
岷江上游干旱河谷荒坡植物群落的β多样性分析
2013-02-05 01:23杨子松权秋梅
中国水土保持 2013年1期
杨子松,权秋梅
(阿坝师范高等专科学校 化学化工与生命科学系,四川 汶川623002)
(责任编辑 徐素霞)
β 多样性也被称为物种替代速率(species replacement rate)、物种周转速率(species turnover rate)和生物变化速率(rate of biotic change),指的是在地貌、土壤和干扰等各种环境因子的控制下,不同生境群落的物种组成情况差异以及它们沿环境梯度的更替程度[1]。测定植物群落的β 多样性,不仅能够体现出生境被物种分隔的程度并进行不同地段的生境多样性比较,而且还有助于对物种稀有和濒危机制的了解[1]。
选择β 多样性指数为指标分析岷江上游干旱河谷荒坡植物群落,能够为岷江上游干旱河谷荒坡的植被保护,特别是“5·12”汶川大地震破坏区域的植被恢复提供一定的理论依据和实践指导。
1 研究区概况
青藏高原和四川盆地之间的岷江上游干旱河谷,位于东经103°10'—103°54'、北纬31°15'—32°32'范围内。该区域年均温仅为11.2 ℃,年均降水量为493 mm,蒸发量为1 332 mm。其特殊的气候条件所孕育的植被,常常呈现出一道道旱生半荒漠景观。常见草本植物主要有高山韭(Allium sikkimense)、细柄草(Capillipedium parviflorum)、披针苔草(Carex lanceolata)、阴地蒿(Artemisia sylvatica)、垫状卷柏(Selaginella pulvinata)、汶川景天(Sedum wenchuanense)、岷江百合(Lilium regale Wilson)等,灌丛植物有白刺花(Sophora viciifolia)、小马鞍叶羊蹄甲 (Bauhinia faberi var.microphylla)等[2,4]。
2 研究方法
2.1 野外调查
2007年6月,在岷江上游干旱河谷荒坡植被生物量最大的时期,于岷江上游及其主要支流黑水河和杂谷脑河两岸,对10 个10 m ×10 m 旱生半荒漠的荒坡样地的海拔、坡向、坡度等生境特征和相应植物种群的盖度、多度、高度、株(丛)数、丛径等种群特征进行了调查。沿海拔梯度对相邻样地的物种数量与个体数量的变化情况进行统计,所得结果如表1 所示。
2.2 植物群落β 多样性测度
β 多样性测度方法比较多,一般可以分成两类,一是用二元属性数据测度,二是用数量数据测度。本研究中,二元属性数据测度方面选取了Cody 指数βc和Whittaker 指数βws来反映β 多样性沿环境梯度的变化,用Jaccard 指数CJ和Sorenson 指数CS来反映群落或生境间的β 多样性。为了充分考虑各个物种的个体数量及其相对多度,有效预防二元属性数据测度β 多样性过程中对稀疏种的作用作出过高估计导致不合理结论,本文选用了Bray-Curtis 指数CN和Morisita-Horn 指数Cmh这两种数量数据测度方法[1,5]。
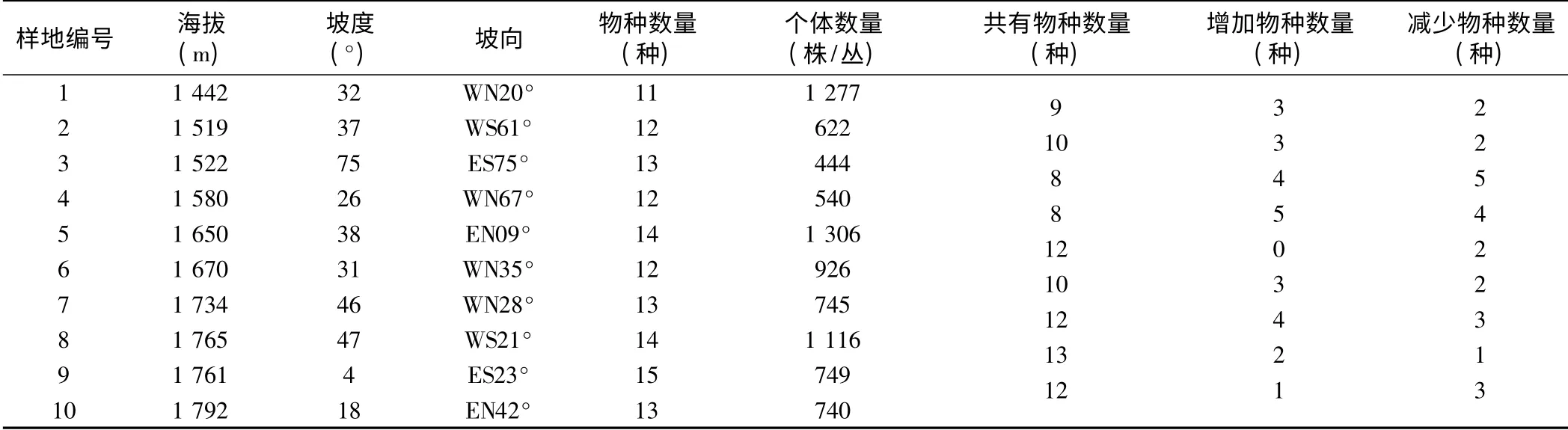
表1 沿海拔梯度按相邻样地统计的调查结果
2.2.1 二元属性数据测度
Cody 指数βc、Whittaker 指数βws、Jaccard 指数CJ、Sorenson 指数CS的计算公式分别为
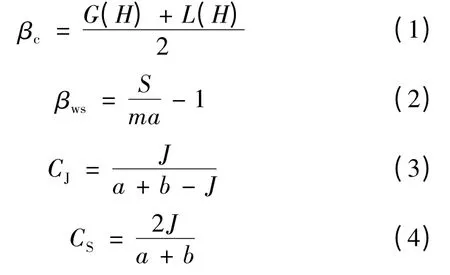
上四式中:G(H)、L(H)分别为沿环境梯度H 增加或减少的物种数量;S 为所研究系统里面所记录到的物种的总数;ma 为各样地平均物种数量;a、b 分别为样地A、样地B 里面出现的物种数量;J 则为A、B 两个样地所共有的物种数量。
2.2.2 数量数据测度
Morisita-Horn 指数Cmh、Bray-Curtis 指数CN的计算公式分别为
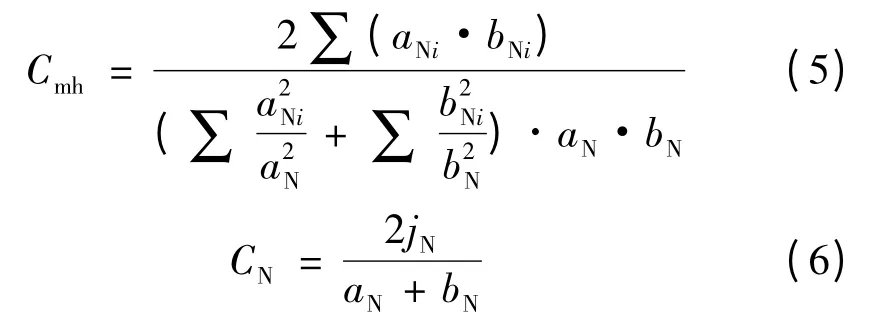
上二式中:aN、bN分别为样地A、样地B 的物种数量,aNi、bNi分别为样地A、样地B 中第i 种的个体数量,jN则是A、B 两样地共有种中个体数目较小者之和,即:jN=∑min(jNa+jNb)。
3 结果与分析
3.1 物种数量和个体数量的变化
从表1 不难看出,随海拔升高和环境梯度的变化,岷江上游干旱河谷荒坡植物群落中的物种数量差异不大,总个体数变化规律不明显。这可能是在岷江上游干旱河谷特殊的气候条件下,荒坡集中分布在海拔1 400—1 800 m 这样一个相对狭窄的范围内,海拔变化所导致的生态环境差异比较小,还没能对荒坡植物的分布和生长产生重要影响。至于为什么样地1、5、8中物种的个体数目显著高于其他样地,应该是其生境条件明显优于其他样地所致。
从样地间的差异上看,随着海拔的增高,各样地之间共有种的数目总体上呈增加趋势但又变化不大,新增加物种数和减少物种数呈现出先增多后又减少的不规律现象,这反映出样地间的差异并不显著。可见,在岷江上游干旱河谷荒坡,生境梯度的变化是非均匀的,样地之间的差异并不明显。
3.2 沿海拔梯度的其他样地与样地1 间植物群落的β 多样性
对其他各样地与样地1 间的β 多样性进行测定,所得结果见表2。不难发现,样地7 与样地1 之间的βws=0.416 7、βc=4.0 值最大,对应的CN=0.749 3、Cmh=0.913 3、CJ=0.388 9、CS=0.560 0;样地9 与样地1 之间的βws和βc最小,分别为0.153 8 和2.0,对应的CN、Cmh、CJ、CS分别为0.328 2、0.427 8、0.611 1、0.758 6。一般地,βws和βc值越大意味着群落间的差异性就越大。据此认为,样地7 与样地1 间植物群落组成和生境差异较大,而样地9 与样地1 间的差异性则最小。
3.3 沿海拔梯度相邻样地植物群落的β 多样性
岷江上游干旱河谷荒坡10 个样地沿海拔梯度相邻样地植物群落的β 多样性测度结果如表3 所示。由表3 结合表1 可以看出:
(1)岷江上游干旱河谷荒坡植物群落的β 多样性及其指数波动幅度都比较小。βws和βc明显非常小,表明岷江上游干旱河谷荒坡植物群落间的相似性较大,相异性较小。βws在样地6 与样地5 之间特别小,为0.076 9,其他也都介于0.1 ~0.4 之间,平均值仅为0.216 1,这与荒坡植物群落的物种丰富度比较小是一致的(10 个样地物种数在11 ~15 之间,平均值也仅为12.9),表明荒坡植物群落的物种组成的差异比较小。1.0≤βc≤4.5,平均值也仅为2.722 2,说明岷江上游干旱河谷荒坡植物群落物种的更替速率比较小,植物群落相对稳定。
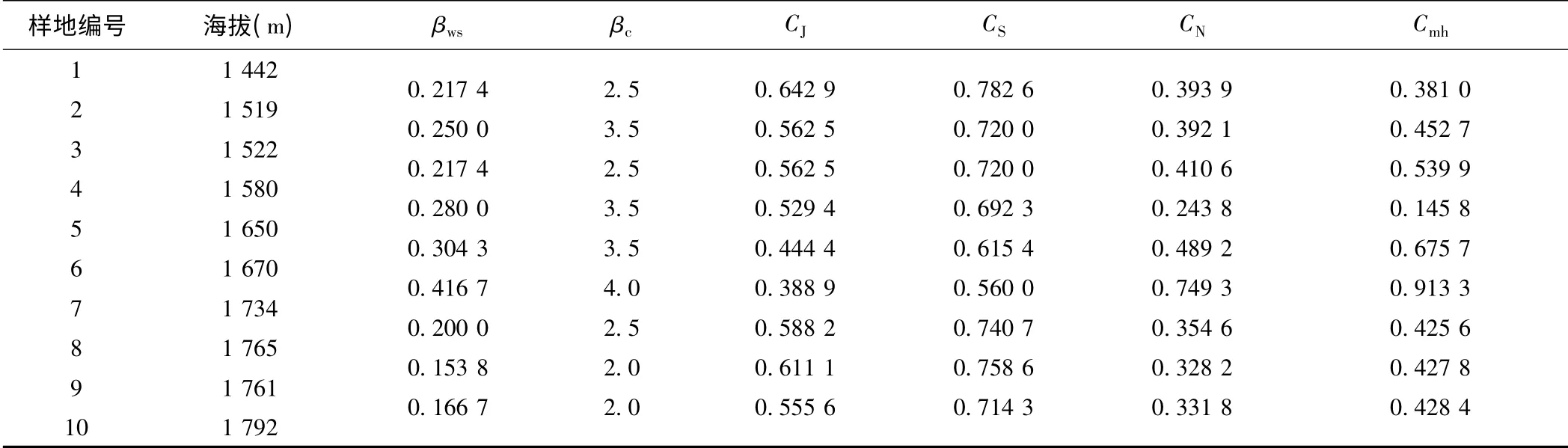
表2 岷江上游干旱河谷荒坡沿海拔梯度其他样地与样地1 间植物群落的β 多样性
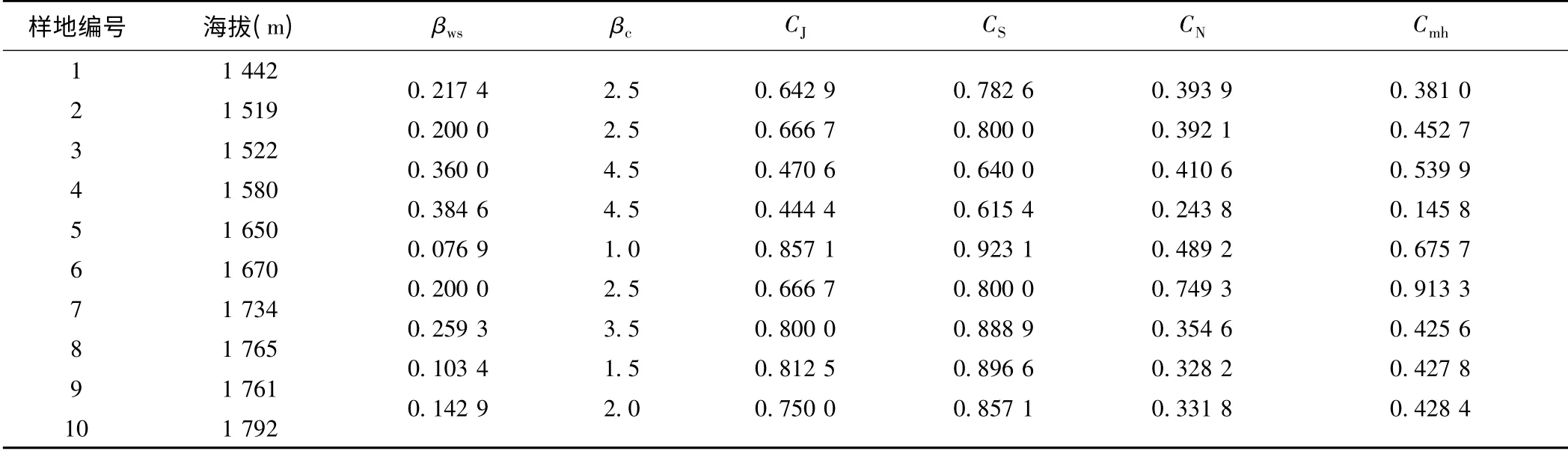
表3 岷江上游干旱河谷荒坡沿海拔梯度相邻样地植物群落的β 多样性
CJ、CS、CN和Cmh普遍较大,其中0.444 4≤CJ≤0.857 1,平均值达到0.679 0;0.615 4≤CS≤0.923 1,平均值达到了0.800 4;0.243 8≤CN≤0.749 3,平均值为0.410 4;0.145 8≤Cmh≤0.913 3,平均值为0.487 8。CJ、CS、CN和Cmh这4 个指数重点考察的是群落间的共有种,共有种越多其值越大,其β 多样性越小。可见,岷江上游干旱河谷荒坡植物群落间共有种较多,物种更替速率较小,群落β 多样性较小。
(2)总体而言,岷江上游干旱河谷荒坡植物群落的β 多样性随着海拔的增加而呈现减小的趋势。样地3(海拔1 522 m)至样地5(海拔1 650 m)间的CJ和CS较小,但是βc和βws较大。其中,样地3、4 之间和样地4、5 之间的βc都达到了最大值4.5;样地5、4 间的βws达到了最大值0.384 6,并且其CJ也同时达到了最小值0.444 4,CS达到最小值0.615 4。这意味着,在岷江上游干旱河谷荒坡,海拔1 522—1 650 m 之间的植物群落多样性差异最显著,更替速率最快。
(3)随海拔升高,不管是其他样地与样地1 之间还是相邻样地之间,虽然作为数量数据的相似性系数的CN、Cmh值的变化并不像二元属性数据的βws、βc和CJ、CS变化那样有着更强的规律性,有时甚至出现不规则跳跃式变化,但是整体上看,CN、Cmh值的总体变化规律与βws、βc值的变化规律大致上是相反的,与CJ、CS值的变化规律则大致相似。这表明,在岷江上游干旱河谷荒坡,Jaccard 指数CJ、Sorenson 指数CS、Bray- Curtis 指数CN、Whittaker 指数βws、Morisita-Horn 指数Cmh和Cody 指数βc都能较好地描述植物群落的β 多样性,但二元属性数据的βws、βc和CJ、CS更为适合。
4 结 语
(1)α 多样性与β 多样性是评价生物多样性最为常用的两个指数。α 多样性主要关注局域均匀生境下的物种数目,反映物种或其个体数量的变化,是生物群落内的多样性(within-habitat diversity)。β 多样性则反映不同地段群落物种构成的差异和环境的异质性,是群落间的多样性(between-habitat diversity)。β 多样性与α 多样性一起构成了总体多样性(overall diversity)或一定地段的生物异质性(biotic heterogeneity)[1]。当然,由于关注点的差异,人们对β 多样性的测度比α 多样性的测度工作少得多,但这并不意味着β 多样性测度的工作不重要,意义没有α 多样性测度大,这种现象需要引起高度的重视。α 多样性较高群落的β 多样性不一定高,α 多样性较低群落的β 多样性也不一定就低[1]。但在岷江上游干旱河谷荒坡,植物群落的α多样性和β 多样性保持一致,都非常低[1,6]。
(2)岷江上游干旱河谷荒坡历来都是该地区植被恢复的重点区域。本文的研究表明,虽然干旱河谷荒坡的植物种类和数量非常贫乏,但是其植物群落的生境差异并不明显,其物种组成和结构差异小且保持着相对稳定。植被恢复过程中,在相关策略和植物种类的选择上应该更加谨慎,这对岷江上游干旱河谷植被的保护和恢复重建,特别是对“5·12”汶川大地震破坏区域的植被恢复具有十分重要的意义。
[1]马克平,刘灿然,刘玉明.生物群落多样性的测度方法Ⅱ.β 多样性的测度方法[J].生物多样性,1995,3(1):38-43.
[2]陈泓,黎燕琼,郑绍伟,等.岷江上游干旱河谷灌丛群落种-面积曲线的拟合及最小面积确定[J].生态学报,2007,27(5):1818-1825.
[3]杨兆平,常禹.我国西南主要干旱河谷生态及其研究进展[J].干旱地区农业研究,2007,25(4):90-93,99.
[4]关文彬,冶民生,马克明,等.岷江干旱河谷植被分类及其主要类型[J].山地学报,2004,22(6):679-686.
[5]王贵霞,李传荣,齐清,等.泰山油松群落β 多样性研究[J].山东农业大学学报:自然科学版,2004,35(3):347-351.
[6]张金屯,陈廷贵.关帝山植物群落物种多样性研究Ⅱ.统一多样性和β 多样性[J].山西大学学报:自然科学版,2002,25(2):173-175.
猜你喜欢
散文诗世界(2021年12期)2021-12-17
小哥白尼(神奇星球)(2021年3期)2021-07-22
小哥白尼(神奇星球)(2021年4期)2021-07-22
东坡赤壁诗词(2020年4期)2020-09-02
东坡赤壁诗词(2019年5期)2019-11-14
Zoological Research(2019年3期)2019-06-28
岷峨诗稿(2019年4期)2019-04-20
农家科技下旬刊(2018年10期)2018-12-15
阿来研究(2018年1期)2018-07-10
乡村地理(2018年4期)2018-03-23
