
阳生与阴生环境梓树叶片的比较解剖学研究
2012-12-26 08:58李永林金日权王迪涵李春姣吴东梅
东北师大学报(自然科学版) 2012年1期
李永林,金日权,王迪涵,李春姣,吴东梅,魏 健
(1.东北师范大学生命科学学院,吉林 长春 130024;
2.金亨稷师范大学生物系,平壤 99903;
3.朝鲜金日成综合大学生物系,平壤 99903)
阳生与阴生环境梓树叶片的比较解剖学研究
李永林1,2,金日权3,王迪涵1,李春姣1,吴东梅1,魏 健1
(1.东北师范大学生命科学学院,吉林 长春 130024;
2.金亨稷师范大学生物系,平壤 99903;
3.朝鲜金日成综合大学生物系,平壤 99903)
采用光学显微技术,对不同生长环境的梓树CatalpaovataG.Don叶片进行了比较解剖研究.结果表明:梓树的阳生叶比阴生叶大得多,为1.94~2.64倍;阳生叶的栅栏组织厚度是阴生叶的1.96倍左右.阳生叶的气孔微下陷,有利于减少水分蒸腾;阴生叶的气孔大部分微上拱.阳生叶的上表皮比阴生叶厚,为1.91倍左右;阳生叶的上表皮外角质层比阴生叶的厚,为1.8倍左右.梓树阳生叶上表皮的外切向面具较厚的角质层,且表面较糙,有利于减少强光灼伤,增强抗旱能力;阴生叶的角质层表面光滑,体现了阴生叶的特点.梓树阳生叶的叶脉机械组织较发达,有利于水分的输导;阴生环境下梓树机械组织不发达,代表了阴生环境下植物的特点.不同环境条件下生长的梓树,演化出明显不同的解剖结构,验证了“环境决定植物结构的建成”理论.
梓树;光学显微技术;阳生;阴生;演化结构
梓(CatalpaovataG.Don),隶属于紫葳科Bignoniaceae梓属Catalpa,别名为花楸,原产于中国,分布于长江流域及其以北地区,如东北三省、河北、山西、陕西、河南、山东、江苏、安徽、浙江、江西、湖南、湖北、四川、云南、贵州、广东及台湾等省和地区,在朝鲜、日本也有分布,多栽培于村庄附近及公路两旁,野生的已不多见[1-4].
梓树冠宽阔,枝条开展,树皮灰褐色、浅纵裂;喜光,稍耐阴,耐寒,适生于温带地区,在暖热气候下生长不良,深根性.叶对生,有时轮生,具长柄,宽卵形或近圆形,先端基部圆形或心形,常有3~5浅裂;叶脉基部有1~6个紫色腺点.圆锥花序,顶生,花冠淡黄色,内有黄色线纹及紫色斑点.蒴果细长如豇豆,常经冬不落[5].种子扁平,两端生有丝状毛丛.
梓树为速生树种,喜深厚、肥沃、湿润的土壤,不耐干旱和瘠薄地,能耐轻盐碱土,抗污染性较强.梓树树体端正,冠幅开展,叶大荫浓,春夏黄花满树,秋冬荚果悬挂,是具有一定观赏价值的树种[6-7],常作为行道树、绿化树种[8].嫩叶可食;根皮或树皮、果实、木材、树叶均可入药,能清热、解毒.木材材质轻软细密,纹理直行,柔而耐腐,花纹美观,适于作家具、建筑、室内装饰、乐器和工艺美术等用材[9],也可作为保温隔热材料.
目前在梓的繁殖、播种、组织培养等研究方面已有报道[10-12],本文通过对梓的阳生、阴生叶解剖结构的比较研究,探讨了其结构与环境的关系,为全面认识植物对环境的生态适应性提供了基础资料.
梓树花粉母细胞呈半圆形排列,小孢子四分体属四面体型,绒毡层属分泌型,花粉粒为复合花粉.小孢子各发育时期与花蕾大小之间存在较密切的相关性[13].
梓树的外绒毡层是由次生周缘层发育而来,内绒毡层是由紧靠造胞组织的一层药隔细胞发育而来,而且内绒毡层细胞在辐射向上伸长并伸入药室的过程中形成“拟胎座”.在梓树的花药中,紧靠表皮的一层细胞,随着其他中层细胞的破坏消失最终也破坏消失,因此,我们认为该层细胞仍属中层,在梓树中不形成药室内壁.
子叶节区下部具有中始式四原型管状中柱,在子叶节区中部的原生木质部处一分为二,逐渐形成具有8枚外韧维管束的中柱结构;在子叶节区上部,组成短轴的2枚维管束各自一分为三,逐渐形成真中柱的雏形,与上胚轴顶端分生性组织分化形成的内始式真中柱相连,至此,子叶节区发育完成.子叶为单隙三迹[14-15].
1 材料与方法
1.1 材料
选择阳生环境和阴生环境下生长的梓树作为实验材料,材料采自长春市东北师范大学校园内.2种材料取其当年新生的茎、叶,并尽量取其中间部位.长春市属温带大陆性季风气候,年平均气温4.8℃,年平均降水量522~615mm.
1.2 实验方法
(1)取材与固定.选择上述2种不同生态环境下生长的梓树作为标本和供试材料,用FAA固定液进行固定,固定时间不少于48h.
(2)脱水.酒精梯度逐级脱水,每个浓度梯度脱水1.5h;最后,100%乙醇处理2.5h,然后更换无水乙醇再处理1h,对材料彻底脱水.
(3)透明.利用不同比例的无水乙醇和二甲苯的混合液,并不断增加二甲苯的比例,逐级透明.每级透明时间为1h.
(4)浸蜡.先用低熔点的切片石蜡(熔点52℃~54℃)在30℃下浸蜡24h,然后用由蜂蜡(熔点58℃~60℃)和低熔点切片石蜡按1∶9的质量比融合在一起的混合石蜡在温箱中60℃保温浸蜡,每2h换一次混合石蜡,重复进行两次,共6h.
(5)包埋.材料浸蜡结束后马上用镊子转移到装有液态上述混合石蜡的小纸盒中,并迅速冷却.材料的轴性系统要垂直于盒底部,以便修整蜡块作横切材料.
(6)切片.根据材料在蜡块中的位置和方向,将蜡块修成梯形,利用切片机进行切片.
(7)展片及粘片.在洁净的载玻片上均匀涂上少许蛋白甘油,将蜡带小心地放在50℃的水面上,待蜡带展平后,用涂有蛋白甘油的载玻片将之捞起,使蜡带光面紧贴载玻片,然后在35℃的温箱中烘干,使蜡带粘附在载玻片上.
(8)脱蜡和染色.二甲苯脱蜡后采用番红亮绿对染法染色.先用纯二甲苯染色,然后进行梯度染色,每个浓度梯度停留2~3min.
(9)封片及观察.用加拿大树胶封片后,将切片放入35℃温箱中烘干.数日后可进行观察和光学显微摄影.
2 实验结果
梓叶比较大,广卵形或圆形,叶长宽几乎相等,叶上端常有3浅裂,阳生叶比阴生叶要大.
2.1 梓阳生叶的形态解剖结构
梓叶片由表皮、叶肉和叶脉3部分组成,阳生叶的厚度为209.26μm(见图1A).
2.1.1 表皮的形态结构
表皮由一层细胞组成,分为上、下表皮.上表皮细胞外切向面具较厚的角质层且粗糙;下表皮角质层不明显,气孔微下限,有利于减少水分的蒸腾(见图1E).
2.1.2 叶肉的形态结构
梓阳生叶的叶肉分为栅栏组织和海绵组织.栅栏组织细胞呈长柱状,内含丰富的叶绿体;海绵组织由不规则细胞组成,且排列较疏松.栅栏组织中含有明显较大的异形胞,异形胞中含有较多水分,是植物抗旱的一种特殊结构(见图1C).
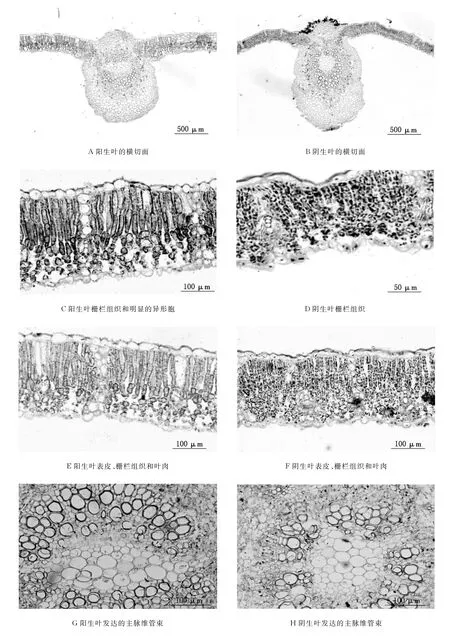
图1 梓阳生叶与阴生叶形态解剖结构
2.1.3 叶脉的形态结构
叶脉由维管束和伴随的机械组织组合而成(见图1G).机械组织发达,有利于支持作用,能将叶片伸展于空间.叶脉上的木质部导管较发达,有很强的输导能力,在基本组织中存在着大量的晶体细胞.
2.2 梓阴生叶的形态解剖结构
梓阴生叶片亦由表皮、叶肉和叶脉3部分组成,厚度为128.35μm(见图1B).
2.2.1 表皮的形态结构
梓阴生叶表皮亦由一层细胞组成,分为上、下表皮.上表皮细胞排列形状较规则,外切向壁具较厚角质层,且光滑.气孔微上拱,具有明显的孔下室,通气组织发达,为典型的阴生结构(见图1F).
2.2.2 叶肉的形态结构
梓阴生叶的叶肉组织不能明显地区别为栅栏组织和海绵组织(见图1F).栅栏组织细胞呈短柱状,叶绿体格外显著;海绵组织细胞排列疏松.叶肉中不存在较大的异形胞,显示了阴生结构的特征(见图1D).
2.2.3 叶脉的形态结构
梓阴生叶的叶脉由维管束和机械组织组成(见图1H).机械组织不发达,支撑能力较弱,主脉中具有一个维管束.木质部导管退化,输导能力较弱.阴生叶也含有较多的晶体细胞.
3 讨论
叶是进行植物光合作用和蒸腾作用的主要器官,具有较大的表面积,与周围环境的接触十分紧密,植物对环境的反应较多地体现在叶的形态结构上,因此,叶的结构对生态环境条件变化的反应最为敏感[10].本实验的结果表明,梓阳生叶与阴生叶在外部形态、叶片内部结构和叶表皮上存在较大差异.具体来说,在叶的大小、叶厚度、叶肉栅栏组织的发达程度、气孔类型、表皮厚度、表皮角质层的厚度和形状、主脉横切面形状、导管列数等方面的演化结构特征存在差异.
梓树阳生叶比阴生叶大得多,为1.94~2.64倍,阳生叶的栅栏组织厚度是阴生叶的1.96倍左右.阳生叶的气孔微下陷,有利于减少蒸腾作用;而阴生叶的气孔大部分微上拱.阳生叶的上表皮是阴生叶的1.91倍左右.这些特征均是典型的阳生、阴生环境的植物叶特征.
梓阳生叶的上表皮具有较为发达的角质层,阳生叶的上表皮外角质层比阴生叶厚得多,为1.8倍左右,而且阳生叶的上表皮外角质层表面较糙,有利于减少强光灼伤,增强抗旱能力;阴生叶的角质层表面光滑,体现了阴生叶的演化结构特点.
在叶的横切面上,阳生叶主脉的形状为圆形或近圆形,而阴生叶主脉的形状为长椭圆形.叶脉的主要功能有两种:一个是运输功能,另一个是机械功能.
综上所述,阳生叶比阴生叶大得多、重的多.阳生叶叶脉的机械组织较发达,有利于支持作用和水分的输导;而阴生叶机械组织一般不发达,代表了阴生环境下植物的特点,生态环境导致了阴生叶脉的演化结构发生了变化.本文验证了“环境决定植物结构的建成”理论的正确性[16].
[1] 陆静梅,李建东,张洪芹.吉林西部草原区7种耐盐碱双子叶植物结构研究[J].应用生态学报,1996,7(3):283-296.
[2] 柯曙华,王传贵,刘秀梅.梓树材性及用途[J].安徽农业大学学报,1994(3):611-612.
[3] 许艳芳,王淑娟.梓树与楸树[J].中国花卉报,2009(12):107-109.
[4] 赵红英,王鑫晓,高江华,等.出土梓木形态结构及物理力学性质研究[J].北京林业大学学报,2008(9):87-89-
[5] 宫玉婷,阿不都拉·阿巴斯.梓树果实有效成分的初步研究[J].食品科学,2008(6):.10-13.
[6] 赵曦阳,王军辉,张金凤,等.梓树属种子表型性状和发芽特性的研究[J].西北农林科技大学学报,2008(12):108-110.
[7] 王改萍,杨红宁,倪果果,等.楸树等4种梓属树种花粉离体培养条件[J].植物资源与环境学报,2009(2):206-207.
[8] 张永忠.梓树的播种育苗技术[J].防护林技术,2006(3):204-206.
[9] 占玉芳,蔺国菊.干旱、半干旱区梓树的繁育技术及其利用[J].防护林技术,2005(1):110-112.
[10] 建德蜂,陈刚,王中武,等.梓树的播种繁殖及养护管理[J].黑龙江农业科学,2009(6):28-31.
[11] 王瑞福,赵颖,乔勇进.楸树的良种壮苗繁殖技术探讨[J].内蒙古林业科技,2003(3):111-113.
[12] 于永明,王军辉,马建伟,等.楸树砧木——梓树播种育苗技术[J].科技创新导报,2009(3):19-21.
[13] 全雪丽,刘继生,金大能,吴松权.梓树花药发育解剖学研究[J].辽宁林业科技,2011(2):23-27.
[14] 闫桂华,王立军,聂小兰.梓树幼苗初生维管系统的解剖学研究[J].植物研究,2011,31(3):261-264.
[15] 马德滋,任笠坤.梓树CatalpaovataDon花药[J].宁夏农学院学报,1988(2):88-91.
[16] 陆静梅.松嫩平原4种盐生植物根的结构研究[J].生态学报,1998,18(3):336-338.
Comparison of anatomical research onCatalpaovataG.Don’s leaves in sun and shade environments
LEE Yong-rim1,2,KIM I1-kwon3,WANG Di-han1,LI Chun-jiao1,WU Dong-mei1,WEI Jian1
(1.School of Life Sciences,Northeast Normal University,Changchun 130024,China;
2.Department of Biology,Kim Hyong Jik University of Education,Pyongyang 99903,Korea;
3.Department of Biology,Kim I1-sung University,Pyongyang 99903,Korea)
CatalpaovataG.Don's leaves in different environments are conducted the comparison of anatomical research through applying optical microscopy.Results show that sun leaves ofCatalpa ovataG.Don are much larger than shade leaves(about 1.94~2.64times).Palisade tissue's thickness of sun leaves is about 1.96larger than shade leaves.Stoma of sun leaves slightly subsides,which is beneficial to reduce transpiration.Most stoma of shade leaves slightly arch.Upper epidermis of sun leaves is about 1.91thicker than shade leaves.Outer cuticle of sun leaves'upper epidermis is about 1.8 thicker than shade leaves.Outer tangential surface of upper epidermis ofCatalpaovataG.Don's sun leaves has relatively thick cuticle and the surface is rough,which is beneficial to reduce the burning of glare and enforce the ability of resisting drought.Surface of sun leaves'cuticle is smooth,which illustrates the characteristic of shade leaves.Leaf vein's machine organization ofCatalpaovataG.Don is more developed,which is helpful to the transportation of water.Machine organization ofCatalpa ovataG.Don in shade environment is not developed,which represents characteristic in shade environment.CatalpaovataG.Don in different environments evolved the obvious different anatomical structures,which proves the theory that environment decides the construction of plant structures.
CatalpaovataG.Don;optical microscopy;sun environment;shade environment;evolved structures
Q 944.1
180·5125
A
1000-1832(2012)01-0123-05
2011-04-12
吉林省科技发展计划项目(20080241).
李永林(1972—),男,讲师,主要从事植物学研究;通讯作者:魏健(1980—),男,博士,讲师,主要从事结构植物学研究.
方 林)
猜你喜欢
动漫界·幼教365(大班)(2021年11期)2021-11-20
毛纺科技(2021年8期)2021-10-14
作文周刊·小学二年级版(2021年40期)2021-01-05
发明与创新(2020年26期)2020-06-27
伴侣(2020年4期)2020-04-27
伴侣(2020年2期)2020-04-13
花卉(2020年22期)2020-01-08
苏州杂志(2019年5期)2019-12-03
小学生学习指导(低年级)(2019年9期)2019-09-25
诗歌月刊(2017年10期)2018-01-28
