
不同投喂方式对革胡子鲶消化酶的影响
2013-02-20 06:59朱国霞孙学亮陈成勋郭永军程民杰黄亚东
饲料工业 2013年14期
■ 朱国霞 孙学亮 陈成勋 郭永军 程民杰 黄亚东
(天津农学院水产科学系天津市水产生态及养殖重点实验室,天津 300384)
由于自然等原因造成生长停滞或负增长时,当胁迫条件改善或消失,动物表现出一个快速的迸发式的生长,被称为补偿生长。充分考虑鱼类的补偿生长,借助一定的数学模型,在渔业资源管理上能够更加准确诊断和预测鱼类种群资源增长对食物的需求量。通过生物补偿生长机制的研究,揭示生物适应饥饿胁迫的生理生态对策,并以此采取科学的饲养措施,已在提高饲料转化效率,缓解因养殖而产生的诸如环境污染等一系列问题和促进养殖业的健康可持续发展等方面显示出巨大的经济、社会和生态潜力,已引起养殖专家的广泛关注和重视。
革胡子鲶(clarias leather)又称埃及塘虱,原产于非洲的尼罗河流域。革胡子鲶具有生长速度快、产量高、食性杂、个体大、耐低氧、抗病力强、水质条件要求低、生产周期短等优点,是值得大力推广的养殖品种。许多学者已经完成了革胡子鲶的生物学特性、养殖理论、人工繁殖、营养需求、生长性能、血液常数值、生长激素cDNA克隆与蛋白质结构分析等研究。但在目前通过改变投喂方式来促使革胡子鲶补偿生长的研究较少,本研究考察了革胡子鲶在不同间歇停喂后,其各消化器官蛋白酶、脂肪酶以及淀粉酶的变化情况。为鱼类补偿生长的研究提供参考数据。
1 材料与方法
1.1 试验材料
试验鱼为革胡子鲶鱼种,均为1年龄。试验鱼外观正常,体质健壮,无病害,体重(128.36±1.42)g,共180尾,分别放在蓝色长方形塑料箱(70 cm×50 cm×50 cm)中,每箱20尾。暂养15 d,使其适应试验环境,减少应激反应。暂养期间水温19~23℃。为了减少试验期内鱼类排泄物对试验的影响,试验前停食1 d。在相同条件下养殖,待生理指标稳定后进行试验。
1.2 试验设计
试验设3种投喂组:第1种连续投喂组,整个试验期间不间断,为对照组。第2种投喂3 d后饥饿1 d;第3种连续投喂7 d后饥饿1 d。3种方式分别以R0、R1/3和R1/7表示。投喂鲜杂饲料∶茨粉=6∶1。每组做3个重复。实际水体积为120 L。模拟实际养殖条件下换水量,每天换水两次,每次换水量为实际水体积的3/4,水温保持(25±1)℃。每日投喂两次,日投饵量为体重4%~7%。试验时间为50 d,每天观察并记录鱼的摄食、残饵及死亡情况。试验用水为前1 d曝气后的自来水。
1.3 消化酶液制备
试验结束时每个水族箱随机选取5尾革胡子鲶,将其置冰盘毁髓处死,解剖分离肝脏、胃、肠道,用去离子水冲洗干净,用剪刀剪成小块。肝脏、胃和肠道称重后加入9倍的生理盐水,研磨均匀,将研磨液低温离心(4 ℃、4 000 r/min,15 min),制成组织匀浆液。对肝脏、前肠、中肠和后肠组织匀浆液进行消化酶活性的测定。
1.4 酶活性测定
蛋白酶活性采用福林酚试剂法测定;淀粉酶和脂肪酶活性均采用南京建成生物工程研究所试剂盒测定;总蛋白(TP)含量采用半自动化测定仪测定,方法为双缩脲法。
1.5 数据分析
所得的原始数据用Excel 2003和SPSS11.0软件处理。利用方差分析检验各组间差异,若差异显著,采用Duncan's法作多重比较。
2 试验结果
2.1 不同处理组对革胡子鲶蛋白酶活性的影响(见表1)
由表1可以看出,R0、R1/3和R1/7组胃、后肠和肝脏中蛋白酶活性差异不显著(P>0.05),但前肠和中肠内蛋白酶活性存在差异,其中R0组的前肠蛋白酶活性最高,R1/7组最低(P<0.05)。中肠R1/7组蛋白酶活性最高,R1/3组最低(P<0.05)。

表1 不同处理组对革胡子鲶蛋白酶活性的影响(U/mg蛋白)
2.2 不同处理组对革胡子鲶淀粉酶活性的影响(见表2)
由表2可以看出,R0组、R1/3组和R1/7组胃、前肠、中肠、后肠和肝脏内淀粉酶活性差异不显著(P>0.05),其中,R1/7组的前肠、中肠和后肠淀粉酶活性最高。
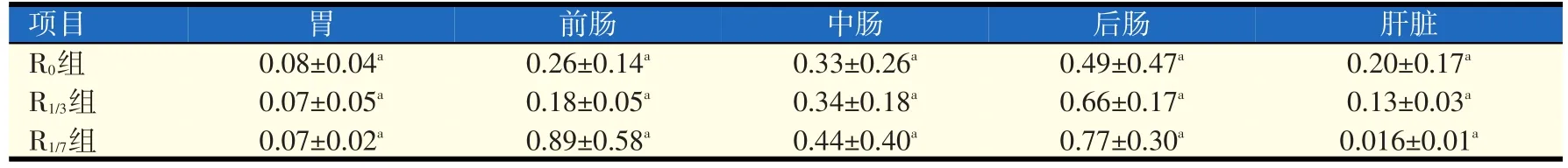
表2 不同处理组对革胡子鲶淀粉酶活性的影响(U/mg蛋白)
2.3 不同处理组对革胡子鲶脂肪酶活性的影响(见表3)
由表3可以看出,R0、R1/3和R1/7组对胃、前肠和肝脏脂肪酶活性影响,其中R0的脂肪酶活性在胃、前肠和肝脏中都是最高的。而R0、R1/3和R1/7组中肠、后肠中脂肪酶活性差异不显著(P>0.05),其中R0中后肠脂肪酶的活性最高,R1/3的中肠脂肪酶活性最高。
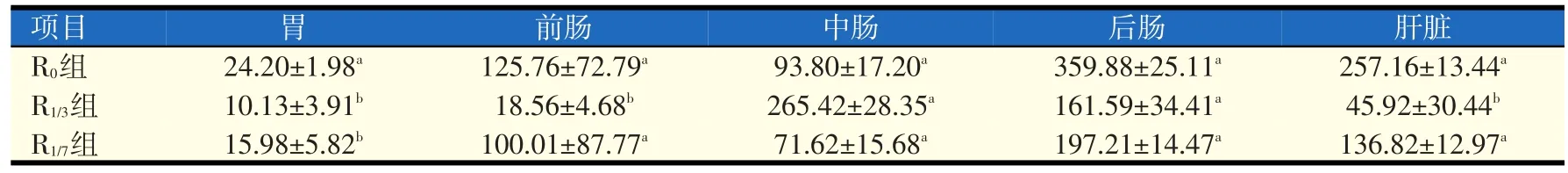
表3 不同处理组对革胡子鲶脂肪酶活性的影响(U/mg蛋白)
3 讨论
动物产生补偿生长的生理机制主要有3种:①饥饿使动物代谢水平降低,但恢复进食时,较低的代谢水平能持续一段时间,这种代谢支出的降低使动物用于生长的能量增加,从而提高了食物转化率,出现补偿性生长;②饥饿后恢复喂食时,动物体内将立即进行大量的合成作用,代谢水平将迅速升高,不可能通过降低代谢水平而明显改善食物转化率,补偿生长的产生是动物在恢复生长中食欲增强,大幅度提高摄食水平实现的;③补偿生长是摄食率和食物转化率同时提高的结果。消化酶主要是消化腺和消化系统分泌的起消化作用的酶类。在消化酶中,依照消化对象的不同而大致划分蛋白酶、淀粉酶、脂肪酶和纤维素酶等几种。消化酶的活力高低决定着对营养物质消化吸收的能力,从而决定其生长发育速度,同时也能反应出水产动物的变化或利用饲料成分的能力。
3.1 不同处理组对革胡子鲶蛋白酶活性的影响
钱云霞对养殖鲈的研究发现,饥饿期间消化道没有食物蠕动的机械刺激,因而蛋白酶的分泌量下降,蛋白酶的活性降低。付世建等对南方鲇幼鱼进行长期饥饿后,发现其胃和肝脏的组织结构变化,从而导致这些器官的消化酶分泌量降低。此外,Yufera等也提出了类似的推论,即长期饥饿后仔鱼呈现出广泛的组织学衰退,特别是消化道和附属腺体。从本试验结果中可以看出,各投喂组的胃、后肠和肝脏中蛋白酶差异不显著,说明隔3 d停喂1 d和隔7 d停喂1 d与不间断投喂对革胡子鲶胃、后肠和肝脏中蛋白酶活性的变化没有影响。分析原因,可能是停喂时间较短,饥饿并未对其消化器官组织结构产生影响。而在前肠内蛋白酶活性随着间隔天数的增加而呈现下降的趋势。隔3 d停喂1 d投喂组与不间断投喂组的中肠蛋白酶活性差异不显著,但当隔7 d停喂1 d时,其饥饿并没有引起消化酶活性的下降,反而使消化酶活性高于对照组,说明短期饥饿可以一定程度上诱导中肠内消化酶活性的升高,这与乔秋实等研究结果相似。
3.2 不同处理组对革胡子鲶淀粉酶活性的影响
王燕妮等通过对鲤鱼饥饿与淀粉酶活性关系的探讨,发现总的趋势是饥饿后鲤鱼的淀粉酶活性大幅上升,而恢复进食后则有所下降。樊启学等研究结果表明,饥饿再投喂后,翘嘴鲌幼鱼消化酶活性迅速升至显著高于饥饿前水平,然后再逐步恢复正常。郑曙明等进行了饥饿和再投喂对翘嘴鲌幼鱼消化酶活性影响的研究,研究结果表明淀粉酶活性在饥饿时间较短时呈现下降趋势,持续饥饿时间增加后则开始上升,恢复生长后酶活性均上升。从本次试验结果中可以看出,各投喂组的各消化器官中淀粉酶活性差异不显著,说明隔3 d停喂1 d和隔7 d停喂1 d与不间断投喂对革胡子鲶胃、后肠和肝脏中淀粉酶活性的变化没有影响。造成这种现象的原因,可能是当水产动物饥饿时首先消耗的不是糖原,优先动用脂肪补充能量,而本试验饥饿时间较短,鱼体并未开始出现由于大量消耗糖原,从而导致淀粉酶活性增加,糖原以转化淀粉作为补充供能物质的现象。
3.3 不同处理组对革胡子鲶脂肪酶活性的影响
Orhan等通过对海鲷鱼幼鱼肠道内脂肪酶的测定,发现饥饿能改变肠道内脂肪酶活性。罗海忠等发现,饥饿开始后鮸鱼幼鱼各器官中的脂肪酶活性下降;随饥饿时间延长,脂肪酶活性出现上升现象。区又君等研究结果表明,在饥饿过程中千年笛鲷幼鱼体内脂肪酶活性下降明显,恢复投喂后,脂肪酶活性与饥饿前相比都有所上升,脂肪酶活性总体上高于对照组水平。从本试验结果中可以看出,各投喂组的中肠和后肠脂肪酶活性差异不显著,说明隔3 d停喂1 d和隔7 d停喂1 d与不间断投喂对革胡子鲶中肠和后肠脂肪酶活性的变化没有影响。但对胃、前肠和肝脏脂肪酶活性有影响,并且胃内脂肪酶活性随着间隔投喂的出现呈现下降的趋势。
4 结论
用间歇性投喂方式投喂革胡子鲶,用间歇性投喂方式投喂革胡子鲶,3种方式对革胡子鲶各消化器官消化酶有影响。隔7 d投喂1次的中肠蛋白酶活力有显著的升高,并且肠道淀粉酶活力是最高的。综合考虑,隔7 d停喂1次喂养革胡子鲶获得的试验效果较理想。在革胡子鲶实际养殖生产中。综合考虑,隔7 d停喂1次喂养革胡子鲶获得的试验效果较理想。在革胡子鲶实际养殖生产中,可以隔1周停喂1次来投喂革胡子鲶,既能保证革胡子鲶消化酶的稳定,又能起到节约饲料,提高饵料利用率,加强鱼体免疫力,提高养殖产量的作用。
猜你喜欢
大连海洋大学学报(2022年5期)2022-11-17
当代水产(2021年9期)2021-12-02
当代水产(2021年8期)2021-11-04
医学前沿(2021年18期)2021-04-14
今日农业(2020年15期)2020-12-15
贵州农业科学(2019年7期)2019-08-03
中国继续医学教育(2015年1期)2016-01-06
小天使·二年级语数英综合(2015年5期)2015-05-15
小说月刊(2015年5期)2015-04-19
语文世界(小学版)(2014年10期)2015-04-02
