
果蝇血液发育标记基因mrj的多克隆抗体制备及检测
2012-11-22 06:04雷旻音李永青吴秀山
湖南师范大学自然科学学报 2012年4期
雷旻音,杨 哲,黄 婷,刘 丹,李永青,吴秀山
(湖南师范大学蛋白质化学及鱼类发育生物学教育部重点实验室,心脏发育研究中心,中国 长沙 410081)
mrj位于果蝇2号染色体右臂,全长18 474 bp,在其2-61个氨基酸处含有一个Dnaj的结构域.Dnaj包括两个结构域:N末端是与DnaK相互作用结构域,C末端是底物结合结构域.识别底物多肽的共有基序中间是4~5个疏水氨基酸残基的核心,两侧是酸性氨基酸残基[1].在蛋白质氨基酸顺序中每有36个氨基酸残基就有一个这样的基序,这种基序常常存在于β片层中.由该结构域表明该基因编码蛋白具有热休克及可稳定蛋白前体的非折叠构象的分子功能.有文献表明mrj和NFAT3c和srp在同一信号通路,NFAT3c突变可导致暂时性缺氧[2],而srp是果蝇血液的标志基因,因此作者推测mrj可能在果蝇血液的发育过程中起作用,因此mrj多克隆抗体的制备可用于果蝇血液发育信号通路的分析.另外有文献表明在果蝇体内mrj基因发生突变后,会影响果蝇细胞间的粘连[3].因此mrj基因的原核表达分析及其多克隆抗体的制备具有一定的研究意义和应用价值[4-5].
1 材料与方法
1.1 实验试剂及材料
大肠杆菌Rosetta菌种,PET-28a载体菌种以及E.coliDH5α菌种为本实验室保种;pMD18-T载体和连接酶购自大连宝生物公司;10×Loading buffer, Taq DNA聚合酶,限制性内切酶NcoⅠ和SacⅠ购自深圳晶美公司;RNase购自Sangon公司;UNIQ-10柱式DNA 胶回收纯化试剂盒、蛋白胨、酵母提取物、甲叉双丙烯酰胺、IPTG(异丙基-в-D-硫代半乳糖苷)丙烯酰胺、氯化钠、过硫酸铵、等购自上海生工公司;质粒提取试剂盒(离心柱型)购自OMEGA;柱式DNA胶回收试剂盒购自TIANGEN;Glutathione SepharoseTM4B,购自Amersham Biotech公司;弗氏完全佐剂、不完全佐剂购自Sigma公司.
1.2 引物设计与合成
利用 Peptide antigen finder软件筛选出mrjDNA序列的亲水区域,然后用Primer Premier 5.0软件设计:mrj正义引物(sense):5′CGCGTCGACCATGGTTGACTAC3′(划线部分为NcoⅠ酶切位点);反义引物(Anti-sense)5′CGCCTCGAGCTATTGAAGGGAG3′(划线部分为SacⅠ酶切位点),引物由上海生工公司合成.
1.3 基因扩增及克隆
收集野生型果蝇约30只,置于液氮中30 min,接着用Trizol法提取总RNA,通过反转录得到果蝇cDNA,并以之为模板进行PCR扩增(反应条件为:95 ℃变性5 min;95 ℃ 30 s,58 ℃ 30 s,72 ℃ 1 min 30 s,28个循环;72 ℃延伸8 min).将含有780 bp目的片段的PCR产物纯化回收后,连接至pMD18-T载体,转入大肠杆菌DH5α感受态细胞中,通过NcoⅠ和SacⅠ双酶切检测后,筛选出阳性单克隆,选择一个阳性克隆送至上海生工公司进行测序分析,将测序正确的pMD18-T-mrj重组质粒上的目的DNA片段用限制性内切酶NcoⅠ和SacⅠ切下,纯化回收后连入pET-28a载体,转入Rosetta感受态细胞中,提取质粒经NcoⅠ和SacⅠ双酶切验证后得到重组表达质粒pET-28a-mrj.
1.4 mrj融合蛋白的诱导表达
将构建好的重组质粒pET-28a-mrj用热激法转入大肠杆菌菌株Rosetta,LB培养基(含100 mg/L 氯霉素,卡那霉素)37 ℃培养,转接扩大培养至OD600约为0.6,按0.1 mmol/L 加入IPTG, 25 ℃诱导,2、4、5、6 h各取1 mL菌液,确定最佳诱导时间[6].
1.5 mrj融合蛋白亲和纯化
通过各时段取样比较,选择在IPTG浓度为0.1 mmol/L,诱导时长5 h的条件下进行大量诱导.离心收集菌体,PBS 重悬细胞后超声裂解细胞至菌液清亮,接着12 000 r/min离心 10 min,收集上清. 4 ℃下与经Binding Buffer漂洗活化后的Ni-IDA凝胶柱结合,用Washing Buffer洗去杂蛋白,再用Elution Buffer洗脱目的蛋白,获得纯化的His-mrj融合蛋白,-80 ℃保存备用[7].
1.6 mrj多克隆抗体的制备
将纯化得到的His-mrj蛋白质冻干,取1 mg 抗原蛋白溶于0.5 mL 生理盐水中,按体积比1∶1 与弗氏完全佐剂(Sigma 公司)在注射器中推成乳剂,对2 月龄雄性新西兰大白兔进行背部皮下免疫注射,分散10~20 个点;在第14天、第21天、第28天再将溶于0.5 mL 生理盐水中的0.5 mg 抗原蛋白与弗氏不完全佐剂(Sigma 公司)按体积比1∶1 在注射器中推成乳剂,进行背部皮下多点注射.第35天颈主动脉取血.兔血放在4 ℃冰箱静置过夜后,以3 000 r/min离心10 min,取血清分装保存于-80 ℃.
1.7 mrj多克隆抗体的检测
收获免疫兔血清,用果蝇心脏组织蛋白进行mrj抗体的效价测定,设置抗体稀释浓度分别为1∶500,1∶1 000,1∶2 000,1∶3 000.同等条件下,用免疫前兔血清进行Western blot分析作为对照.
2 结果
2.1 mrj基因克隆至表达载体
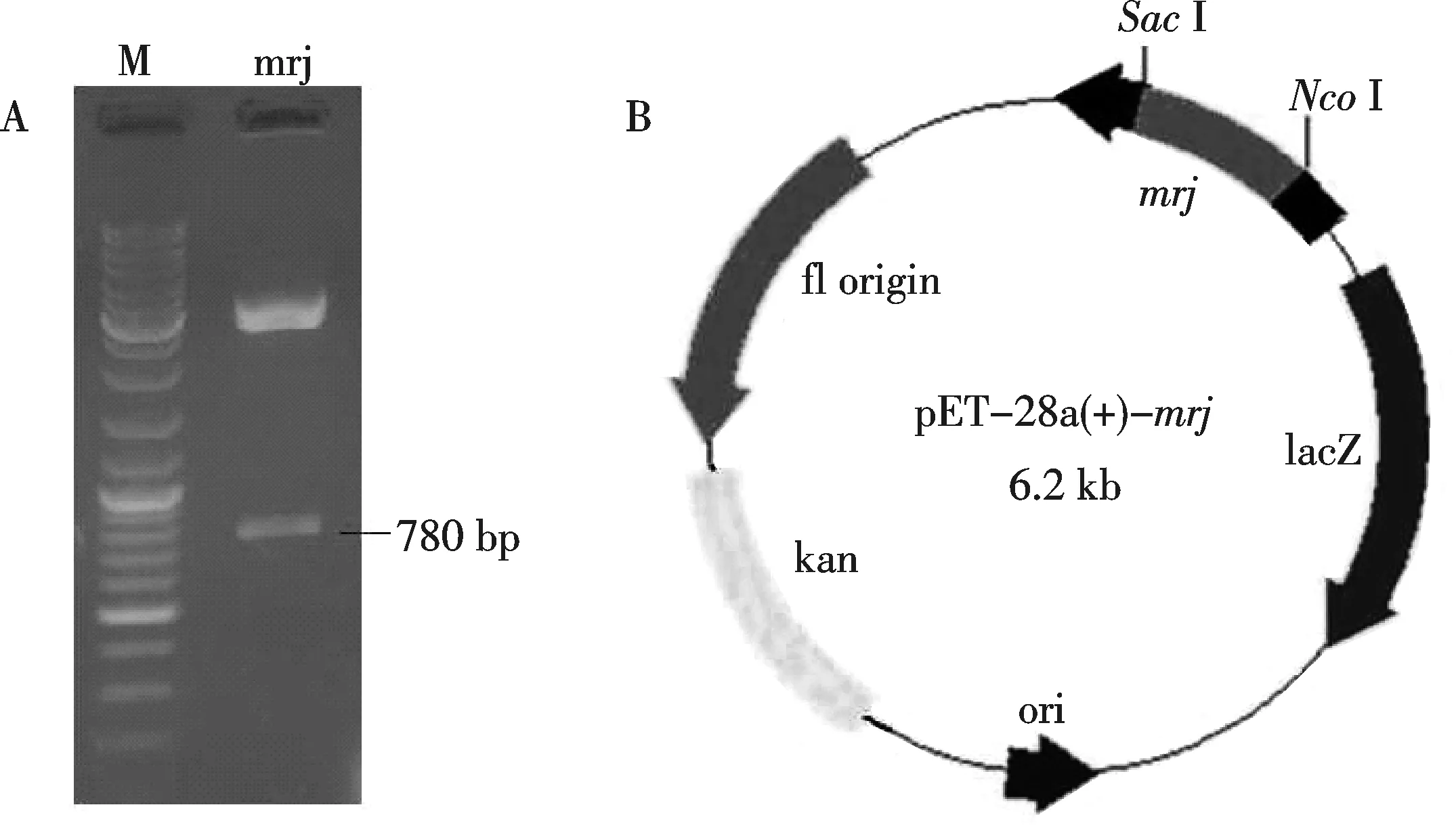
A: NcoⅠ/Sac Ⅰ双酶切结果;B: pET-28a-mrj质粒示意图图1 双酶切鉴定及pET-28a-mrj质粒示意图
将mrj基因部分编码序列连入pMD-18-T载体中,再用限制性内切酶NcoⅠ和SacⅠ双酶切pMD18-T-mrj,将得到的mrj基因目的片段连入pET-28a,并转入大肠杆菌DH5α,经NcoⅠ和SacⅠ双酶切鉴定(如图1:A)及测序分析,结果显示已成功构建了pET-28a-mrj(如图1:B).
2.1 mrj融合蛋白诱导表达及纯化
将重组质粒pET-28a-mrj转入Rosetta菌株,以不同浓度IPTG(0、0.1、0.2、0.4 mmol/L)25 ℃下进行诱导,取IPTG浓度为0.1 mmol/L诱导2,4,5,6 h,IPTG浓度为0.2 mmol/L诱导2,4,6 h的菌液各1 mL进行检测,可见一条相对分子质量约为33 000大小的特异条带,与预期的mrj融合蛋白相对分子质量一致(如图2)[8-9].
2.2 mrj融合蛋白的表达及纯化
通过对比,最终选择在诱导温度25 ℃,IPTG浓度为0.1 mmol/L,诱导时长5 h的条件下大量诱导得到可溶蛋白.并将诱导所得融合蛋白经Ni-IDA凝胶柱亲和纯化,通过免疫印记及考马斯亮蓝染色后显示目的条带纯度较高,可进行蛋白免疫(如图3).

M:Marker0671;1:pET-28a-mrj未诱导蛋白表达;2:0.1 mmol/L IPTG 诱导pET-28a-mrj 2 h时蛋白表达;3:0.1 mmol/L IPTG 诱导pET-28a-mrj 4 h时蛋白表达;4:0.1 mmol/L IPTG 诱导pET-28a-mrj 5 h时蛋白表达;5:0.1 mmol/L IPTG 诱导pET-28a-mrj 6 h时蛋白表达;6:0.2 mmol/L IPTG 诱导pET-28a-mrj 2 h时蛋白表达;7:0.2 mmol/L IPTG 诱导pET-28a-mrj 4 h时蛋白表达;8:0.2 mmol/L IPTG 诱导pET-28a-mrj 6 h时蛋白表达.图2 pET-28a-mrj诱导表达图

图3 融合蛋白His-mrj的纯化图
2.3 mrj多克隆抗体Western Blot鉴定
用pET-28a-mrj转化Rosetta菌株诱导表达的总蛋白对所制备的mrj多克隆抗体的特异性进行Western blot检测,将制备的多克隆抗体按浓度梯度稀释(1∶500、1∶1 000、1∶2 000、1∶3 000)作为一抗进行检测,并用空白血清作为阴性对照,可见His-mrj蛋白所处位置有一条特异性的杂交带出现(如图4),检测结果表明该抗体的特异性和敏感性均可达到今后实验需要[10].
2.4 mrj多克隆抗体有效性的鉴定
用RNAi干扰品系,利用western blot鉴定所制备mrj多克隆抗体的有效性.将制备的mrj多克隆抗体按照1∶2 000的浓度比稀释作为一抗,用野生型果蝇胚胎作阴性对照[11](如图5).

图4 mrj多克隆抗体Western-blotting鉴定

图5 mrj多克隆抗体有效性的鉴定
3 讨论
真核细胞的密码子和原核系统的不同,因此,在用原核系统表达真核基因的时候,真核基因中的一些密码子对于原核细胞来说可能是稀有密码子,从而导致表达效率和表达水平很低.本实验所用宿主菌株为Rosetta菌株,因为相比较BL21菌株,Rosetta 是携带pRARE2质粒的BL21衍生菌,能补充大肠杆菌缺乏的7种稀有密码子(AUA, AGG, AGA, CUA, CCC, GGA 及CGG)对应的 tRNA,可提高外源基因、尤其是真核基因在原核系统中的表达水平.另外,Rosetta还能够促进二硫键的形成,帮助表达需要借助二硫键形成正确折叠构象的真核蛋白.
本实验所用免疫动物为2月龄雄性新西兰大白兔,因为雌性动物特别是妊娠动物用于制备免疫有时不产生抗体.由于对免疫应答存在个别差异,免疫时应同时选用数只动物进行免疫. 抗原的免疫剂量依照给予动物的种类、免疫周期以及所要求的抗体特性等不同而不同.剂量过低,不能引起足够强的免疫刺激;免疫剂量过多,有可能引起免疫耐受.在一定的范围内,抗体的效价随注射剂量的增加而增高.蛋白质抗原的免疫剂量比多糖类抗原高. 免疫剂量与注射途径有关.通常,静脉注射剂量大于皮下注射,而皮下注射又比掌内和跖内皮下注射剂量大,也可采用淋巴结内注射法.加佐剂比不加佐剂的注射剂量小.如要制备高度特异性的抗血清,可选用低剂量抗原短程免疫法.如需要获得高效价的抗血清,宜采用大剂量长程免疫法.免疫周期长者,可少量多次;免疫周期短者,应大量少次. 两次注射的间隔时间应长短适宜,太短起不到再次反应的效果,太长则失去了前一次激发的敏感作用.一般间隔时间应为5~7 d,加佐剂者应为2周左右.
虽然用纯化蛋白检测所制备多克隆抗体的效价时在一定程度上验证了干抗体的有效性,但western-blot实验时所用蛋白一般为果蝇胚胎提取的全蛋白,此时,若所制备的抗体特异性不强,实验中非目的条带过多会严重影响实验的准确性.
mrj是果蝇发育中一个关键的标志基因[12],我们所熟知的是它可与尿激酶受体共同作用增强细胞间的粘连[13].但有文献表明它很可能与果蝇的血液发育相关,因此mrj多克隆抗体在今后的实验中还可用做体内CO-ip实验,寻找在体内与其有相互作用的蛋白,或干扰其所在信号通路的其他蛋白,将该多克隆抗体用作胚胎抗体染色,或western-blot等实验.
参考文献:
[1] MITRA A, MENEZES M E, SHEVDE L A,etal. DNAJB6 induces degradation of beta-catenin and causes partial reversal of mesenchymal phenotype[J].J Biol Chem, 2010,285(32):24686-24694.
[2] WATSON E D, HUGHES M, SIMMONS D G,etal. Cell-cell adhesion defects inMrjmutant trophoblast cells are associated with failure to pattern the chorion during early placental development[J]. Dev Dyn, 2011,240(11):2505-2519.
[3] NANDURI J, YUAN G, KUMAR G K,etal. Transcriptional responses to intermittent hypoxia[J].Respir Physiol Neurobiol, 2008,164(1-2):277-281.
[4] 吴秀山,王跃群,袁婺洲,等.心脏发育概论[M].北京:科学出版社, 2006:141-143.
[5] 吴秀山,袁婺洲,王跃群,等.心脏发育研究[M].长沙:湖南科学技术出版社, 2004:214-302.
[6] 满 贤,吴秀山,李永青,等.斑马鱼Lefty1特异性多克隆抗体的制备[J].激光生物学报, 2010,19(5):658-689.
[7] 刘 红,任笑蒙,李 君,等.果蝇组织总RNA的电泳谱型鉴定[J].医学研究通讯, 2004,33(4):48-49.
[8] 唐威华,张景六,王宗阳,等.SDS-PAGE法测定His-tag融合蛋白分子量产生偏差的原因[J].植物生理学报, 2010,33(1):64-68.
[9] 张守涛,郭蔼光.Fibrolase表达菌自诱导摇瓶发酵研究[J].安徽农业科学, 2008,36(9):3715-3716.
[10] 王雁云,罗开梅,李永青,等.一种快速的胚胎组织总RNA的提取方法[J].生命科学研究, 2001,5(1):88-90.
[11] 龚 琳,王跃群,袁婺洲,等.果蝇nulp1特异性多克隆抗体制备及检测[J].湖南师范大学自然科学学报, 2011,34(4):69-73.
[12] 万永奇,谢 维.科学与人类疾病研究的重要模型—果蝇[J].生命科学, 2006,18(5):425-429.
[13] BOCK C E, LIN Z, MEKKAWY A H,etal. Interaction between urokinase receptor and heat shock protein MRJ enhances cell adhesion[J]. Int J Oncol, 2010,36(5):1155-1163.
猜你喜欢
学苑创造·A版(2022年3期)2022-03-29
昆明医科大学学报(2021年3期)2021-07-22
烟台果树(2021年2期)2021-07-21
学苑创造·A版(2019年6期)2019-07-11
测控技术(2018年7期)2018-12-09
中成药(2018年10期)2018-10-26
山西医科大学学报(2017年11期)2017-12-01
湖南中医药大学学报(2016年1期)2016-12-01
天津科技大学学报(2016年3期)2016-08-02
食品研究与开发(2016年9期)2016-06-13
