is crucial for channel function,postsynaptic macromolecular organization,and actin cytoskeleton at hippocampal CA3synapses[J].J Neurosci,2009,29(35):10869-10882.
[3]Mawhinney LJ,de Rivero Vaccari JP,Alonso OF,et al.Isoflurane/nitrous oxide anesthesia induces increases in NMDA receptor subunit NR2B protein expression in the aged rat brain[J].Brain Res,2012,1431(11):23-34.
[4]Xie Z,Dong Y,Maeda U,et al.The common inhalation anesthetic isoflurane induces apoptosis and increases amyloid beta protein levels[J].Anesthesiology,2006,104:988-994.
[5]Jevtovic-Todorovic V,Hartman RE,Izumi Y,et al.Early exposure tocommon anesthetic agents causes widespread neurodegeneration in the developing rat brain and persistent learning deficits[J].J Neurosci,2003,23:876-882.
[6]Wise-Faberowski L,Zhang H,Ing R,et al.Isoflurane-induced neuronal degeneration:an evaluation in organotypic hippocampal slice cultures[J].Anesth Analg,2005,101:651-657.
[7]Culley DJ,Baxter MG,Yukhananov R,et al.Long-term impairment of acquisition of a spatial memory task following isoflurane-nitrous oxide anesthesia in rats[J].Anesthesiology,2004,100:309-314.
[8]Culley DJ,Baxter M,Yukhananov R,et al.The memory effects of general anesthesia persist for weeks in young and aged rats[J].Anesth Analg,2003,96:1004-1009.
[9]Culley DJ,Baxter MG,Crosby CA,et al.Impaired acquisition of spatial memory 2 weeks after isoflurane and isoflurane-nitrous oxide anesthesia in aged rats[J].Anesth Analg,2004,99:1393-1397.
[10]Moller JT,Cluitmans P,Rasmussen LS,et al.Long-term postoperative cognitive dysfunction in the elderly ISPOCD1 study.ISPOCD investigators.International Study of Post-Operative Cognitive Dysfunction[J].Lancet,1998,351:857-861.
[11]Johnson T,Monk T,Rasmussen LS,et al.Postoperative cognitive dysfunction in middle-aged patients[J].Anesthesiology,2002,96:1351-1357.
[12]Savic MM,Obradovic DI,Ugresic ND,et al.Bidirectional effects of benzodiazepine binding site ligands on passive avoidance acquisition and retention:differential antagonism by flumazenil and beta-CCt[J].Psychopharmacology(Berl),2005,180(3):455-465.
[13]张 捷,吴新民.咪达唑仑对东蓑若碱致大鼠认知功能障碍的影响[J].中华麻醉学杂志,2007,27(2):160-163.
[14]金 茹,刘传飞,赵 娟,等.咪达唑仑和芬太尼的相互作用[J].药学与临床研究,2010,18(5):432-434.
[15]Shimizu E,Tang YP,Rampon C,et al.NMDA receptor-dependent synaptic reinforcement as a crucial process for memory consolidation[J].Science,2000,290(5494):1170-1174.
[16]程少容,刘能保,张敏海,等.慢性复合应激对大鼠学习与记忆及海马NMDA 受体亚基NR2A 和NR2B 表达的影响[J].神经解剖学杂志,2004,20:275-280.
[17]LauWK,Yeung CW,Lui PW,et al.Different trends in modulation of NMDAR1 and NMDAR2B gene exp ression in cultured cortical and hippocampal neurons after lead exposure[J].Brain Research,2002,932:10-24.
[18]Tang YP,Shimizu E,Dube GR,et al.Genetic enhancement of learning and memory in mice[J].Nature,1999,401:63-69.
[19]孙钦建,刘学武,王爱华,等.颞叶癫痫大鼠认知功能与海马区NR2B 表达的关系[J].山东大学学报(医学版),2008,46(7):676-680.
[20]陈 谊,蔡文玮,盛 净.糖尿病模型大鼠认知功能障碍及海马N-甲基-D-天冬氨酸受体表达变化[J].实用医学杂志,2010,26(17):3098-3101.
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05 01:33:06
中华养生保健(2020年3期)2020-11-16 00:53:20
广东医科大学学报(2020年4期)2020-08-24 07:11:20
创新作文(小学版)(2019年10期)2019-09-25 08:12:28
小学生学习指导(低年级)(2017年5期)2017-05-04 04:14:38
海南医学(2016年8期)2016-06-08 05:43:00
中国卫生标准管理(2015年5期)2016-01-14 05:17:05
中国社区医师(2015年14期)2015-12-24 00:37:31
作文与考试·小学高年级版(2015年17期)2015-05-30 10:48:04
生殖医学杂志(2015年11期)2015-02-28 16:32:16
