
桑沟湾楮岛近岸海域大叶藻生态学特征的基础研究
2013-03-07 07:12刘炳舰刘旭佳张晓梅
海洋科学 2013年1期
刘炳舰, 周 毅, 刘旭佳, 刘 鹏, 张晓梅
(1. 中国科学院 海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071; 2. 中国科学院 研究生院,北京 100049; 3. 广西海洋研究所 海洋生物技术重点实验室, 广西 北海 536000)
桑沟湾楮岛近岸海域大叶藻生态学特征的基础研究
刘炳舰1,2, 周 毅1, 刘旭佳3, 刘 鹏1,2, 张晓梅1,2
(1. 中国科学院 海洋研究所 海洋生态与环境科学重点实验室, 山东 青岛 266071; 2. 中国科学院 研究生院,北京 100049; 3. 广西海洋研究所 海洋生物技术重点实验室, 广西 北海 536000)
2010年8月至2011年9月期间, 对桑沟湾楮岛沿海的大叶藻(Zostera marinaL.)进行了一周年调查。结果显示, 大叶藻周年平均株高变化范围为(16.97±5.99)~(87.60±20.68)cm, 大叶藻平均密度为(613±201)株/m2, 单株生物量为0.97~5.31g/株, 单位面积平均生物量为594.61~3255.03 g/m2, 大叶藻高度和湿重的周年生长变化与水温的变化趋于一致, 呈正相关性; 大叶藻生长环境水温为 5.3~25.6℃,平均温度16.3℃; 盐度为28.9~31.3, 平均盐度30.6; 底质类型为砾砂。大叶藻根茎、叶鞘和叶中碳含量的平均值分别为32.68%±1.27%、33.78%±3.06%、37.01%±1.86%, 氮含量平均值分别为1.62%±0.63%、2.79%±0.81%、3.10%±0.81%, 磷含量的平均值分别为0.28%±0.04%、0.51%±0.10%、0.48%±0.07%。
大叶藻(Zostera marinaL.); 海草; 生物量; 栖息环境; 生态学特征
大叶藻(Zostera marinaL.)作为海草的典型代表,通常在沿海潮间带和潮下带较浅的水域形成广大的群落, 形成海草床[1], 在滨海生态系统中发挥着重要的生态功能, 例如: 提高滨海生态系统的生产力; 为一些重要的经济鱼类和贝类提供复合的食物网、栖息场所和育幼场所[2]; 缓冲减弱水流并可以固定底质[3]; 加速悬浮颗粒物的沉降, 增加水体透明度, 净化水质[4-5], 加速滨海生态系统中的营养循环等等。但长期以来, 受自然因素以及人类活动的影响, 造成了全世界海草资源退化严重。
国外学者对海草生态学方面都做了很多工作, 包括海草时空分布、能量流动、物质循环等方面[6-9], 为海草生态系统研究奠定了基础。中国对海草研究起步较晚, 最早有杨宗岱和吴宝玲等[10-12]论述了中国海草的地理分布、生产力以及海草床的结构等, 我国南方对海草的研究较多, 例如范航清, 韩秋影, 黄小平, 李颖虹等[13-17]对我国华南沿海的海草床做了较为系统的研究, 另外任国忠, 刘志鸿, 叶春江[18-22]等也做过一些基础研究。本研究以桑沟湾楮岛沿岸的大叶藻为研究对象, 调查了桑沟湾大叶藻生长高度、生物量、碳、氮、磷含量的季节变化等生态特征。为我国海草的进一步深入研究提供科学的基础数据。
1 材料与方法
1.1 调查时间及地点
调查时间为2010年8月至2011年9月, 调查地点位于山东荣成楮岛海区, 该海区位于桑沟湾南岸的最东端, 潮汐为不规则混合半日潮, 沿岸为侵蚀崖岸。大叶藻平行于海岸分布, 由低潮带延伸至水深3m 处, 这里集中分布着多种海草, 其中大叶藻为主要优势种, 形成了桑沟湾内面积最大的大叶藻床。调查位置示意图如图1所示。
1.2 调查方法
根据大叶藻的分布区域、离岸远近以及水深等条件, 在桑沟湾楮岛海区的调查地点设置两个站位: SGW-I和 SGW-II, 它们的坐标分别为(37° 02′483″N, 122°34′129″E)和(37°02′529″N, 122°34′125″E)。2010年8月~2011年 9月逐月调查(因实验条件限制, 缺少2011年1月和2月数据), 在每月退大潮时的低潮时段到大叶藻海草场调查采样, 首先在现场测量大叶藻密度、高度, 其生长环境的水温、盐度等数据, 然后在每个站点采集大叶藻样品及其生长环境的水样、底泥样品。大叶藻样品是在设置的站点随机的采集约 50株, 水样是用水样瓶采集表层海水约 200 mL, 底泥样品是用采泥器采集底泥约200 g。

图1 桑沟湾海草调查地理位置Fig. 1 Geographical location of eelgrass survey in Sanggou Bay
1.3 数据的分析与处理
现场采集的样品带回实验室后, 其中大叶藻样品洗净后挑15株称其湿质量, 然后取新鲜的大叶藻,将叶、叶鞘和根茎分开处理, 60℃烘干后用研钵磨碎,过80目样品筛, 测定大叶藻植株各部位C、N、P元素含量; 水样用营养盐自动分析仪测定其营养盐含量; 底泥样品除用激光粒度分析仪测定泥沙的粒径外, 也测定其中C、N、P元素的含量。大叶藻及底泥中的C、N元素利用Perkin-Elmer 240C元素分析仪(Perkin-Elmer Company, USA)测定; P采用磷钼蓝分光光度法进行测定。样品的现场采集、保存、处理、测定和分析等过程参照《海洋调查规范》(GB/T 12763-2007)、《海洋监测规范》(GB17378-2007)和《海水水质标准》(GB3097-1997)等进行。
2 结果与分析
2.1 大叶藻生长环境的水质和底质特征
水质特征: 调查海域水温周年变化范围在5.3~25.6℃, 平均水温16.3℃。盐度范围为28.9~31.3,平均盐度 30.6(图 2)。底质特征: 分析结果显示, 该海域大叶藻海草床底质平均粒径1.75mm, 砾石、砂、粉砂、黏土、贝壳所占比例分别为20.32%、59.04%、16.91%、3.74%、8.42%(图3), 因此大叶藻生长环境底质定义为砾砂。底泥中的氮含量为0.014%~0.099%,平均氮含量为 0.034%±0.029%; 碳含量为0.246%~0.977%, 平均碳含量为 0.599%±0.264%;磷含量为 0.031%~0.126%, 平均磷含量为 0.084%± 0.027%。
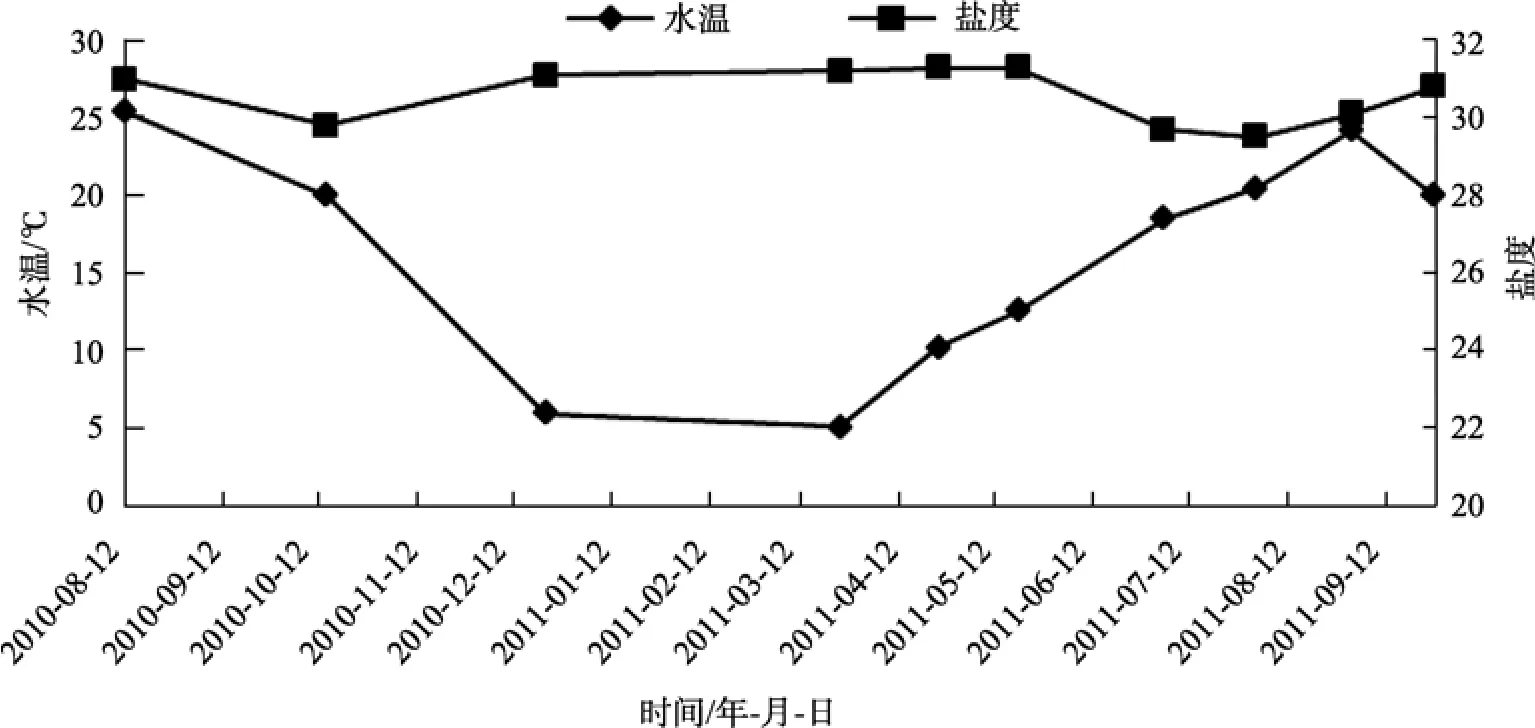
图2 桑沟湾水温和盐度的周年变化Fig. 2 Annual changes of water temperature and salinity in Sanggou Bay

图3 桑沟湾大叶藻底质泥沙粒径分析Fig. 3 Particle-size spectra of sediment for Z.marina in Sanggou Bay
2.2 桑沟湾大叶藻株高和生物量的周年变化
大叶藻的平均株高随水温的周年变化见图4。SGW-I站点大叶藻平均株高最大值为(87.60±20.68)cm, 出现在 2011年8月; 最小值为(18.80±4.13)cm, 出现在2011年3月。SGW-II站点大叶藻平均高度最大值为(68.30±21.88)cm, 出现在2011年7月; 最小值为(16.97±5.99)cm, 出现在2011年3月。
大叶藻的地上部分平均湿重随水温的周年变化见图5。SGW-I站点大叶藻地上部分平均湿重最大值为3.84 g/株, 出现在2011年8月; 最小值为1.16 g/株, 出现在2011年4月。SGW-II站点大叶藻地上部分平均湿重最大值为 5.31g/株, 也是出现在 2011年 8月; 最小值为 0.97 g/株, 出现在2011年3月。现场测的调查站点大叶藻的平均密度约为(613±201) 株/m2, 因此可以计算出 SGW-I站位大叶藻地上部分的生物量为 711.08~2353.92 g/m2; SGW-II站位大叶藻地上部分的生物量为594.61 ~3255.03 g/m2。

图4 桑沟湾各站点大叶藻(营养枝)平均株高变化Fig. 4 Average hight changes of Z. marina with water temperature in sites SGW-I and SGW-II
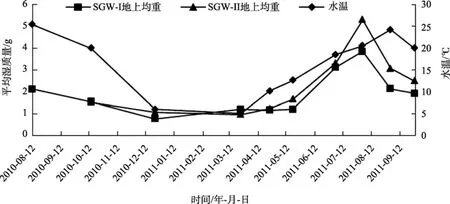
图5 桑沟湾各站点大叶藻(营养枝)地上部分平均湿质量变化Fig. 5 Average wet weight changes of the overground part of Z. marina with water temperature in sites SGW-I and SGW-II
从图4和图5中可以看出, 大叶藻平均高度和地上部分平均湿质量与水温变化趋于一致, 呈正相关性。从2010年8月到2011年9月水温呈先降低后升高的变化趋势, 大叶藻高度和湿质量的变化也是一样, 从夏季到冬季随水温的降低而迅速下降, 到冬季达到最小值, 春季到夏季又随水温的上升而迅速增加, 到夏季达到最大值。大叶藻与其他温带陆地高等植物一样, 其生长随着季节的变化而变化, 有着周年中萌发、生长、凋落的变化节律, 而在其生长过程中水温是影响其形态、生长和繁殖的一个很重要因素。与很多陆地草本植物不同的是, 大叶藻繁殖上存在着有性繁殖和无性繁殖相互交替的现象, 在相邻生境中采取一年生和多年生地策略, 因此在凋落时期的秋冬季仍然保存有一定的地上部分生物量。
2.3 大叶藻碳、氮、磷元素含量及比值变化
大叶藻各部位中碳、氮、磷元素含量的周年变化见图6(其中 7月份碳氮测量数据由于偏差过大舍弃)。根茎、叶鞘和叶中碳含量的平均值分别为32.68%±1.27%、33.78%±3.06%、37.01%±1.86%, 氮含量平均值分别为 1.62%±0.63%、2.79%±0.81%、3.10%±0.81%, 磷 含 量 的 平 均 值 分 别 为0.28%±0.04%、0.51%±0.10%、0.48%±0.07%。从图6中可以总结出, 叶, 叶鞘和根茎有机碳、氮和磷含量变化趋势类似, 它们含量多少的关系为, 根茎<叶鞘<叶。地上部分碳含量略高于地下部分; 叶鞘和叶中的氮磷含量相差不大, 约为根茎氮磷含量的两倍。由此分析, 叶是大叶藻进行光合作用产生有机物的主要部位, 叶鞘跟叶的分化并不明显, 主要起支撑和运输有机物的作用, 而根茎主要起固着和储存作用,所以地上部分有机质含量比地下部分高, 而叶鞘和叶中的有机质含量相差不大。
从大叶藻各部位碳、氮、磷含量的周年变化曲线中可以得出, 自2010年8月到2011年8月, 大叶藻各部位碳氮磷含量的变化均呈先升高后降低的趋势, 即从夏季到秋季升高, 从春季到夏季再降低, 结合大叶藻高度和生物量的变化情况, 我们分析, 夏季过后大叶藻开始凋落和腐烂, 因此其有机质含量也开始降低, 大叶藻是营一年生和多年生的植物,到冬季是根茎萌发出侧生苗的时间, 而春季是大叶藻营养生长旺盛的季节, 光合作用强, 所以冬季和春季其有机质含量较高, 然后从春末大叶藻开始生殖生长, 有机质多转移到花和种子中去, 大叶藻营养枝有机质含量又开始降低。
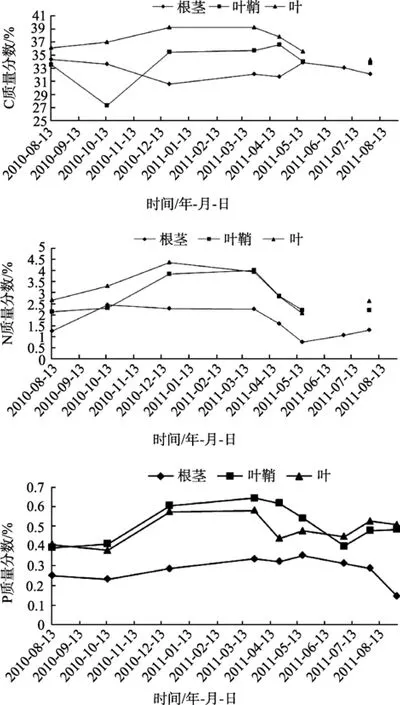
图6 SGW-I站点大叶藻不同部位碳、氮、磷元素含量周年变化Fig.6 Seasonal changes of carbon, nitrogen and phosphorus contents in different parts of Z. marina in site SGW-I
大叶藻各部位C:N、C:P、N:P比见图7。根茎、叶鞘和叶碳氮比的平均值分别为 20.81%±6.64%、12.77%±2.92%、12.54%±2.78%, 碳磷比的平均值分别为 112.06%±18.78%、65.51%±10.16%、78.19%± 12.54%, 氮磷比的平均值分别为 5.93%±2.33%、5.27%±0.88%、6.45%±1.49%。碳氮比和碳磷比反映植物生长速度以及植物对氮和磷的利用效率, 氮磷比则可以反映植物生长受 N或 P的限制情况。从C:N、C:P的周年变化中可以总结出, 大叶藻各部位的C:N、C:P比自2010年8月到2011年8月先降低后升高, 说明大叶藻从夏季到秋季生长速度减慢,冬季生长速度最慢, 春季生长速度又加快。N:P比值<20说明氮元素是大叶藻生长的限制性因子。

图7 SGW-I大叶藻不同部位碳氮比、氮磷比和碳磷比周年变化Fig. 7 Seasonal changes of C:N ratio, N:P ratio and C:P ratio in different parts of Z. marina in site SGW-I
3 讨论
大叶藻形成的海草床是近岸水域重要的湿地生态系统, 具有重要的生态价值和经济价值[23], 尽管其面积只占海洋总面积的约 0.15%, 但却是海洋中重要的初级生产者。据报道, 海草场内有近20种生物种群生存栖息, 有丰富的饵料生物, 如浮游的桡足类、端足类, 底栖的沙蚕、海蚯蚓等。海草场本身也具有较高的固碳能力, 杨宗岱[12]研究指出, 80年代山东沿海大叶藻碳的固定量为564 g/m2。另外, 海草场的存在, 增加了海草场内流体的阻力, 从而改变了海草场内的流体动力过程, 加速了悬浮颗粒的沉降, 对稳定海底底质和净化水质有着积极的作用,因而被称为生态系统工程师[25]。
但长期以来, 由于受到自然因素和人为因素的影响, 使世界上大部分分布区域大叶藻的资源都受到了很大程度的破坏。飓风、台风等剧烈天气情况引起的风暴潮、台风浪冲刷海草,可能会将海草连根拔起, 或是将滩涂中的泥沙冲刷起来埋没海草从而影响海草的生长, 对海草场造成破坏, 导致海草资源的损失。另外病害也可能是大叶藻衰退的原因之一, 例如, 1930年到1933年间北大西洋沿岸由于枯萎病(Wasting disease)爆发, 导致该地区大部分的大叶藻死亡[26]。相比于自然因素, 人为因素对海草床的破坏更大, 人类活动如海水养殖、围网捕鱼、填海、溢油、船舶活动、人为污染与开挖航道等提高了水体的浑浊度, 导致藻类的过量繁殖, 最终造成了海草的消亡, 直接和间接的导致了海草床的衰退。同世界上其他国家和地区一样, 我国的大叶藻资源也处于不断衰退之中。20世纪80年代以前, 山东省沿岸海域潮间带2~ 5 m水深范围内生长有丰富的大叶藻,如今已大面积退化、衰竭, 有些甚至不复存在[27]。
大叶藻的大范围消失引起了全世界范围内对大叶藻生理、生态及恢复等众多方面的关注, 并且展开了关于大叶藻各方面的很多研究。研究较多的主要集中在欧美等国家, 而且他们的研究也很全面和成熟; 国内对大叶藻的研究起步较晚, 研究也相对较少, 但近些年来国内关于大叶藻研究方面的文章也迅速增加, 本文通过对桑沟湾沿岸大叶藻生态学性状的研究, 为之后的研究工作提供一些基础参考。
[1] Den Hartog C.The sea-grasses of the world[M].North Holland Pub1 Co., Amsterdam,1970.
[2] Newell S Y. Multiyear patterns of fungal biomass dynamics and productivity within naturally decaying smooth cord grass shoots[J]. Limnology and Oceanography, 2001, 46(3): 573-583.
[3] Fonseca M S. Sediment stabilization byHalophiladecipiensin comparison to other seagrasses [J]. Estuary Coastal Shelf Sciences, 1989, 29: 501-507.
[4] Fonseca M S, Fisher J S. A comparison of canopy friction and sediment movement between four species of seagrass with reference to their ecology and restoration [J]. Marine Ecology Progress Series, 1986, 29: 15-22.
[5] Hemminga M A, Harrison P G, Lent F van. The balance of nutrient losses and gains in seagrass meadows [J], Marine Ecology Progress Series, 1991, 7l: 85-96.
[6] Hemminga M A, Duarte C M. Seagrass Ecology[M]. Cambridge: Cambrige UniversityPress, 2000: 20-22.
[7] Jackson E L, Rowden A A, Attrill M J, et al. The importance of seagrass beds as a habitat for fishery species[J].Oceanology and Marine Biology-An Annum Review, 2001, 39: 269-303.
[8] Gambi M C, Nowell A R M, Jumars P A.Flume observations on flow dynamics in Zosteramarna (eelgrass) beds[J]. Marine Ecology Progress Series, 1990, 61: 159-169.
[9] Bouma T J, deVries M B, LowE, et al. Trade-offs related to ecosystem engineering: a case study on stiffness of emerging macrophytes[J]. Ecology, 2005, 86: 2187-2199.
[10] 杨宗岱. 中国的海草生态学研究[J].海洋科学, 1982, 2: 34-37.
[11] 杨宗岱.中国海草植物地理学的研究[J].海洋湖沼通报, 1979, 2:41-45.
[12] 杨宗岱, 吴宝铃. 中国海草场的分布、生产力及其结构与功能的初步探讨[J]. 生态学报, 1981, 1(1): 84-88.
[13] 范航清, 彭胜, 石雅君, 等. 广西北部湾沿海海草资源与研究现状[J].广西科学, 2007, 14(3): 289-295.
[14] 范航清, 郑杏雯. 海草光合作用研究进展[J].广西科学, 2007, 14(2): 180-185, 192.
[15] 韩秋影, 黄小平, 施平, 等. 2007. 广西合浦海草床生态系统服务功能价值评估[J]. 海洋通报, 26(3): 33-38.
[16] 韩秋影, 黄小平, 施平, 等. 2007. 人类活动对广西合浦海草床服务功能价值的影响[J]. 生态学杂志, 26(4): 544-548.
[17] 李颖虹, 黄小平, 许战洲, 等. 2007广西合浦海草床面临的威胁与保护对策[J]. 海洋环境科学, 26(6): 587-590.
[18] 叶春江, 赵可夫. 高等植物大叶藻研究进展及其对海洋沉水生活的适应[J]. 植物学通报, 2002, 19(2): 184-193.
[19] 叶春江, 赵可夫. 盐分胁迫对大叶藻某些胞内酶耐盐性及其生理功能的影响[J]. 植物学报, 2002, 44(7): 788-794
[20] 任国忠, 张起信. 王继成, 等. 移植大叶藻提高池养对虾产量的研究[J]. 海洋科学, 1991, 1: 52-57.
[21] 刘志鸿, 董树刚, 牟海津, 等. 青岛汇泉湾大叶藻种群遗传多样性的研究[J]. 海洋水产研究, 1998, 19(2): 27-32.
[22] 汪文俊, 王广策, 黄勃, 等. 大叶藻(Zostera marinaL.)PSI和 PSII复合物的分离鉴定[J]. 海洋与湖沼, 2004, 35(5): 446-452.
[23] 于函, 马有会, 张岩, 原永党. 大叶藻的生态学特征及其与环境的关系[J].海洋湖沼通报, 2007增刊: 112-120.
[24] Edmund P G, Short F T. World Atlas of Seagrasses[M]. California : University of California press, 2003.
[25] 李文涛,张秀梅. 海草场的生态功能[J]. 中国海洋大学学报, 2009, 39(5): 933-939.
[26] Green E P, Short F T. World atlas of seagrasses [M]. California: Universityof California press, 2003.
[27] 李恒, 李美真. 藻场的生态作用及人工藻场建设的现状[J]. 中国水产, 2006(11): 77-79.
(本文编辑: 梁德海)
Ecological characteristics research ofZostera marinaL. in Sanggou Bay
LIU Bing-jian1,2, ZHOU Yi1, LIU Xu-jia3, LIU Peng1,2, ZHANG Xiao-mei1,2
(1. Key Laboratory of Marine Ecology & Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China; 2. Graduate University of Chinese Academy of Science,Beijing 100049, China; 3. Key Laboratory of Marine Biological technology, Guangxi Institute of Oceanology, Beihai 536000, China)
May,2,2012
Zostera marinaL.; seagrass; biomass; habitat; ecological characteristics
The ecological characteristics ofZostera marinaL. in Sanggou Bay was studied from August 2010 to September 2011. The results showed that the annual mean shoot height of eelgrass ranged from (16.97±5.99) cm to (87.60±20.68) cm, the mean density was (613±201) ind/m2, and the individual biomass was 0.97~5.31 g/ind. The average biomass of eelgrass meadow changed from 594.61 to 3255.03 g/m2. The average height and wet weight of the eelgrass changed with water temperature. The water temperature range of the eelgrass habitat was 5.3~25.6℃with an average temperature of 16.3 ℃. The salinity ranged from 28.9 to 31.3 ppt with an average salinity of 30.6 ppt. The main sediment was gravel sand. The annual average contents of carbon in rhizome, leaf sheath and leaf of eelgrass were 32.68%±1.27%, 33.78%±3.06% and 37.01%±1.86%, respectively. Similarly, the annual average contents of nitrogen were 1.62%±0.63%, 2.79%±0.81%, and 3.10%±0.81 %, respectively, and the annual average contents of phosphorus were 0.28%±0.04%, 0.51%±0.10%, 0.48%±0.07% respectively in rhizome, leaf sheath and leaf of eelgrass.
S917.3
A
1000-3096(2013)01-0042-07
2012-05-02;
2012-08-13
海洋公益性行业科研专项重点项目(200805069/201005009);国家科技支撑计划项目(2011BAD13B06/2010BAC68B01); 国家自然科学基金创新研究群体科学基金项目(41121064; 30972268); 青岛市市南区科技发展计划项目(2009517HY)
刘炳舰(1984-), 男, 硕士, 主要从事海洋生态学研究, 电话: 15866805287, E-mail: lbj0812@gmail.com; 周毅, 通信作者, 研究员,电话: 0532-82898646, E-mail: yizhou@qdio.ac.cn
猜你喜欢
汽车实用技术(2022年14期)2022-07-30
孩子(2020年5期)2020-06-08
电子制作(2019年7期)2019-04-25
智慧少年·故事叮当(2019年12期)2019-02-18
电子制作(2018年17期)2018-09-28
动漫界·幼教365(小班)(2018年5期)2018-05-14
琴童(2017年7期)2017-07-31
小学科学(2017年5期)2017-05-26
花卉(2016年21期)2016-07-31
创业家(2015年3期)2015-02-27
